
Genome-Wide Identification and Expression Analysis of the Aux/IAA and Auxin Response Factor Gene Family in Medicago truncatula


Abstract
:1. Introduction
2. Results
2.1. Identification of IAA and ARF Members in M. truncatula
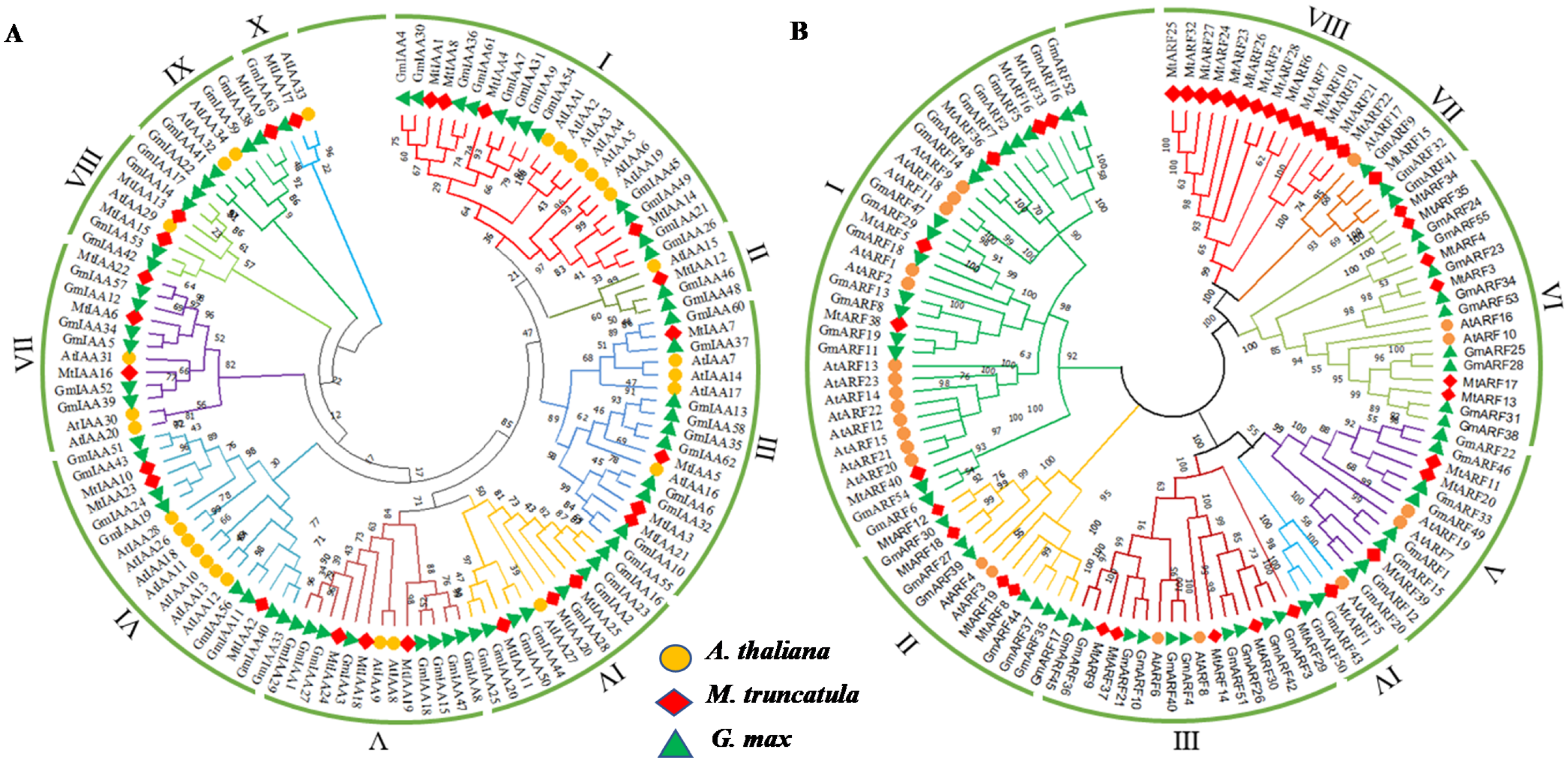
2.2. Phylogenetic Analysis of IAAs and ARFs among M. truncatula, Soybean and Arabidopsis
2.3. Gene Structure and Domain Architecture
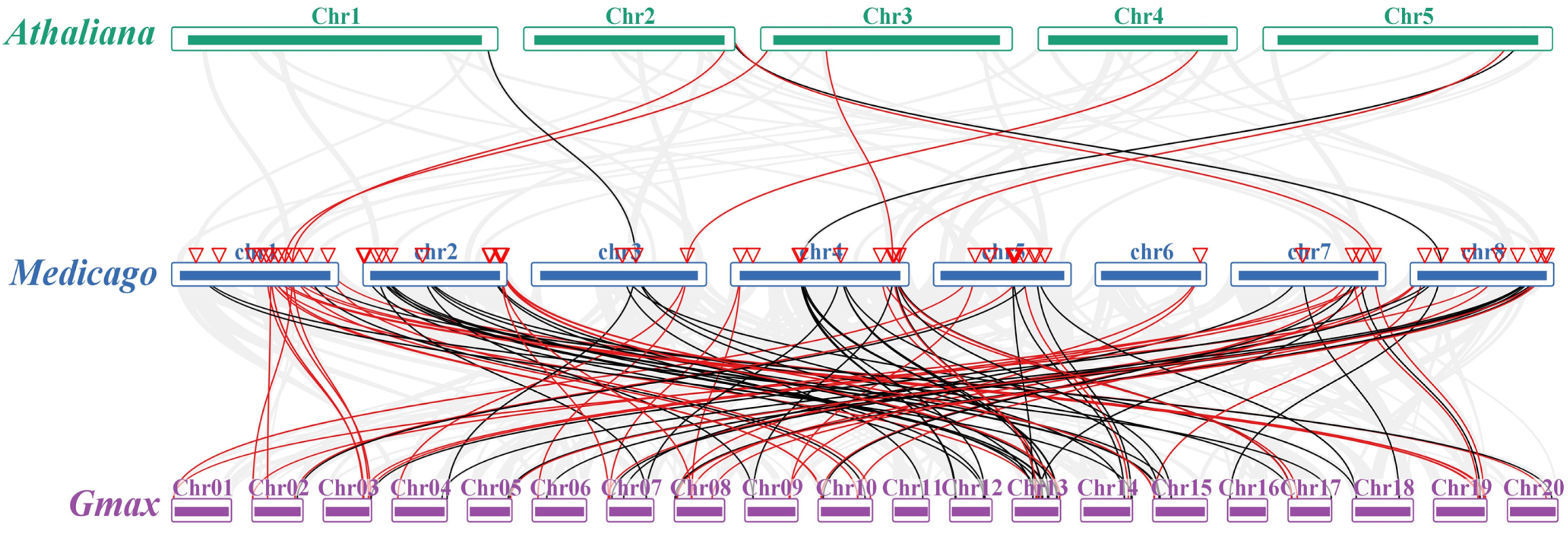
2.4. Chromosomal Location and Synteny Analysis of MtIAA and MtARF Genes
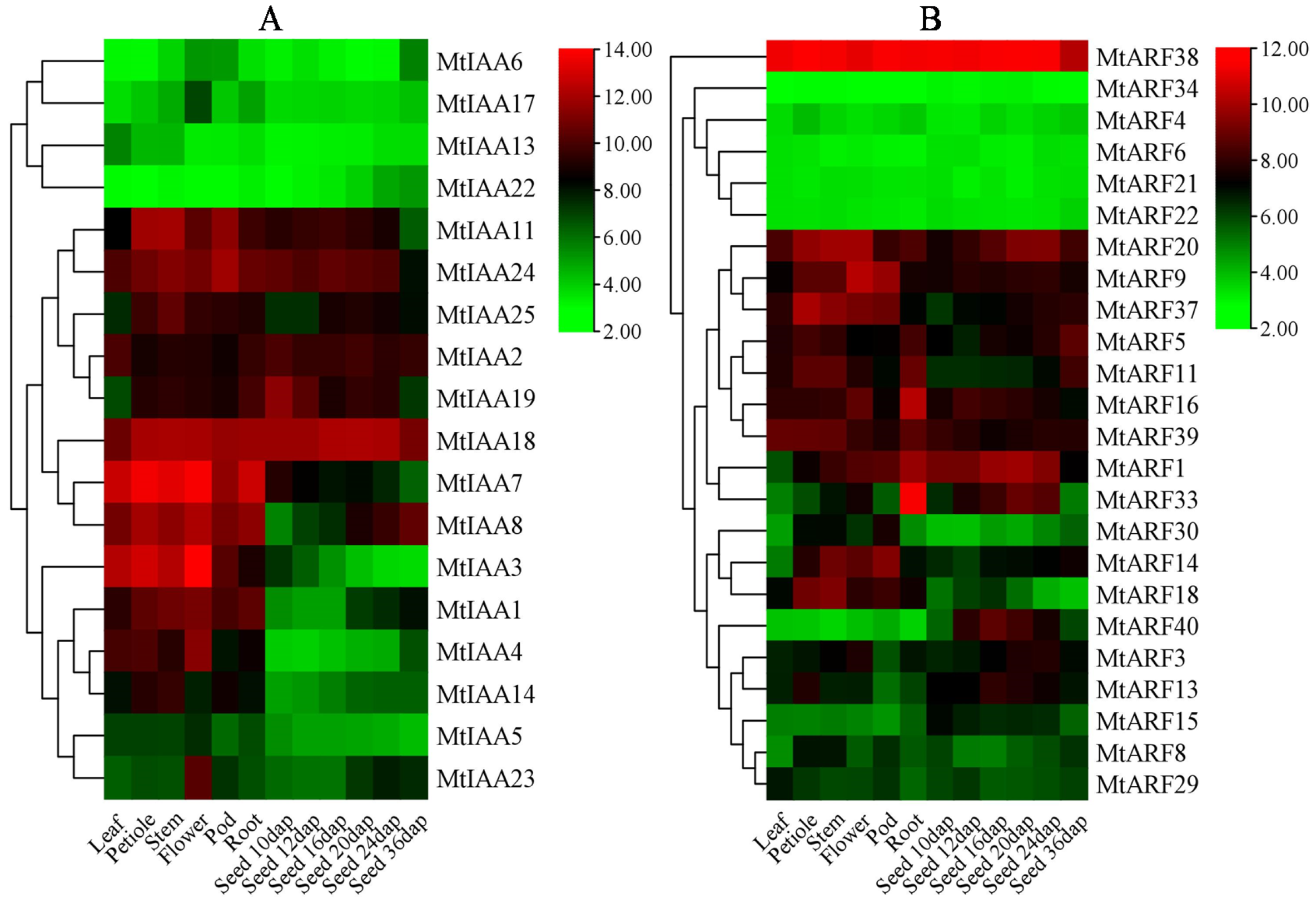
2.5. Spatial and Temporal Expression of MtIAAs and MtARFs
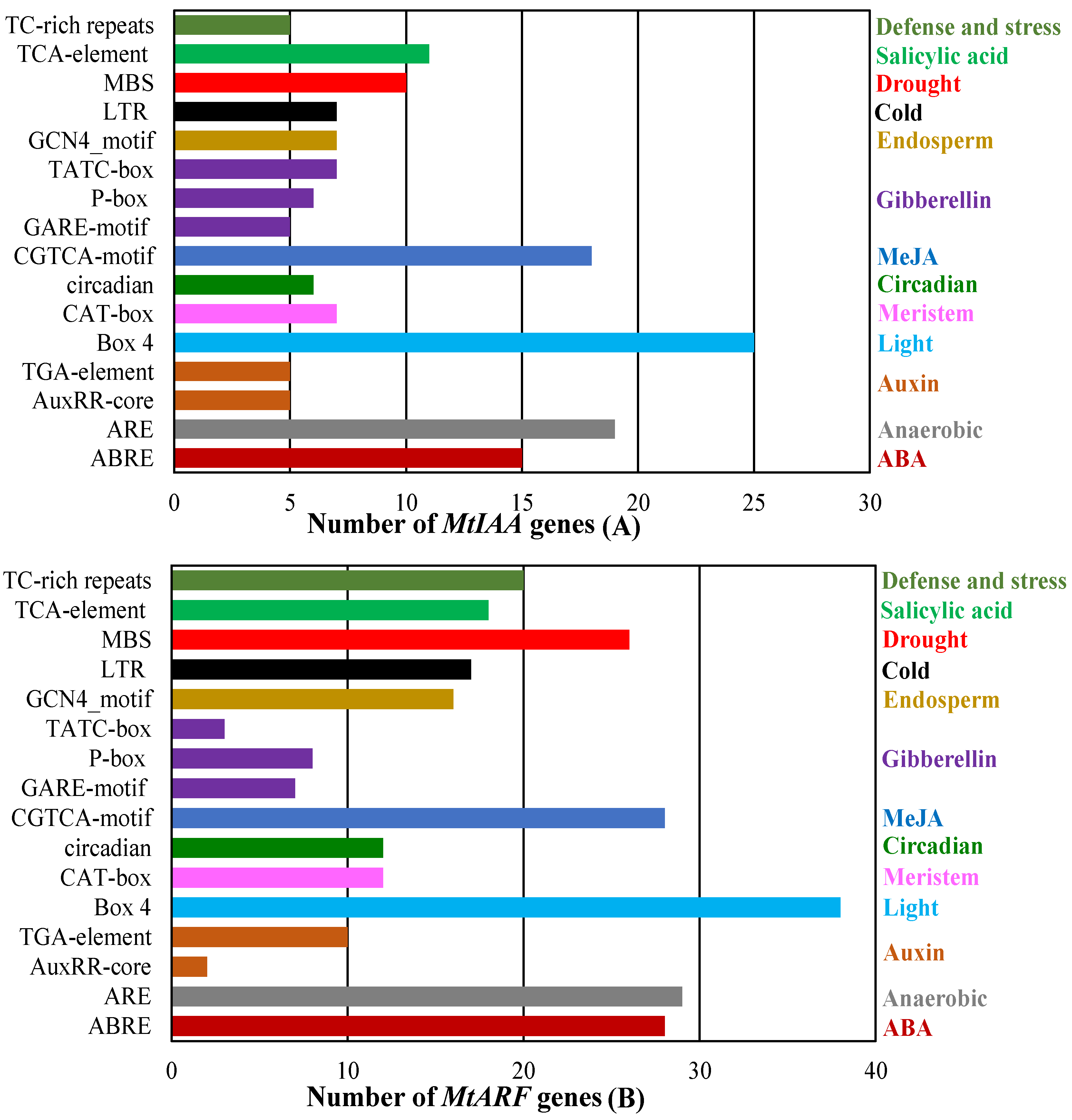
2.6. Analysis of cis-Acting Element in the Promoter Region of MtIAA and MtARF Genes
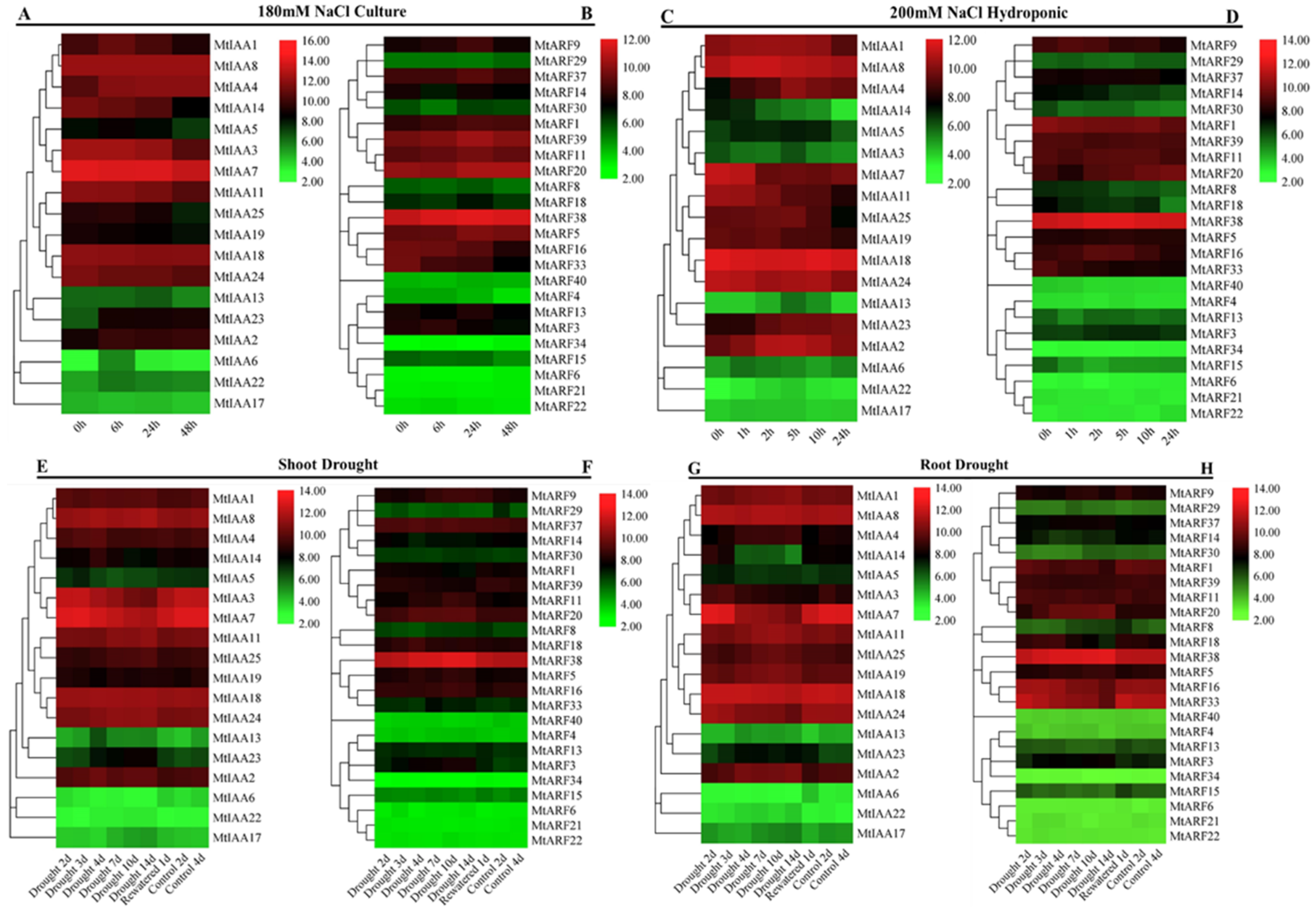
2.7. Responses of MtIAAs and MtARFs to Salt, Drought and Cold
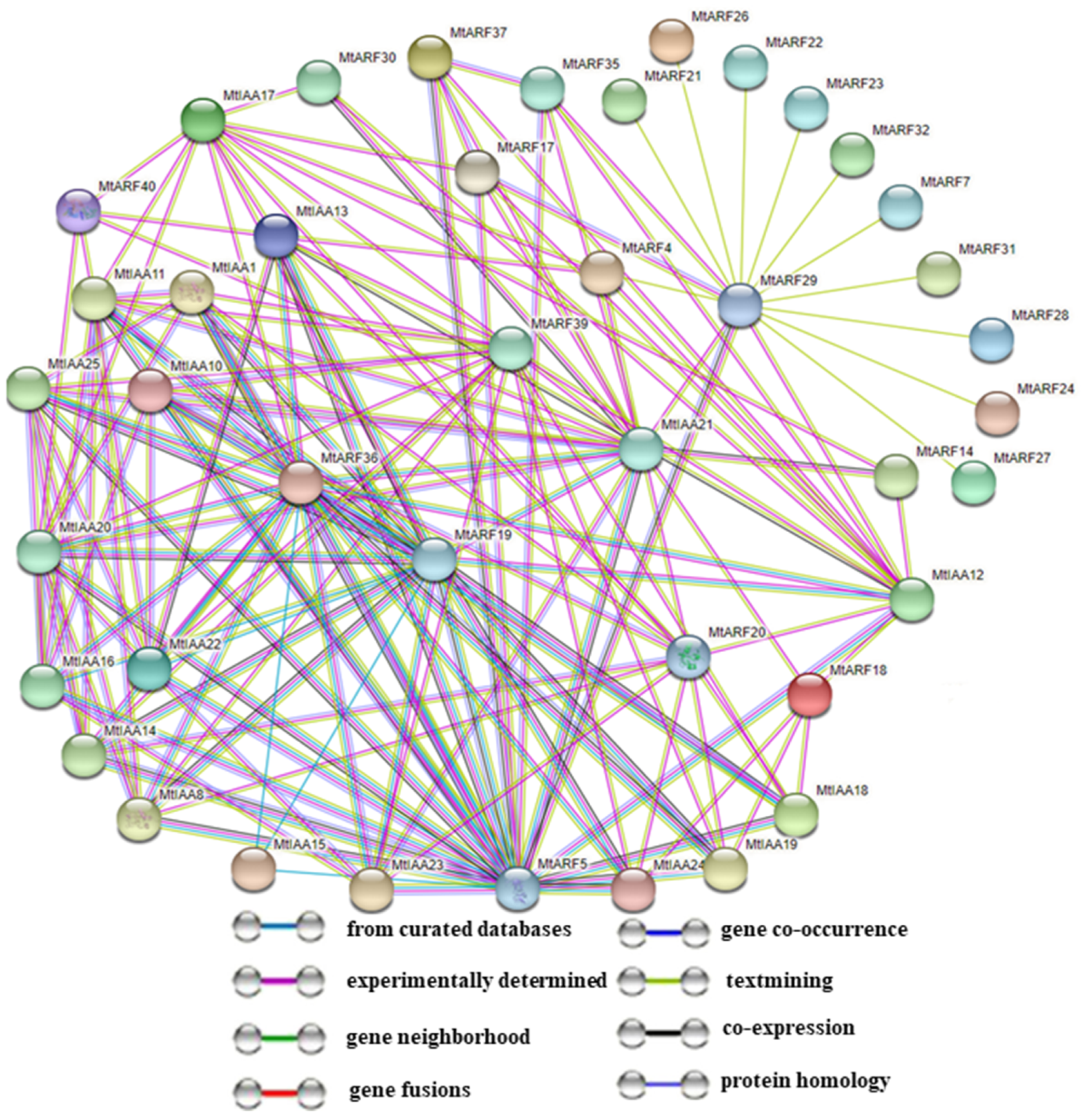
2.8. Predicted MtIAA and MtARF Family Interaction Networks
3. Discussion
4. Material and Methods
4.1. Identification of IAA and ARF Genes in M. truncatula
4.2. Analysis of Conserved Domain, Gene Structure and Characterization of MtIAA and MtARF Genes
4.3. Phylogenetic Relationships of MtIAA and MtARF Proteins in M. truncatula, Arabidopsis and Soybean
4.4. Chromosomal Locations of MtIAA and MtARF Genes
4.5. Analysis of cis-Acting Elements of MtIAA and MtARF Genes
4.6. Analysis of Microarray Expression Profile
4.7. Analysis of Relative Expression of MtIAAs and MtARFs in Response to Cold
4.8. Predicted Protein Interaction Network and Co-Expression Network Construction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.H.; Maharjan, P.M.; Lee, O.; Fujioka, S.; Jang, S.Y.; Kim, B.; Takatsuto, S.; Tsujimoto, M.; Kim, H.; Cho, S.; et al. Auxin stimulates DWARF4 expression and brassinosteroid biosynthesis in Arabidopsis. Plant J. 2011, 66, 564–578. [Google Scholar] [CrossRef]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef]
- Dreher, K.A.; Brown, J.; Saw, R.E.; Callis, J. The Arabidopsis Aux/IAA protein family has diversified in degradation and auxin responsiveness. Plant Cell 2006, 18, 699–714. [Google Scholar] [CrossRef] [Green Version]
- Lavy, M.; Estelle, M. Mechanisms of auxin signaling. Development 2016, 143, 3226–3229. [Google Scholar] [CrossRef] [Green Version]
- Overvoorde, P.J.; Okushima, Y.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Liu, A.; Onodera, C.; Quach, H.; et al. Functional genomic analysis of the AUXIN/INDOLE-3-ACETIC ACID gene family members in Arabidopsis thaliana. Plant Cell 2005, 17, 3282–3300. [Google Scholar] [CrossRef] [Green Version]
- Szemenyei, H.; Hannon, M.; Long, J.A. Topless mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 2008, 319, 1384–1386. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.A.; Zenser, N.; Leyser, O.; Callis, J. Rapid degradation of auxin/indoleacetic acid proteins requires conserved amino acids of domain II and is proteasome dependent. Plant Cell 2001, 13, 2349–2360. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W. Auxin response factors. Plant Cell Environ. 2016, 39, 1014–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [Green Version]
- De Smet, I.; Jürgens, G. Patterning the axis in plants–auxin in control. Curr. Opin. Genet. Dev. 2007, 17, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Manners, J.M. Linking development to defense: Auxin in plant–pathogen interactions. Trends Plant Sci. 2009, 14, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Uberti-Manassero, N.G.; Lucero, L.E.; Viola, I.L.; Vegetti, A.C.; Gonzalez, D.H. The class I protein AtTCP15 modulates plant development through a pathway that overlaps with the one affected by CIN-like TCP proteins. J. Exp. Bot. 2012, 63, 809–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakehal, A.; Chaabouni, S.; Cavel, E.; Le Hir, R.; Ranjan, A.; Raneshan, Z.; Novák, O.; Păcurar, D.I.; Perrone, I.; Jobert, F.; et al. A molecular framework for the control of adventitious rooting by TIR1/AFB2-Aux/IAA-dependent auxin signaling in Arabidopsis. Mol. Plant 2019, 12, 1499–1514. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.S.; Yu, Q.Q.; Liu, J.J.; Wen, X.J.; Yan, Z.W.; Hu, K.Q.; Li, H.B.; Kong, X.P.; Li, C.L.; Tian, H.Y.; et al. Non-canonical AUX/IAA protein IAA33 competes with canonical AUX/IAA repressor IAA5 to negatively regulate auxin signaling. EMBO J. 2020, 39, e101515. [Google Scholar] [CrossRef]
- Audran-Delalande, C.; Bassa, C.; Mila, I.; Regad, F.; Zouine, M.; Bouzayen, M. Genome-wide identification, functional analysis and expression profiling of the Aux/IAA gene family in tomato. Plant Cell Physiol. 2012, 53, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Gan, D.; Zhuang, D.; Ding, F.; Yu, Z.; Zhao, Y. Identification and expression analysis of primary auxin-responsive Aux/IAA gene family in cucumber (Cucumis sativus). J. Genet. 2013, 92, 513–521. [Google Scholar] [CrossRef]
- Ludwig, Y.; Zhang, Y.; Hochholdinger, F. The maize (Zea mays L.) AUXIN/INDOLE-3- ACETIC ACID gene family: Phylogeny, synteny, and unique root-type and tissue-specific expression patterns during development. PLoS ONE 2013, 8, e78859. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Kaur, N.; Garg, R.; Thakur, J.K.; Tyagi, A.K.; Khurana, J.P. Structure and expression analysis of early auxin-responsive Aux/IAA gene family in rice (Oryza sativa). Funct. Integr. Genom. 2006, 6, 47–59. [Google Scholar] [CrossRef]
- Okushima, Y.; Overvoorde, P.J.; Arima, K.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Lui, A.; Nguyen, D.; et al. Functional genomic analysis of the auxin response factor gene family members in Arabidopsis thaliana: Unique and overlapping functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.D.; Yuan, C.C.; Li, H.L.; Lin, W.H.; Yang, Y.J.; Shen, C.J.; Zheng, X.L. Genome-wide identification and characterization of auxin response factor (ARF) family genes related to flower and fruit development in papaya (Carica papaya L.). BMC Genom. 2015, 16, 901. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.J.; Wang, S.K.; Bai, Y.H.; Wu, Y.R.; Zhang, S.N.; Chen, M.; Guilfoyle, T.J.; Wu, P.; Qi, Y.H. Functional analysis of the structural domain of ARF proteins in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 3971–3981. [Google Scholar] [CrossRef] [Green Version]
- Roosjen, M.; Paque, S.; Weijers, D. Auxin response factors: Output control in auxin biology. J. Exp. Bot. 2018, 69, 179–188. [Google Scholar] [CrossRef]
- Liu, Z.N.; Miao, L.M.; Huo, R.X.; Song, X.Y.; Johnson, C.; Kong, L.J.; Sundaresan, V.; Yu, X.L. ARF2–ARF4 and ARF5 are essential for female and male gametophyte development in Arabidopsis. Plant Cell Physiol. 2018, 59, 179–189. [Google Scholar] [CrossRef]
- Ghelli, R.; Brunetti, P.; Napoli, N.; De Paolis, A.; Cecchetti, V.; Tsuge, T.; Serino, G.; Matsui, M.; Mele, G.; Rinaldi, G.; et al. A newly identified flower-specific splice variant of AUXIN RESPONSE FACTOR8 regulates stamen elongation and endothecium lignification in Arabidopsis. Plant Cell 2018, 30, 620–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Zhang, K.; Guo, L.; Liu, X.; Zhang, Z. AUXIN RESPONSE FACTOR3 plays distinct role during early flower development. Plant Signal. Behav. 2018, 13, e1467690. [Google Scholar] [CrossRef] [Green Version]
- Le, B.; Nawaz, M.A.; Rehman, H.M.; Le, T.; Yang, S.H.; Golokhvast, K.S.; Son, E.; Chung, G. Genome-wide characterization and expression pattern of auxin response factor (ARF) gene family in soybean and common bean. Genes Genom. 2016, 38, 1165–1178. [Google Scholar] [CrossRef]
- Xing, H.Y.; Pudake, R.N.; Guo, G.G.; Xing, G.F.; Hu, Z.R.; Zhang, Y.R.; Sun, Q.X.; Ni, Z. Genome-wide identification and expression profiling of auxin response factor (ARF) gene family in maize. BMC Genom. 2011, 12, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Hao, L.; Zhao, P.; Xu, Y.; Zhong, N.; Zhang, H.; Liu, N. Genome-wide identification, expression profiling and evolutionary analysis of auxin response factor gene family in potato (Solanum tuberosum group phureja). Sci. Rep. 2019, 9, 1–13. [Google Scholar]
- Zhou, X.J.; Wu, X.Z.; Li, T.J.; Jia, M.L.; Liu, X.S.; Zou, Y.L.; Liu, Z.X.; Wen, F. Identification, characterization, and expression analysis of auxin response factor (ARF) gene family in Brachypodium distachyon. Funct. Integr. Genom. 2018, 18, 709–724. [Google Scholar] [CrossRef]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Roe, B.A. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.J.; Yue, R.Q.; Yang, Y.J.; Zhang, L.; Sun, T.; Xu, L.Q.; Tie, S.G.; Wang, H.Z. Genome-wide identification and expression profiling analysis of the Aux/IAA gene family in Medicago truncatula during the early phase of Sinorhizobium meliloti infection. PLoS ONE 2014, 9, e107495. [Google Scholar]
- Shen, C.J.; Yue, R.Q.; Sun, T.; Zhang, L.; Xu, L.Q.; Tie, S.G.; Wang, H.Z.; Yang, Y.J. Genome-wide identification and expression analysis of auxin response factor gene family in Medicago truncatula. Front. Plant Sci. 2015, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Jain, M. Genome-wide survey and comprehensive expression profiling of Aux/IAA gene family in chickpea and soybean. Front. Plant Sci. 2015, 6, 918. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, B.; Zhang, Q.; Wang, J.; King, G.J.; Liu, K. Genome-wide analysis of the auxin/indoleacetic acid (Aux/IAA) gene family in allotetraploid rapeseed (Brassica napus L.). BMC Plant Biol. 2017, 17, 204. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.W. Roles and activities of Aux/IAA proteins in Arabidopsis. Trends Plant Sci. 2001, 6, 420–425. [Google Scholar] [CrossRef]
- Li, D.; Su, Z.; Dong, J.; Wang, T. An expression database for roots of the model legume Medicago truncatula under salt stress. BMC Genom. 2009, 10, 517. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wu, M.; Liu, H.; Gao, Y.; Xiang, Y. Systematic identification and expression pattern analysis of the Aux/IAA and ARF gene families in moso bamboo (Phyllostachys edulis). Plant Physiol. Biochem. 2018, 130, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.Q.; Zheng, R.; Ge, D.F.; Zheng, W.; Wang, X.H.; Gu, Y.T.; Gelová, Z.; et al. TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nature 2019, 568, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Kubeš, M.; Napier, R. Non-canonical auxin signalling: Fast and curious. J. Exp. Bot. 2019, 70, 2609. [Google Scholar] [CrossRef] [PubMed]
- Mai, Y.X.; Wang, L.; Yang, H.Q. A gain-of-function mutation in IAA7/AXR2 confers late flowering under short-day light in Arabidopsis F. J. Integr. Plant Biol. 2011, 53, 480–492. [Google Scholar] [CrossRef]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.Y.; Yang, X.; Lu, J.; Song, F.Y.; Sun, J.H.; Wang, C.; Lian, J.; Zhao, L.L.; Zhao, B.C. OsIAA20, an Aux/IAA protein, mediates abiotic stress tolerance in rice through an ABA pathway. Plant Sci. 2021, 308, 110903. [Google Scholar] [CrossRef]
- Zouine, M.; Fu, Y.Y.; Chateigner-Boutin, A.L.; Mila, I.; Frasse, P.; Wang, H.; Audran, C.; Roustan, J.P.; Bouzayen, M. Characterization of the tomato ARF gene family uncovers a multi-levels post-transcriptional regulation including alternative splicing. PLoS ONE 2014, 9, e84203. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, D.K.; Do Choi, Y.; Kim, J.K. OsIAA6, a member of the rice Aux/IAA gene family, is involved in drought tolerance and tiller outgrowth. Plant Sci. 2015, 236, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Bai, Y.H.; Shen, C.J.; Wu, Y.R.; Zhang, S.N.; Jiang, D.A.; Guilfoyle, T.J.; Chen, M.; Qi, Y.H. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genom. 2010, 10, 533–546. [Google Scholar] [CrossRef]
- Piya, S.; Shrestha, S.K.; Binder, B.; Stewart Jr, C.N.; Hewezi, T. Protein-protein interaction and gene co-expression maps of ARFs and Aux/IAAs in Arabidopsis. Front. Plant Sci. 2014, 5, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Agarwal, P.; Pareek, A.; Tyagi, A.K.; Sharma, A.K. Genomic survey, gene expression, and interaction analysis suggest diverse roles of ARF and Aux/IAA proteins in Solanaceae. Plant Mol. Biol. Rep. 2015, 33, 1552–1572. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shi, H.F.; Huang, R.S.; Ye, R.; Luo, Y.R.; Guo, Z.F.; Lu, S.Y. AIR12 confers cold tolerance through regulation of the CBF cold response pathway and ascorbate homeostasis. Plant Cell Environ. 2021, 44, 1522–1533. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Locus ID | ORF (bp) | A.A. | MW (KDa) | pI | GRAVY |
|---|---|---|---|---|---|---|
| MtIAA1 | Medtr1g040675 | 561 | 186 | 20.76 | 6.75 | −0.554 |
| MtIAA2 | Medtr1g069495 | 819 | 272 | 30.11 | 7.15 | −0.814 |
| MtIAA3 | Medtr1g070520 | 738 | 245 | 27.48 | 7.57 | −0.689 |
| MtIAA4 | Medtr1g070830 | 615 | 204 | 22.91 | 6.20 | −0.732 |
| MtIAA5 | Medtr1g080860 | 762 | 253 | 27.51 | 6.75 | −0.512 |
| MtIAA6 | Medtr1g085750 | 531 | 176 | 19.80 | 5.56 | −0.535 |
| MtIAA7 | Medtr1g093240 | 711 | 236 | 25.72 | 8.60 | −0.552 |
| MtIAA8 | Medtr1g093350 | 537 | 178 | 19.98 | 7.68 | −0.752 |
| MtIAA9 | Medtr1g109510 | 537 | 178 | 20.19 | 4.77 | −0.493 |
| MtIAA10 | Medtr2g100780 | 810 | 269 | 29.96 | 8.86 | −0.863 |
| MtIAA11 | Medtr2g101500 | 981 | 326 | 35.36 | 8.11 | −0.539 |
| MtIAA12 | Medtr2g102490 | 597 | 198 | 22.50 | 8.80 | −0.529 |
| MtIAA13 | Medtr3g106850 | 642 | 213 | 24.73 | 9.00 | −0.883 |
| MtIAA14 | Medtr4g011880 | 537 | 178 | 20.14 | 6.21 | −0.598 |
| MtIAA15 | Medtr4g115075 | 627 | 208 | 24.00 | 6.16 | −0.853 |
| MtIAA16 | Medtr4g124300 | 555 | 184 | 20.88 | 6.51 | −0.497 |
| MtIAA17 | Medtr4g128070 | 522 | 173 | 19.22 | 7.78 | −0.524 |
| MtIAA18 | Medtr5g030710 | 1008 | 335 | 36.26 | 8.52 | −0.468 |
| MtIAA19 | Medtr5g067350 | 1044 | 347 | 38.01 | 8.62 | −0.567 |
| MtIAA20 | Medtr6g488150 | 765 | 254 | 28.32 | 8.43 | −0.580 |
| MtIAA21 | Medtr7g096090 | 753 | 250 | 27.90 | 5.36 | −0.558 |
| MtIAA22 | Medtr7g110790 | 486 | 161 | 18.19 | 5.68 | −0.358 |
| MtIAA23 | Medtr8g014520 | 816 | 271 | 30.12 | 8.28 | −0.755 |
| MtIAA24 | Medtr8g067530 | 1071 | 356 | 38.67 | 6.38 | −0.455 |
| MtIAA25 | Medtr8g103030 | 882 | 293 | 31.89 | 8.04 | −0.509 |
| Name | Locus ID | ORF (bp) | A.A. | MW (KDa) | pI | GRAVY |
|---|---|---|---|---|---|---|
| MtARF1 | Medtr1g024025 | 3798 | 1265 | 141.16 | 5.26 | −0.439 |
| MtARF2 | Medtr1g058210 | 1380 | 459 | 51.30 | 8.32 | −0.383 |
| MtARF3 | Medtr1g064430 | 2070 | 689 | 76.95 | 6.69 | −0.475 |
| MtARF4 | Medtr1g094960 | 1860 | 619 | 68.96 | 7.53 | −0.477 |
| MtARF5 | Medtr2g005240 | 2016 | 671 | 74.98 | 5.71 | −0.570 |
| MtARF6 | Medtr2g006270 | 1371 | 456 | 51.92 | 8.67 | −0.426 |
| MtARF7 | Medtr2g006380 | 1371 | 456 | 52.06 | 6.92 | −0.464 |
| MtARF8 | Medtr2g014770 | 2049 | 682 | 74.20 | 6.11 | −0.323 |
| MtARF9 | Medtr2g018690 | 2727 | 908 | 100.83 | 6.14 | −0.480 |
| MtARF10 | Medtr2g024430 | 1548 | 515 | 57.55 | 6.20 | −0.372 |
| MtARF11 | Medtr2g043250 | 3345 | 1114 | 124.37 | 5.99 | −0.721 |
| MtARF12 | Medtr2g093740 | 2433 | 810 | 90.08 | 7.24 | −0.430 |
| MtARF13 | Medtr2g094570 | 2268 | 755 | 84.01 | 8.64 | −0.379 |
| MtARF14 | Medtr3g064050 | 2550 | 849 | 94.27 | 5.92 | −0.503 |
| MtARF15 | Medtr3g073420 | 1782 | 593 | 64.87 | 6.15 | −0.218 |
| MtARF16 | Medtr4g021580 | 2004 | 667 | 74.19 | 6.22 | −0.470 |
| MtARF17 | Medtr4g058930 | 2097 | 698 | 77.25 | 7.21 | −0.340 |
| MtARF18 | Medtr4g060460 | 2379 | 792 | 88.10 | 6.36 | −0.455 |
| MtARF19 | Medtr4g088210 | 2139 | 712 | 79.09 | 6.50 | −0.469 |
| MtARF20 | Medtr4g124900 | 3363 | 1120 | 125.41 | 6.08 | −0.687 |
| MtARF21 | Medtr5g040740 | 1233 | 410 | 45.10 | 8.42 | −0.414 |
| MtARF22 | Medtr5g040880 | 1575 | 524 | 58.20 | 6.33 | −0.385 |
| MtARF23 | Medtr5g060630 | 1572 | 523 | 58.09 | 5.14 | −0.407 |
| MtARF24 | Medtr5g060770 | 1071 | 356 | 39.51 | 5.20 | −0.264 |
| MtARF25 | Medtr5g060780 | 1200 | 399 | 44.40 | 6.02 | −0.321 |
| MtARF26 | Medtr5g061220 | 1587 | 528 | 58.55 | 6.37 | −0.363 |
| MtARF27 | Medtr5g061890 | 1200 | 399 | 44.14 | 5.86 | −0.390 |
| MtARF28 | Medtr5g062970 | 720 | 239 | 26.85 | 7.23 | −0.042 |
| MtARF29 | Medtr5g074840 | 492 | 163 | 18.06 | 8.87 | 0.258 |
| MtARF30 | Medtr5g076270 | 2526 | 841 | 93.28 | 5.93 | −0.472 |
| MtARF31 | Medtr5g082140 | 1383 | 460 | 51.65 | 6.47 | −0.431 |
| MtARF32 | Medtr5g460920 | 1200 | 399 | 44.45 | 5.86 | −0.306 |
| MtARF33 | Medtr7g062540 | 2043 | 680 | 76.17 | 5.74 | −0.545 |
| MtARF34 | Medtr7g101275 | 1218 | 405 | 46.08 | 8.10 | −0.491 |
| MtARF35 | Medtr7g101280 | 1947 | 648 | 72.45 | 8.41 | −0.539 |
| MtARF36 | Medtr8g027440 | 2034 | 677 | 75.74 | 6.18 | −0.502 |
| MtARF37 | Medtr8g079492 | 2745 | 914 | 102.26 | 6.08 | −0.605 |
| MtARF38 | Medtr8g100050 | 2502 | 833 | 93.08 | 6.06 | −0.704 |
| MtARF39 | Medtr8g101360 | 3291 | 1096 | 121.07 | 6.23 | −0.522 |
| MtARF40 | Medtr8g446900 | 1146 | 381 | 43.48 | 5.86 | −0.078 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Guo, Z.; Lu, S. Genome-Wide Identification and Expression Analysis of the Aux/IAA and Auxin Response Factor Gene Family in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 10494. https://doi.org/10.3390/ijms221910494
Liu R, Guo Z, Lu S. Genome-Wide Identification and Expression Analysis of the Aux/IAA and Auxin Response Factor Gene Family in Medicago truncatula. International Journal of Molecular Sciences. 2021; 22(19):10494. https://doi.org/10.3390/ijms221910494
Chicago/Turabian StyleLiu, Rui, Zhenfei Guo, and Shaoyun Lu. 2021. "Genome-Wide Identification and Expression Analysis of the Aux/IAA and Auxin Response Factor Gene Family in Medicago truncatula" International Journal of Molecular Sciences 22, no. 19: 10494. https://doi.org/10.3390/ijms221910494