
Biobanked Glioblastoma Patient-Derived Organoids as a Precision Medicine Model to Study Inhibition of Invasion

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Cytotoxicity of the Compounds on U-251 MG Spheroids
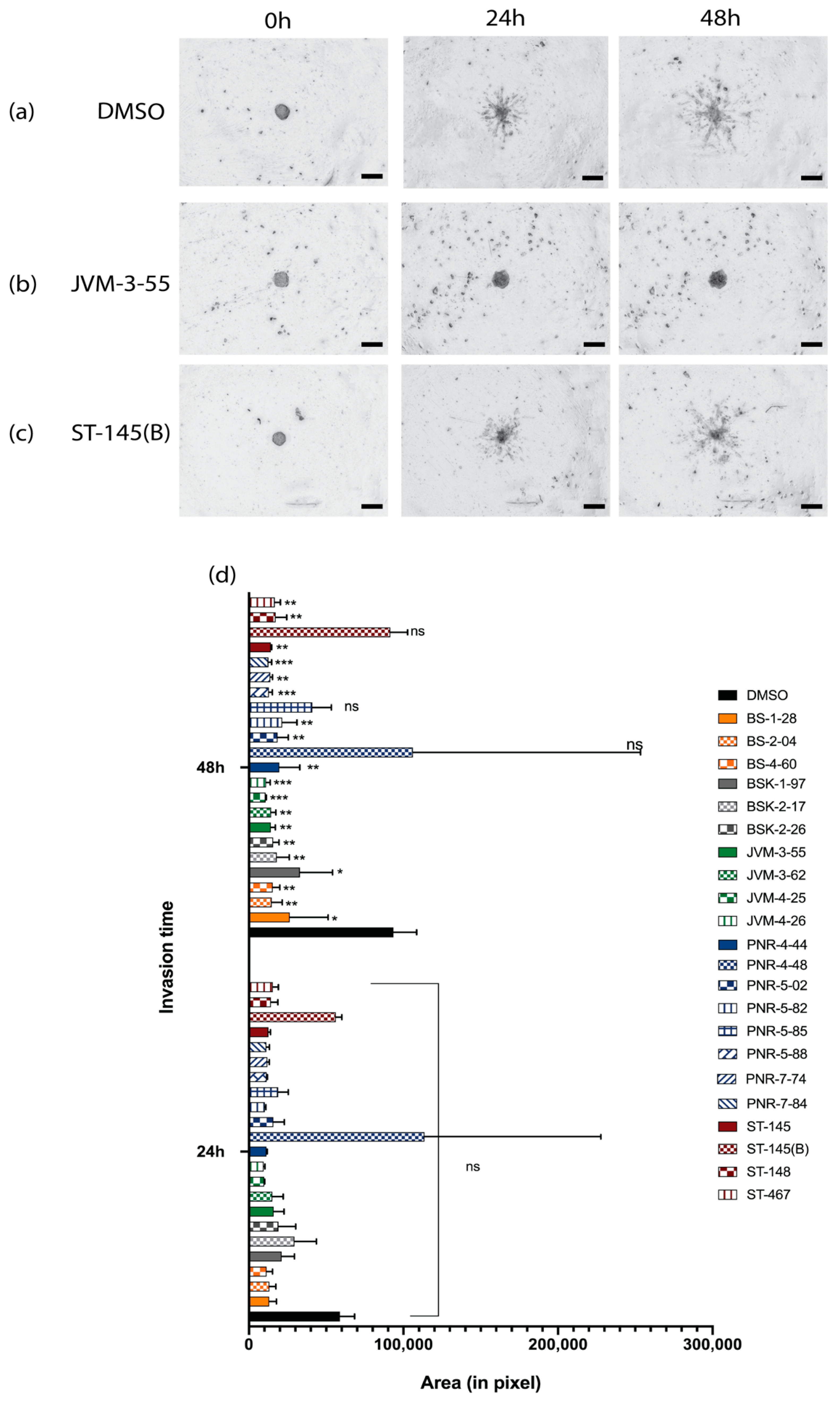
2.2. Invasion Inhibition of the Compounds on U-251 MG Cell Spheroids and the Development and Implementation of Automatic Invasion Assay Analysis
2.3. Development of GBOs
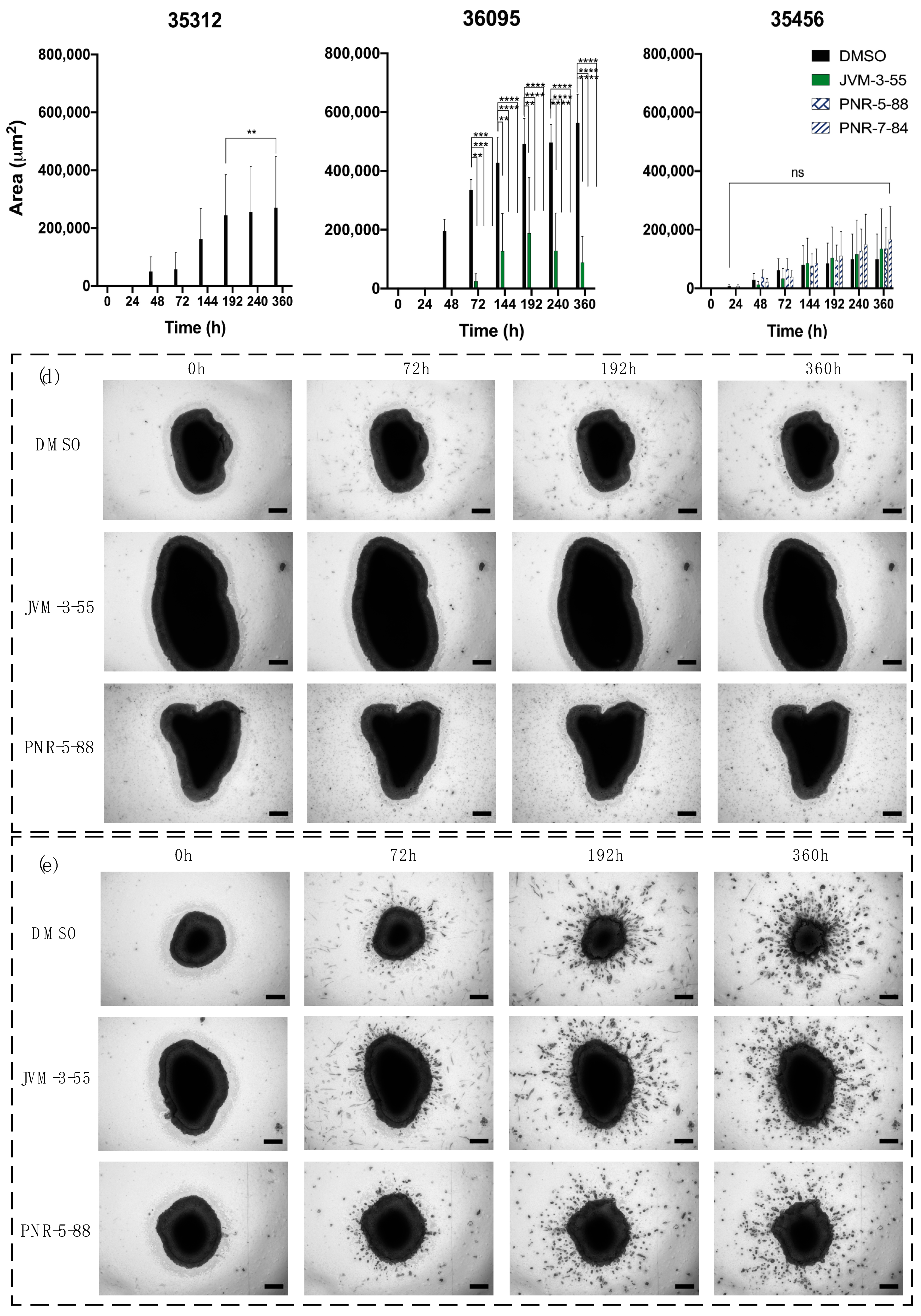
2.4. Determination of Invasion Inhibition on GBOs by Selected Compounds
3. Discussion and Conclusions
4. Materials and Methods
4.1. Drug Panel Selected for Experimentation
4.2. Spheroid Culture, Standard Curve Generation, and Cellular Viability in 3D GBM Spheroids
4.3. Spheroid Culture for Invasion Assay
4.4. Spheroid Measurements for Automatic Invasion Assay Analysis
4.5. Collection, Dissection, and Processing of Patient Glioblastoma
4.6. Generation of GBOs from Resected Patient Glioblastoma Tissue
4.7. GBO Culture for Invasion Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vollmann-Zwerenz, A.; Leidgens, V.; Feliciello, G.; Klein, C.A.; Hau, P. Tumor cell invasion in glioblastoma. Int. J. Mol. Sci. 2020, 21, 1932. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.Y.; Oliva, C.R.; Noman, A.S.M.; Allen, B.G.; Goswami, P.C.; Zakharia, Y.; Monga, V.; Spitz, D.R.; Buatti, J.M.; Griguer, C.E. Radioresistance in glioblastoma and the development of radiosensitizers. Cancers 2020, 12, 2511. [Google Scholar] [CrossRef]
- Da Ros, M.; De Gregorio, V.; Iorio, A.L.; Giunti, L.; Guidi, M.; De Martino, M.; Genitori, L.; Sardi, I. Glioblastoma chemoresistance: The double play by microenvironment and blood-brain barrier. Int. J. Mol. Sci. 2018, 19, 2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekman, M.L.; Maas, S.L.; Abels, E.R.; Mempel, T.R.; Krichevsky, A.M.; Breakefield, X.O. Multidimensional communication in the microenvirons of glioblastoma. Nat. Rev. Neurol. 2018, 14, 482–495. [Google Scholar] [CrossRef]
- Zhang, X.; Ding, K.; Wang, J.; Li, X.; Zhao, P. Chemoresistance caused by the microenvironment of glioblastoma and the corresponding solutions. Biomed. Pharmacother. 2019, 109, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ou, A.; Yung, W.; Majd, N. Molecular mechanisms of treatment resistance in glioblastoma. Int. J. Mol. Sci. 2021, 22, 351. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, B.; Zhou, C.; Li, Y.; Li, B.; Yu, M.; Luo, Y.; Gao, L.; Zhang, D.; Xue, Q.; et al. The comparison genomics analysis with glioblastoma multiforme (GBM) cells under 3D and 2D cell culture conditions. Colloids Surf. B Biointerfaces 2018, 172, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Hicks, W.H.; Bird, C.E.; Traylor, J.I.; Shi, D.D.; El Ahmadieh, T.Y.; Richardson, T.E.; McBrayer, S.K.; Abdullah, K.G. Contemporary mouse models in glioma research. Cells 2021, 10, 712. [Google Scholar] [CrossRef]
- Paolillo, M.; Comincini, S.; Schinelli, S. In vitro glioblastoma models: A journey into the third dimension. Cancers 2021, 13, 2449. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Mukohara, T.; Shimono, Y.; Funakoshi, Y.; Chayahara, N.; Toyoda, M.; Kiyota, N.; Takao, S.; Kono, S.; Nakatsura, T.; et al. Comparison of 2D-and 3D-culture models as drug-testing platforms in breast cancer. Oncol. Rep. 2015, 33, 1837–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darrigues, E.; Nima, Z.A.; Nedosekin, D.A.; Watanabe, F.; Alghazali, K.M.; Zharov, V.P.; Biris, A.S. Tracking gold nanorods’ interaction with large 3D pancreatic-stromal tumor spheroids by multimodal imaging: Fluorescence, photoacoustic, and photothermal microscopies. Sci. Rep. 2020, 10, 3362. [Google Scholar] [CrossRef]
- Darrigues, E.; Nima, Z.A.; Griffin, R.J.; Anderson, J.M.; Biris, A.S.; Rodriguez, A. 3D cultures for modeling nanomaterial-based photothermal therapy. Nanoscale Horiz. 2020, 5, 400–430. [Google Scholar] [CrossRef]
- Darrigues, E.; Al Sudani, Z.A.N.; Watanabe, F.; Biris, A.S. Plasmonic gap-enhanced Raman tag nanorods for imaging 3D pancreatic spheroids using surface-enhanced Raman spectroscopy and darkfield microscopy. Nanotechnology 2020, 32, 095104. [Google Scholar] [CrossRef]
- Torsvik, A.; Stieber, D.; Enger, P.Ø.; Golebiewska, A.; Molven, A.; Svendsen, A.; Westermark, B.; Niclou, S.P.; Olsen, T.K.; Enger, M.C.; et al. U-251 revisited: Genetic drift and phenotypic consequences of long-term cultures of glioblastoma cells. Cancer Med. 2014, 3, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Holbeck, S.L.; Camalier, R.; Crowell, J.A.; Govindharajulu, J.P.; Hollingshead, M.; Anderson, L.W.; Polley, E.; Rubinstein, L.; Srivastava, A.; Wilsker, D.; et al. The National Cancer Institute ALMANAC: A comprehensive screening resource for the detection of anticancer drug pairs with enhanced therapeutic activity. Cancer Res. 2017, 77, 3564–3576. [Google Scholar] [CrossRef] [Green Version]
- Watkins, S.; Robel, S.; Kimbrough, I.F.; Robert, S.M.; Ellis-Davies, G.; Sontheimer, H. Disruption of astrocyte-vascular coupling and the blood–brain barrier by invading glioma cells. Nat. Commun. 2014, 5, 4196. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Chen, Z.-L.; Norris, E.H.; Strickland, S. Astrocytic laminin regulates pericyte differentiation and maintains blood brain barrier integrity. Nat. Commun. 2014, 5, 3413. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, F.; Xiong, N.; Xu, H.; Chai, S.; Wang, H.; Wang, J.; Zhao, H.; Jiang, X.; Fu, P.; et al. Remodelling and treatment of the blood-brain barrier in glioma. Cancer Manag. Res. 2021, 13, 4217–4232. [Google Scholar] [CrossRef] [PubMed]
- Hubert, C.G.; Rivera, M.; Spangler, L.C.; Wu, Q.; Mack, S.C.; Prager, B.C.; Couce, M.; McLendon, R.E.; Sloan, A.E.; Rich, J.N. A three-dimensional organoid culture system derived from human glioblastomas recapitulates the hypoxic gradients and cancer stem cell heterogeneity of tumors found in vivo. Cancer Res. 2016, 76, 2465–2477. [Google Scholar] [CrossRef] [Green Version]
- Jacob, F.; Ming, G.-L.; Song, H. Generation and biobanking of patient-derived glioblastoma organoids and their application in CAR T cell testing. Nat. Protoc. 2020, 15, 4000–4033. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Salinas, R.D.; Zhang, D.Y.; Nguyen, P.T.T.; Schnoll, J.G.; Wong, S.Z.H.; Thokala, R.; Sheikh, S.; Saxena, D.; Prokop, S.; et al. A patient-derived glioblastoma organoid model and biobank recapitulates inter- and intra-tumoral heterogeneity. Cell 2020, 180, 188–204.e22. [Google Scholar] [CrossRef] [PubMed]
- Lenin, S.; Ponthier, E.; Scheer, K.G.; Yeo, E.C.; Tea, M.N.; Ebert, L.M.; Oksdath Mansilla, M.; Poonnoose, S.; Baumgartner, U.; Day, B.W.; et al. A drug screening pipeline using 2D and 3D patient-derived in vitro models for pre-clinical analysis of therapy response in glioblastoma. Int. J. Mol. Sci. 2021, 22, 4322. [Google Scholar] [CrossRef]
- Darrigues, E.; Elberson, B.W.; De Loose, A.; Lee, M.P.; Green, E.; Benton, A.M.; Sink, L.G.; Scott, H.; Gokden, M.; Day, J.D.; et al. Brain tumor biobank development for precision medicine: Role of the neurosurgeon. Front. Oncol. 2021, 11, 1260. [Google Scholar] [CrossRef]
- Shoemaker, R.H. The NCI60 human tumour cell line anticancer drug screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Del Duca, D.; Werbowetski, T.; Del Maestro, R.F. Spheroid preparation from hanging drops: Characterization of a model of brain tumor invasion. J. Neuro.-Oncol. 2004, 67, 295–303. [Google Scholar] [CrossRef]
- Bommagani, S.; Ponder, J.; Penthala, N.R.; Janganati, V.; Jordan, C.T.; Borrelli, M.J.; Crooks, P.A. Indole carboxylic acid esters of melampomagnolide B are potent anticancer agents against both hematological and solid tumor cells. Eur. J. Med. Chem. 2017, 136, 393–405. [Google Scholar] [CrossRef]
- Janganati, V.; Ponder, J.; Jordan, C.T.; Borrelli, M.J.; Penthala, N.R.; Crooks, P.A. Dimers of melampomagnolide B exhibit potent anticancer activity against hematological and solid tumor cells. J. Med. Chem. 2015, 58, 8896–8906. [Google Scholar] [CrossRef] [Green Version]
- Janganati, V.; Ponder, J.; Balasubramaniam, M.; Bhat-Nakshatri, P.; Bar, E.E.; Nakshatri, H.; Jordan, C.T.; Crooks, P.A. MMB triazole analogs are potent NF-κB inhibitors and anti-cancer agents against both hematological and solid tumor cells. Eur. J. Med. Chem. 2018, 157, 562–581. [Google Scholar] [CrossRef] [Green Version]
- Penthala, N.R.; Ketkar, A.; Sekhar, K.R.; Freeman, M.L.; Eoff, R.L.; Balusu, R.; Crooks, P.A. 1-Benzyl-2-methyl-3-indolylmethylene barbituric acid derivatives: Anti-cancer agents that target nucleophosmin 1 (NPM1). Bioorganic Med. Chem. 2015, 23, 7226–7233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penthala, N.R.; Madhukuri, L.; Thakkar, S.; Madadi, N.R.; Lamture, G.; Eoff, R.L.; Crooks, P.A. Synthesis and anti-cancer screening of novel heterocyclic-(2H)-1,2,3-triazoles as potential anti-cancer agents. MedChemComm 2015, 6, 1535–1543. [Google Scholar] [CrossRef] [Green Version]
- Penthala, N.R.; Zong, H.; Ketkar, A.; Madadi, N.R.; Janganati, V.; Eoff, R.L.; Guzman, M.L.; Crooks, P.A. Synthesis, anticancer activity and molecular docking studies on a series of heterocyclic trans-cyanocombretastatin analogues as antitubulin agents. Eur. J. Med. Chem. 2015, 92, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Madadi, N.R.; Zong, H.; Ketkar, A.; Zheng, C.; Penthala, N.R.; Janganati, V.; Bommagani, S.; Eoff, R.L.; Guzman, M.L.; Crooks, P.A. Synthesis and evaluation of a series of resveratrol analogues as potent anti-cancer agents that target tubulin. MedChemComm 2015, 6, 788–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madadi, N.R.; Penthala, N.R.; Howk, K.; Ketkar, A.; Eoff, R.L.; Borrelli, M.J.; Crooks, P.A. Synthesis and biological evaluation of novel 4,5-disubstituted 2H-1,2,3-triazoles as cis-constrained analogues of combretastatin A-4. Eur. J. Med. Chem. 2015, 103, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H. Role of the NFκB-signaling pathway in cancer. OncoTargets Ther. 2018, 11, 2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriconi, C.; Palmieri, V.; Di Santo, R.; Tornillo, G.; Papi, M.; Pilkington, G.; De Spirito, M.; Gumbleton, M. INSIDIA: A FIJI Macro delivering high-throughput and high-content spheroid invasion analysis. Biotechnol. J. 2017, 12, 1700140. [Google Scholar] [CrossRef] [PubMed]
- Forster, B.; Van De Ville, D.; Berent, J.; Sage, D.; Unser, M. Complex wavelets for extended depth-of-field: A new method for the fusion of multichannel microscopy images. Microsc. Res. Tech. 2004, 65, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akay, M.; Hite, J.; Avci, N.G.; Fan, Y.; Akay, Y.; Lu, G.; Zhu, J.-J. Drug screening of human GBM spheroids in brain cancer chip. Sci. Rep. 2018, 8, 15423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivakumar, H.; Devarasetty, M.; Kram, D.E.; Strowd, R.E.; Skardal, A. Multi-cell type glioblastoma tumor spheroids for evaluating sub-population-specific drug response. Front. Bioeng. Biotechnol. 2020, 8, 538663. [Google Scholar] [CrossRef]
- Fedrigo, C.A.; Grivicich, I.; Schunemann, D.P.; Chemale, I.M.; dos Santos, D.; Jacovas, T.; Boschetti, P.S.; Jotz, G.P.; Braga Filho, A.; da Rocha, A.B. Radioresistance of human glioma spheroids and expression of HSP70, p53 and EGFr. Radiat. Oncol. 2011, 6, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinci, M.; Box, C.; Eccles, S.A. Three-dimensional (3D) tumor spheroid invasion assay. J. Vis. Exp. 2015, 99, e52686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma, C.R.; Zimmermann, M.; Agarkova, I.; Kelm, J.M.; Krek, W. 3D cell culture systems modeling tumor growth determinants in cancer target discovery. Adv. Drug Deliv. Rev. 2014, 69–70, 29–41. [Google Scholar] [CrossRef]
- Rebelo, S.P.; Pinto, C.; Martins, T.R.; Harrer, N.; Estrada, M.F.; Loza-Alvarez, P.; Cabeçadas, J.; Alves, P.M.; Gualda, E.J.; Sommergruber, W.; et al. 3D-3-culture: A tool to unveil macrophage plasticity in the tumour microenvironment. Biomaterials 2018, 163, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Calar, K.; De La Puente, P. Mimicking tumor hypoxia and tumor-immune interactions employing three-dimensional in vitro models. J. Exp. Clin. Cancer Res. 2020, 39, 75. [Google Scholar] [CrossRef] [PubMed]
- Linkous, A.; Balamatsias, D.; Snuderl, M.; Edwards, L.; Miyaguchi, K.; Milner, T.; Reich, B.; Cohen-Gould, L.; Storaska, A.; Nakayama, Y.; et al. Modeling patient-derived glioblastoma with cerebral organoids. Cell Rep. 2019, 26, 3203–3211.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goranci-Buzhala, G.; Mariappan, A.; Gabriel, E.; Ramani, A.; Ricci-Vitiani, L.; Buccarelli, M.; D’Alessandris, Q.G.; Pallini, R.; Gopalakrishnan, J. Rapid and efficient invasion assay of glioblastoma in human brain organoids. Cell Rep. 2020, 31, 107738. [Google Scholar] [CrossRef]
- Anderson, H.J.; Galileo, D.S. Small-molecule inhibitors of FGFR, integrins and FAK selectively decrease L1CAM-stimulated glioblastoma cell motility and proliferation. Cell. Oncol. 2016, 39, 229–242. [Google Scholar] [CrossRef]
- Eilenberger, C.; Rothbauer, M.; Ehmoser, E.-K.; Ertl, P.; Küpcü, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Sci. Rep. 2019, 9, 4863. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Lin, C.; Cheng, J.; Su, J.; Zhao, H.; Liu, T.; Wen, X.; Zhao, P. Generation of multicellular tumor spheroids with microwell-based agarose scaffolds for drug testing. PLoS ONE 2015, 10, e0130348. [Google Scholar] [CrossRef] [Green Version]
- Achilli, T.-M.; McCalla, S.; Meyer, J.; Tripathi, A.; Morgan, J.R. Multilayer spheroids to quantify drug uptake and diffusion in 3D. Mol. Pharm. 2014, 11, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Shuford, S.; Lipinski, L.; Abad, A.; Smith, A.M.; Rayner, M.; O’Donnell, L.; Stuart, J.; Mechtler, L.L.; Fabiano, A.J.; Edenfield, J.; et al. Prospective prediction of clinical drug response in high-grade gliomas using an ex vivo 3D cell culture assay. Neuro-Oncol. Adv. 2021, 3, vdab065. [Google Scholar] [CrossRef] [PubMed]
- Arganda-Carreras, I.; Kaynig, V.; Rueden, C.; Schindelin, J.; Cardona, A.; Seung, H.S. Trainable_Segmentation: Release V3.1.2. Available online: https://zenodo.org/record/59290#.YVlJjLgzZPZ (accessed on 3 August 2016).
- Arganda-Carreras, I.; Kaynig, V.; Rueden, C.; Eliceiri, K.; Schindelin, J.E.; Cardona, A.; Seung, H.S. Trainable Weka Segmentation: A machine learning tool for microscopy pixel classification. Bioinformatics 2017, 33, 2424–2426. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Simplified Group Identification | Family/Chemical Group | Activity | References |
|---|---|---|---|---|
| BS-1-28 | BS | MMB (Melampomagnolide b) indole esters | NFk-B inhibitors | [27] |
| BS-2-04 | BS | MMB indole esters | NFk-B inhibitors | [27] |
| BS-4-60 | BS | MMB indole esters | NFk-B inhibitors | [27] |
| BSK-1-97 | BSK | MMB-Thiadiazolidinone | NFk-B and GSK-3 B inhibitors | Non-published |
| BSK-2-17 | BSK | MMB-Thiadiazolidinone | NFk-B and GSK-3 B inhibitors | Non-published |
| BSK-2-26 | BSK | MMB-Thiadiazolidinone | NFk-B and GSK-3 B inhibitors | Non-published |
| JVM-3-55 | JVM | MMB dimers | NFk-B inhibitors | [28] |
| JVM-3-62 | JVM | MMB dimers | NFk-B inhibitors | [28] |
| JVM-4-25 | JVM | MMB Triazole dimers | NFk-B inhibitors | [29] |
| JVM-4-26 | JVM | MMB Triazole dimers | NFk-B inhibitors | [29] |
| PNR-4-44 | PNR | Heterocyclic trans-cyanocombretastatin analogues | Tubulin inhibitors | [32] |
| PNR-4-48 | PNR | Heterocyclic trans-cyanocombretastatin analogues | Tubulin inhibitors | [32] |
| PNR-5-02 | PNR | CA-4 triazole | Tubulin inhibitors | [31] |
| PNR-5-82 | PNR | Indole barbiturate and thiobarbiturates | COX-2 inhibitors | [30] |
| PNR-5-85 | PNR | Indole barbiturate and thiobarbiturates | COX-2 inhibitors | [30] |
| PNR-5-88 | PNR | Indole barbiturate and thiobarbiturates | COX-2 inhibitors | [30] |
| PNR-7-74 | PNR | Heterocyclic trans-cyanocombretastatin | Tubulin inhibitors | [32] |
| PNR-7-84 | PNR | Heterocyclic trans-cyanocombretastatin | Tubulin inhibitors | [32] |
| ST-145 | ST | Resveratrol analogues | Tubulin inhibitors | [33] |
| ST-145(B) | ST | Novel 4,5-disubstituted 2H-1,2,3-triazoles as cis-constrained analogues of combretastatin | Tubulin inhibitors | [34] |
| ST-148 | ST | Resveratrol analogues | Tubulin inhibitors | [33] |
| ST-467 | ST | Novel 4,5-disubstituted 2H-1,2,3-triazoles as cis-constrained analogues of combretastatin | Tubulin inhibitors | [34] |
| Compound | Activity/Inhibitors | Cytotoxicity | Cytotoxicity Effect in Comparision to Control | Invasion (48 h) | Invasion Inhibition |
|---|---|---|---|---|---|
| PNR-5-85 | COX-2 | * | - | *** | Low |
| BSK-1-97 | NFk-B and GSK-3 B | *** | - | * | High |
| JVM-4-26 | NFk-B | **** | - | ** | Moderate |
| BS-1-28 | NFk-B | **** | - | * | High |
| BS-4-60 | NFk-B | **** | - | ** | Moderate |
| JVM-3-62 | NFk-B | * | + | *** | Low |
| PNR-5-02 | Tubulin | * | + | ** | Moderate |
| PNR-7-74 | Tubulin | * | + | ** | Moderate |
| ST-467 | Tubulin | * | + | ** | Moderate |
| JVM-3-55 | NFk-B | *** | + | ** | Moderate |
| ST-148 | Tubulin | *** | + | ** | Moderate |
| ST-145 | Tubulin | **** | + | ** | Moderate |
| ST-145(B) | Tubulin | **** | + | ns | Very High |
| BS-2-04 | NFk-B | ns | = | ** | Moderate |
| BSK-2-17 | NFk-B and GSK-3 B | ns | = | ** | Moderate |
| BSK-2-26 | NFk-B and GSK-3 B | ns | = | ** | Moderate |
| PNR-4-48 | Tubulin | ns | = | ** | Moderate |
| PNR-5-88 | COX-2 | ns | = | ** | Moderate |
| JVM-4-25 | NFk-B | ns | = | *** | Low |
| PNR-7-84 | Tubulin | ns | = | *** | Low |
| PNR-4-44 | Tubulin | ns | = | ns | Very High |
| PNR-5-82 | COX-2 | ns | = | ns | Very High |
| Sample ID | Age | Sex | Diagnosis | MGMT Mutation | IDH Mutation |
|---|---|---|---|---|---|
| 35312 | 36 | Female | Gliosarcoma, rare glioblastoma, primary | no | no |
| 36095 | 57 | Male | Glioblastoma, recurrent | no | no |
| 35456 | 43 | Female | Glioblastoma, primary | yes | yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darrigues, E.; Zhao, E.H.; De Loose, A.; Lee, M.P.; Borrelli, M.J.; Eoff, R.L.; Galileo, D.S.; Penthala, N.R.; Crooks, P.A.; Rodriguez, A. Biobanked Glioblastoma Patient-Derived Organoids as a Precision Medicine Model to Study Inhibition of Invasion. Int. J. Mol. Sci. 2021, 22, 10720. https://doi.org/10.3390/ijms221910720
Darrigues E, Zhao EH, De Loose A, Lee MP, Borrelli MJ, Eoff RL, Galileo DS, Penthala NR, Crooks PA, Rodriguez A. Biobanked Glioblastoma Patient-Derived Organoids as a Precision Medicine Model to Study Inhibition of Invasion. International Journal of Molecular Sciences. 2021; 22(19):10720. https://doi.org/10.3390/ijms221910720
Chicago/Turabian StyleDarrigues, Emilie, Edward H. Zhao, Annick De Loose, Madison P. Lee, Michael J. Borrelli, Robert L. Eoff, Deni S. Galileo, Narsimha R. Penthala, Peter A. Crooks, and Analiz Rodriguez. 2021. "Biobanked Glioblastoma Patient-Derived Organoids as a Precision Medicine Model to Study Inhibition of Invasion" International Journal of Molecular Sciences 22, no. 19: 10720. https://doi.org/10.3390/ijms221910720