
IL-33 Enhances IFNγ and TNFα Production by Human MAIT Cells: A New Pro-Th1 Effect of IL-33


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
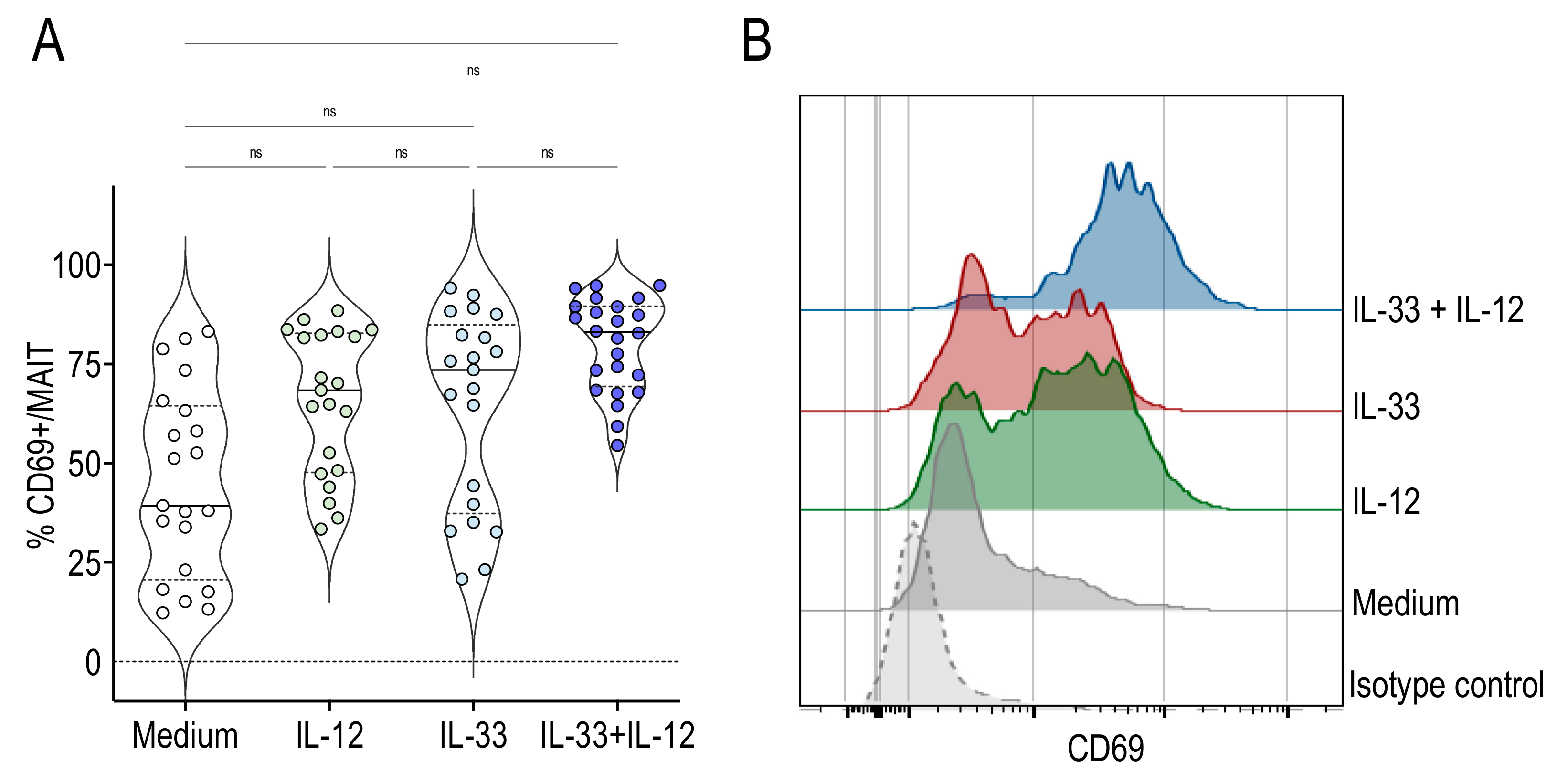
2.1. IL-33 Associated with IL-12 Activated Human MAIT Cells
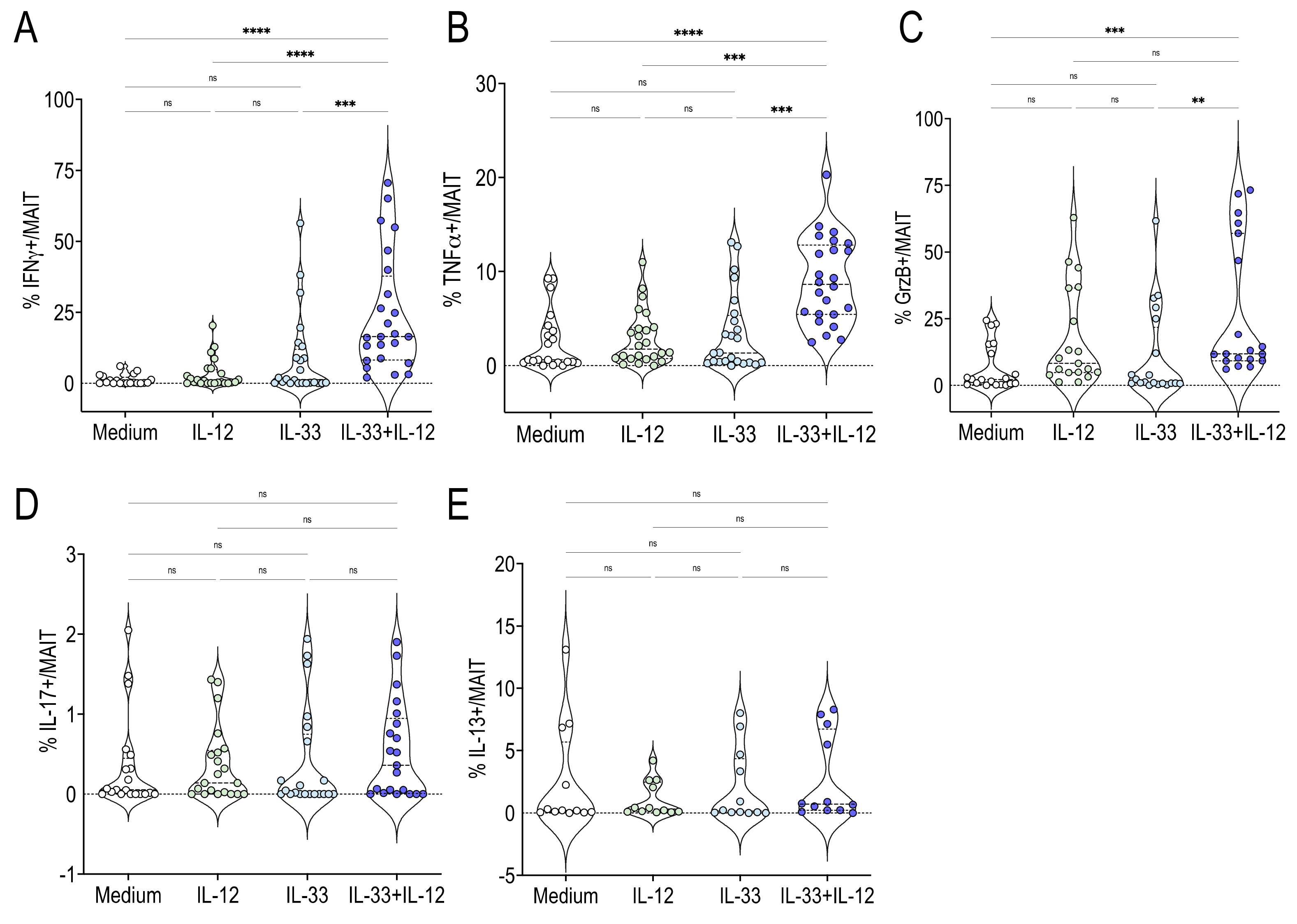
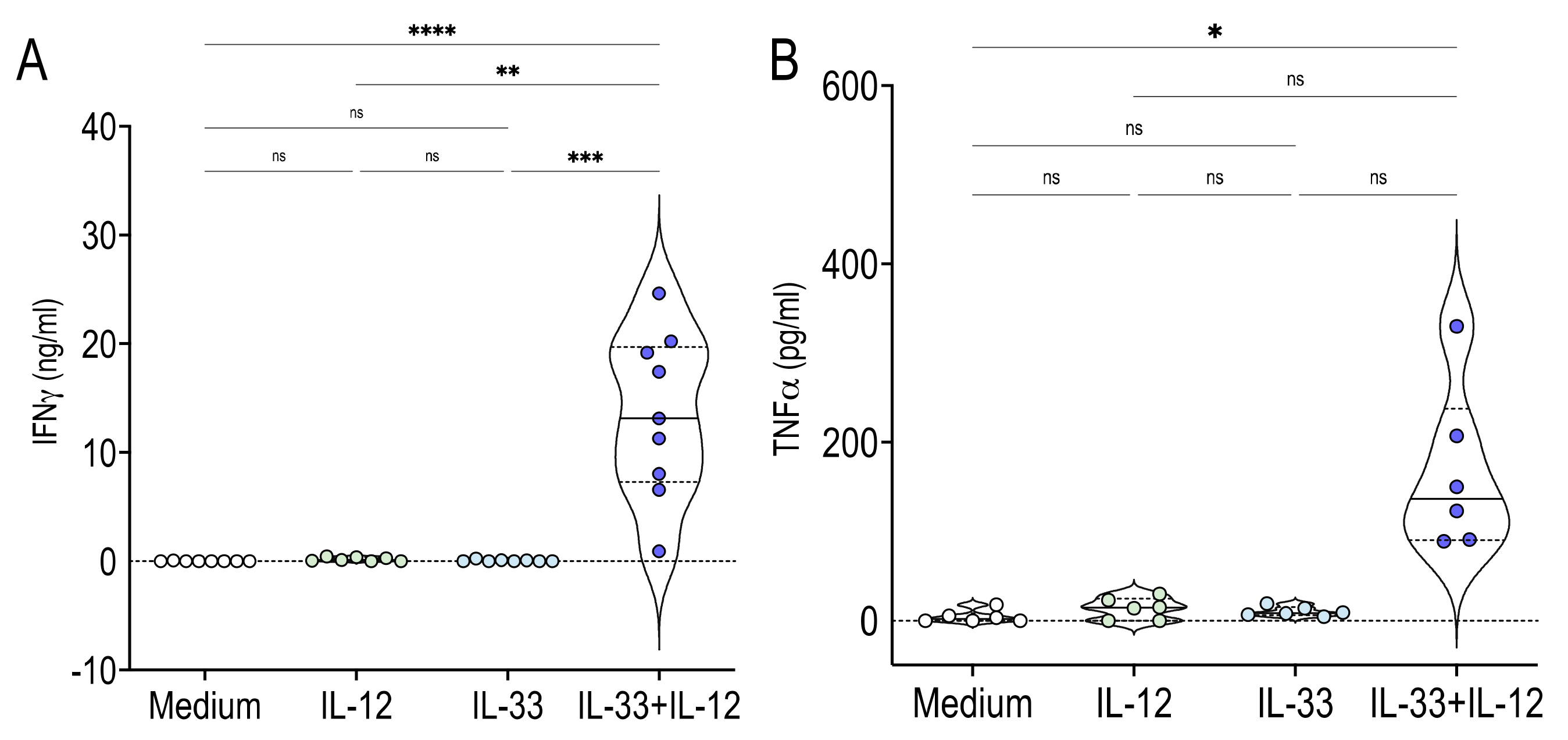
2.2. Human MAIT Cells Promptly Produced IFNγ TNFα and GrzB in Response to IL-33 + IL-12 Stimulation
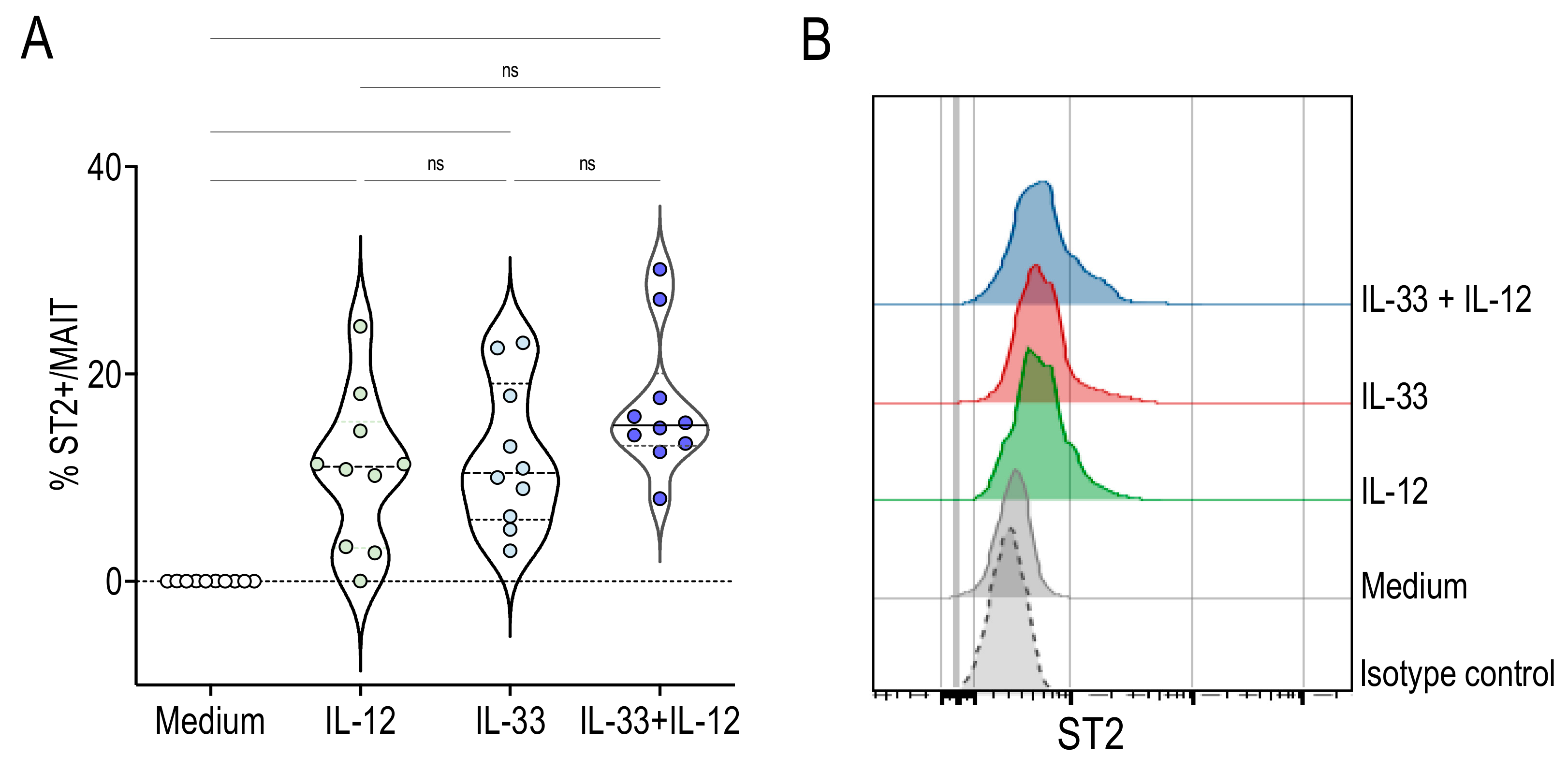
2.3. Human MAIT Cells Express the IL-33 Receptor ST2 in Response to IL-33 + IL-12 Stimulation
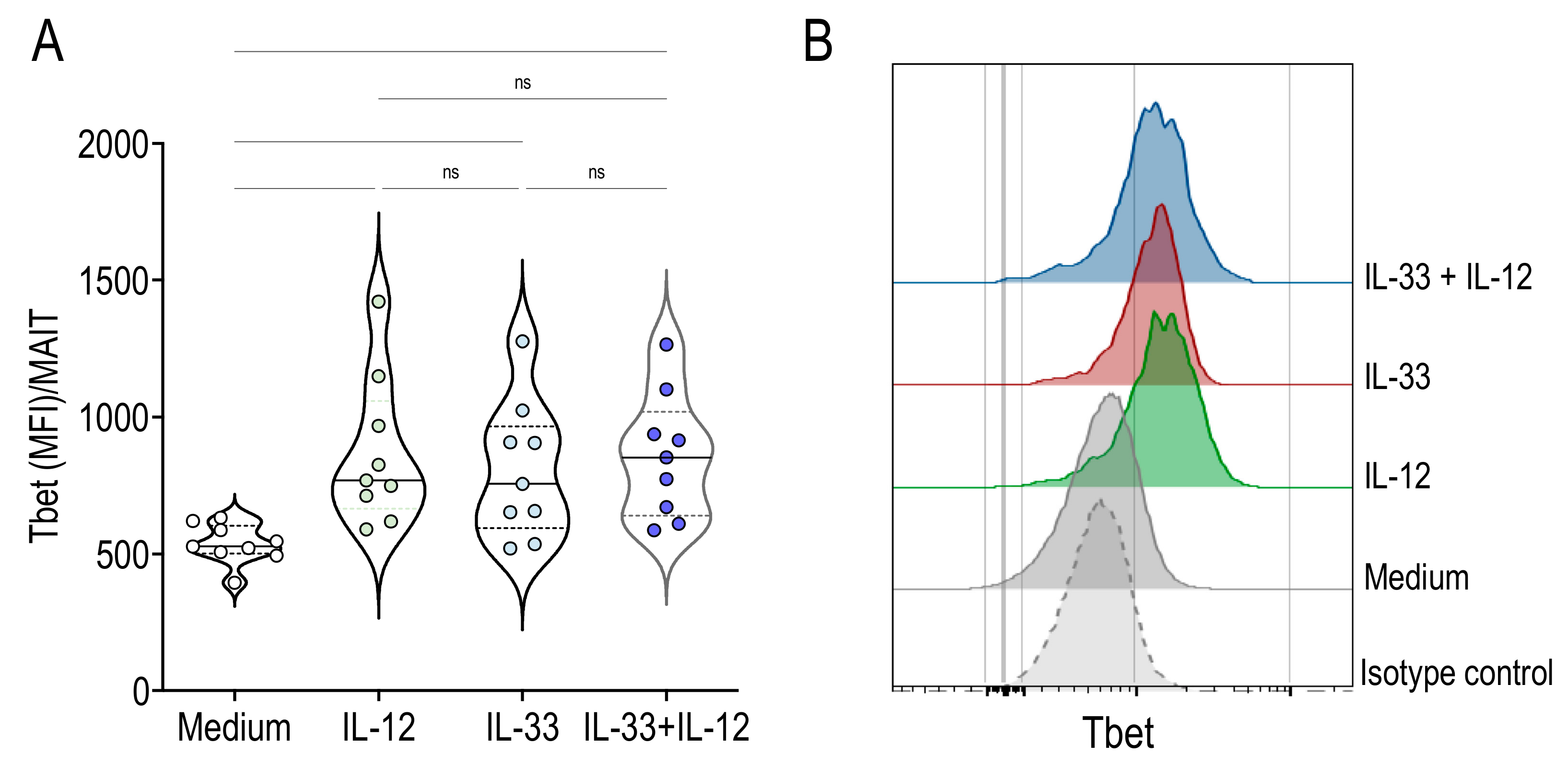
2.4. Activation of MAIT Cells by IL-33 + IL-12 Resulted in High Expression of Tbet
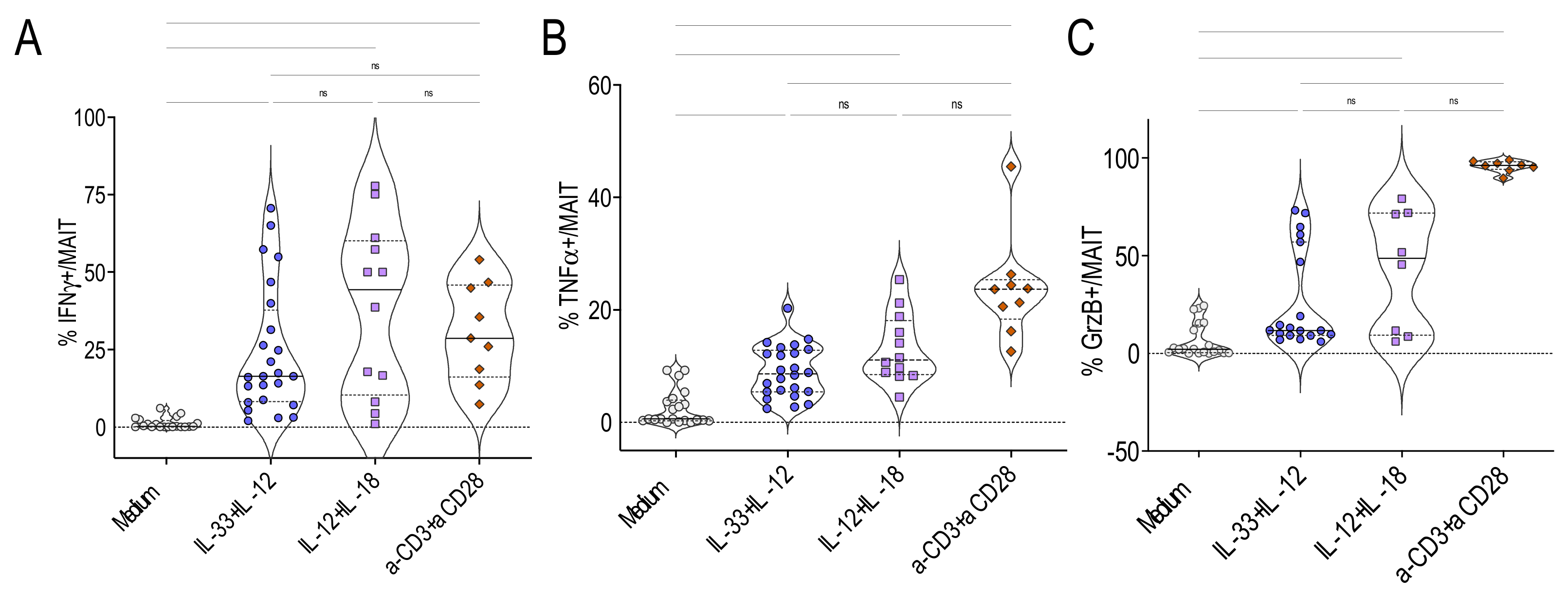
2.5. IL-33+ IL-12 Did Not Require Further TCR Cross-Linking to Activate MAIT Cells
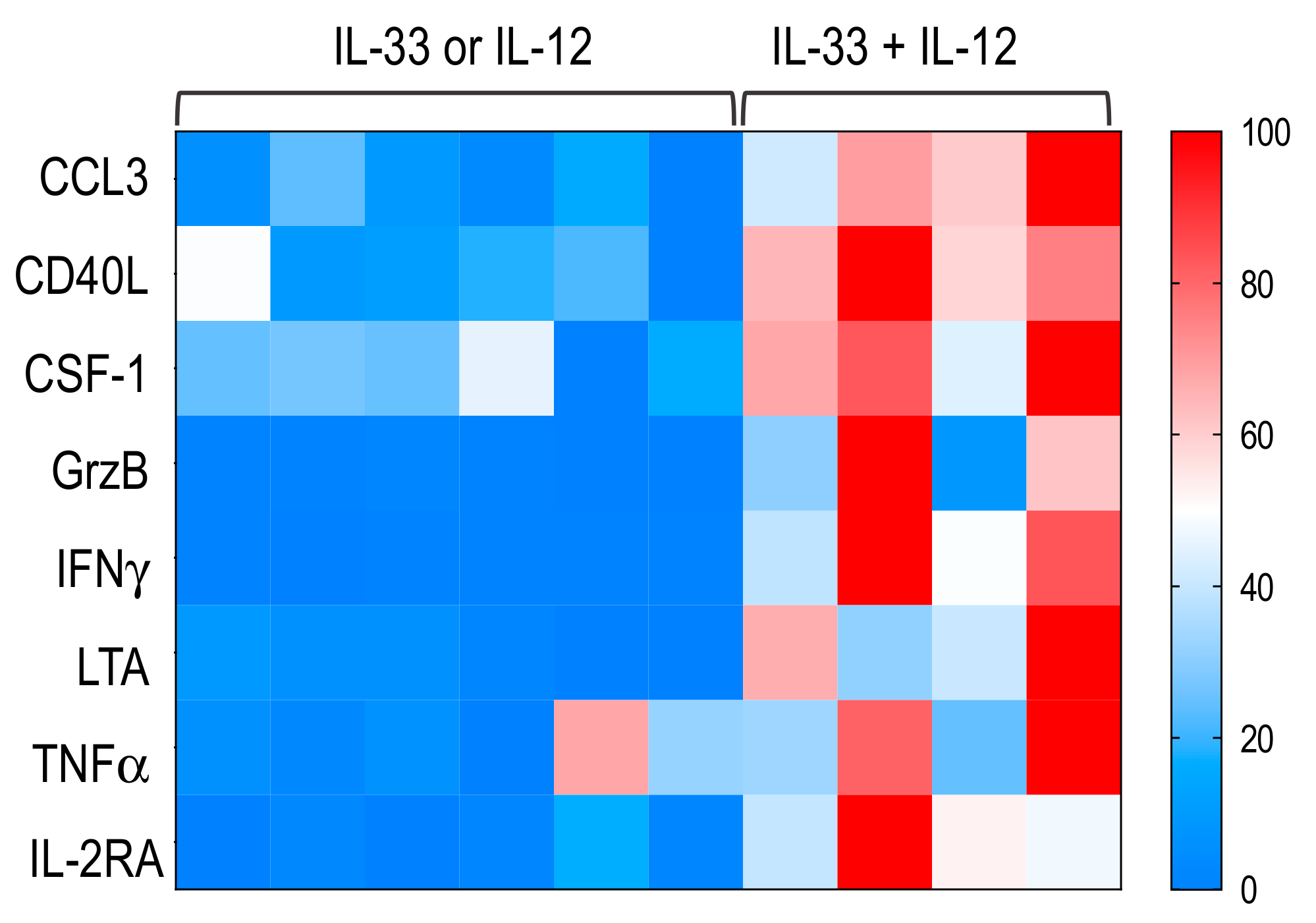
2.6. MAIT Cells Activated by IL-33+ IL-12 Expressed a Pro-Th1 Profile
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. In Vitro Culture
4.3. Flow Cytometry
4.4. Cell Sorting and Cytokine Measure
4.5. Real-Time Quantitative PCR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfrey, D.I.; Koay, H.F.; McCluskey, J.; Gherardin, N.A. The biology and functional importance of MAIT cells. Nat. Immunol. 2019, 20, 1110–1128. [Google Scholar] [CrossRef] [PubMed]
- Legoux, F.; Salou, M.; Lantz, O. MAIT Cell Development and Functions: The Microbial Connection. Immunity 2020, 53, 710–723. [Google Scholar] [CrossRef]
- Victor, J.R.; Lezmi, G.; Leite-de-Moraes, M. New Insights into Asthma Inflammation: Focus on iNKT, MAIT, and gammadeltaT Cells. Clin. Rev. Allergy Immunol. 2020, 59, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Toubal, A.; Nel, I.; Lotersztajn, S.; Lehuen, A. Mucosal-associated invariant T cells and disease. Nat. Rev. Immunol. 2019, 19, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Treiner, E.; Duban, L.; Bahram, S.; Radosavljevic, M.; Wanner, V.; Tilloy, F.; Affaticati, P.; Gilfillan, S.; Lantz, O. Selection of evolutionarily conserved mucosal-associated invariant T cells by MR1. Nature 2003, 422, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Kjer-Nielsen, L.; Patel, O.; Corbett, A.J.; Le Nours, J.; Meehan, B.; Liu, L.; Bhati, M.; Chen, Z.; Kostenko, L.; Reantragoon, R.; et al. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature 2012, 491, 717–723. [Google Scholar] [CrossRef]
- Le Bourhis, L.; Dusseaux, M.; Bohineust, A.; Bessoles, S.; Martin, E.; Premel, V.; Core, M.; Sleurs, D.; Serriari, N.E.; Treiner, E.; et al. MAIT cells detect and efficiently lyse bacterially-infected epithelial cells. PLoS Pathog. 2013, 9, e1003681. [Google Scholar] [CrossRef] [Green Version]
- Wong, E.B.; Ndung’u, T.; Kasprowicz, V.O. The role of mucosal-associated invariant T cells in infectious diseases. Immunology 2017, 150, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Ussher, J.E.; Bilton, M.; Attwod, E.; Shadwell, J.; Richardson, R.; de Lara, C.; Mettke, E.; Kurioka, A.; Hansen, T.H.; Klenerman, P.; et al. CD161++ CD8+ T cells, including the MAIT cell subset, are specifically activated by IL-12+IL-18 in a TCR-independent manner. Eur. J. Immunol. 2014, 44, 195–203. [Google Scholar] [CrossRef]
- Leite-De-Moraes, M.C.; Hameg, A.; Arnould, A.; Machavoine, F.; Koezuka, Y.; Schneider, E.; Herbelin, A.; Dy, M. A distinct IL-18-induced pathway to fully activate NK T lymphocytes independently from TCR engagement. J. Immunol. 1999, 163, 5871–5876. [Google Scholar]
- Brigl, M.; Bry, L.; Kent, S.C.; Gumperz, J.E.; Brenner, M.B. Mechanism of CD1d-restricted natural killer T cell activation during microbial infection. Nat. Immunol. 2003, 4, 1230–1237. [Google Scholar] [CrossRef]
- Bourgeois, E.; Van, L.P.; Samson, M.; Diem, S.; Barra, A.; Roga, S.; Gombert, J.M.; Schneider, E.; Dy, M.; Gourdy, P.; et al. The pro-Th2 cytokine IL-33 directly interacts with invariant NKT and NK cells to induce IFN-gamma production. Eur. J. Immunol. 2009, 39, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y.; Girard, J.P.; Turnquist, H.R. Interleukin-33 in health and disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef]
- Cayrol, C.; Girard, J.P. Interleukin-33 (IL-33): A nuclear cytokine from the IL-1 family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef]
- Kelly, J.; Minoda, Y.; Meredith, T.; Cameron, G.; Philipp, M.S.; Pellicci, D.G.; Corbett, A.J.; Kurts, C.; Gray, D.H.; Godfrey, D.I.; et al. Chronically stimulated human MAIT cells are unexpectedly potent IL-13 producers. Immunol. Cell Biol. 2019, 97, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Wei, G.; Zhu, J.; Liao, W.; Leonard, W.J.; Zhao, K.; Paul, W. IL-1 family members and STAT activators induce cytokine production by Th2, Th17, and Th1 cells. Proc. Natl. Acad. Sci. USA 2009, 106, 13463–13468. [Google Scholar] [CrossRef] [Green Version]
- Ioannidis, M.; Cerundolo, V.; Salio, M. The Immune Modulating Properties of Mucosal-Associated Invariant T Cells. Front. Immunol. 2020, 11, 1556. [Google Scholar] [CrossRef]
- Suliman, S.; Murphy, M.; Musvosvi, M.; Gela, A.; Meermeier, E.W.; Geldenhuys, H.; Hopley, C.; Toefy, A.; Bilek, N.; Veldsman, A.; et al. MR1-Independent Activation of Human Mucosal-Associated Invariant T Cells by Mycobacteria. J. Immunol. 2019, 203, 2917–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesteadt, E.; Zhang, I.; Yu, H.; Meierovics, A.; Chua Yankelevich, W.J.; Cowley, S. Interleukin-18 Is Critical for Mucosa-Associated Invariant T Cell Gamma Interferon Responses to Francisella Species In Vitro but Not In Vivo. Infect. Immun. 2018, 86, e00117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arend, W.P.; Palmer, G.; Gabay, C. IL-1, IL-18, and IL-33 families of cytokines. Immunol. Rev. 2008, 223, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.A. The IL-1 family of cytokines and receptors in rheumatic diseases. Nat. Rev. Rheumatol. 2019, 15, 612–632. [Google Scholar] [CrossRef]
- Komai-Koma, M.; Xu, D.; Li, Y.; McKenzie, A.N.; McInnes, I.B.; Liew, F.Y. IL-33 is a chemoattractant for human Th2 cells. Eur. J. Immunol. 2007, 37, 2779–2786. [Google Scholar] [CrossRef]
- Lezmi, G.; Abou-Taam, R.; Garcelon, N.; Dietrich, C.; Machavoine, F.; Delacourt, C.; Adel-Patient, K.; Leite-de-Moraes, M. Evidence for a MAIT-17-high phenotype in children with severe asthma. J. Allergy Clin. Immunol. 2019, 144, 1714–1716.e6. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Liu, M.; Wang, J.; Fan, H.; Yang, D.; Zhang, L.; Gu, X.; Nie, J.; Chen, Z.; Corbett, A.J.; et al. IL-17 production by tissue-resident MAIT cells is locally induced in children with pneumonia. Mucosal Immunol. 2020, 13, 824–835. [Google Scholar] [CrossRef] [Green Version]
- Luo, P.; Deng, S.; Ye, H.; Yu, X.; Deng, Q.; Zhang, Y.; Jiang, L.; Li, J.; Yu, Y.; Han, W. The IL-33/ST2 pathway suppresses murine colon cancer growth and metastasis by upregulating CD40 L signaling. Biomed. Pharmacother. 2020, 127, 110232. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Villarino, G.; Maldonado-Lopez, R.; Possemato, R.; Penaranda, C.; Glimcher, L.H. T-bet is required for optimal production of IFN-gamma and antigen-specific T cell activation by dendritic cells. Proc. Natl. Acad. Sci. USA 2003, 100, 7749–7754. [Google Scholar] [CrossRef] [Green Version]
- Le Bourhis, L.; Martin, E.; Peguillet, I.; Guihot, A.; Froux, N.; Core, M.; Levy, E.; Dusseaux, M.; Meyssonnier, V.; Premel, V.; et al. Antimicrobial activity of mucosal-associated invariant T cells. Nat. Immunol. 2010, 11, 701–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazarian, L.; Caillat-Zucman, S.; Houdouin, V. Mucosal-Associated Invariant T Cell Interactions with Commensal and Pathogenic Bacteria: Potential Role in Antimicrobial Immunity in the Child. Front. Immunol. 2017, 8, 1837. [Google Scholar] [CrossRef] [Green Version]
- Trottein, F.; Paget, C. Natural Killer T Cells and Mucosal-Associated Invariant T Cells in Lung Infections. Front. Immunol. 2018, 9, 1750. [Google Scholar] [CrossRef] [PubMed]
- Ussher, J.E.; Willberg, C.B.; Klenerman, P. MAIT cells and viruses. Immunol. Cell Biol. 2018, 96, 630–641. [Google Scholar] [CrossRef]
- van Wilgenburg, B.; Scherwitzl, I.; Hutchinson, E.C.; Leng, T.; Kurioka, A.; Kulicke, C.; de Lara, C.; Cole, S.; Vasanawathana, S.; Limpitikul, W.; et al. MAIT cells are activated during human viral infections. Nat. Commun 2016, 7, 11653. [Google Scholar] [CrossRef] [Green Version]
- van Wilgenburg, B.; Loh, L.; Chen, Z.; Pediongco, T.J.; Wang, H.; Shi, M.; Zhao, Z.; Koutsakos, M.; Nussing, S.; Sant, S.; et al. MAIT cells contribute to protection against lethal influenza infection in vivo. Nat. Commun 2018, 9, 4706. [Google Scholar] [CrossRef] [Green Version]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Zizzo, G.; Cohen, P.L. Imperfect storm: Is interleukin-33 the Achilles heel of COVID-19? Lancet Rheumatol. 2020, 2, e779–e790. [Google Scholar] [CrossRef]
- Munitz, A.; Edry-Botzer, L.; Itan, M.; Tur-Kaspa, R.; Dicker, D.; Marcoviciu, D.; Goren, M.G.; Mor, M.; Lev, S.; Gottesman, T.; et al. Rapid seroconversion and persistent functional IgG antibodies in severe COVID-19 patients correlates with an IL-12p70 and IL-33 signature. Sci. Rep. 2021, 11, 3461. [Google Scholar] [CrossRef]
- Flament, H.; Rouland, M.; Beaudoin, L.; Toubal, A.; Bertrand, L.; Lebourgeois, S.; Rousseau, C.; Soulard, P.; Gouda, Z.; Cagninacci, L.; et al. Outcome of SARS-CoV-2 infection is linked to MAIT cell activation and cytotoxicity. Nat. Immunol. 2021, 22, 322–335. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azzout, M.; Dietrich, C.; Machavoine, F.; Gastineau, P.; Bottier, A.; Lezmi, G.; Leite-de-Moraes, M. IL-33 Enhances IFNγ and TNFα Production by Human MAIT Cells: A New Pro-Th1 Effect of IL-33. Int. J. Mol. Sci. 2021, 22, 10602. https://doi.org/10.3390/ijms221910602
Azzout M, Dietrich C, Machavoine F, Gastineau P, Bottier A, Lezmi G, Leite-de-Moraes M. IL-33 Enhances IFNγ and TNFα Production by Human MAIT Cells: A New Pro-Th1 Effect of IL-33. International Journal of Molecular Sciences. 2021; 22(19):10602. https://doi.org/10.3390/ijms221910602
Chicago/Turabian StyleAzzout, Mourad, Céline Dietrich, François Machavoine, Pauline Gastineau, Alix Bottier, Guillaume Lezmi, and Maria Leite-de-Moraes. 2021. "IL-33 Enhances IFNγ and TNFα Production by Human MAIT Cells: A New Pro-Th1 Effect of IL-33" International Journal of Molecular Sciences 22, no. 19: 10602. https://doi.org/10.3390/ijms221910602