
Genome-Wide Analysis of the TCP Transcription Factor Genes in Dendrobium catenatum Lindl.


Abstract
:1. Introduction
2. Results
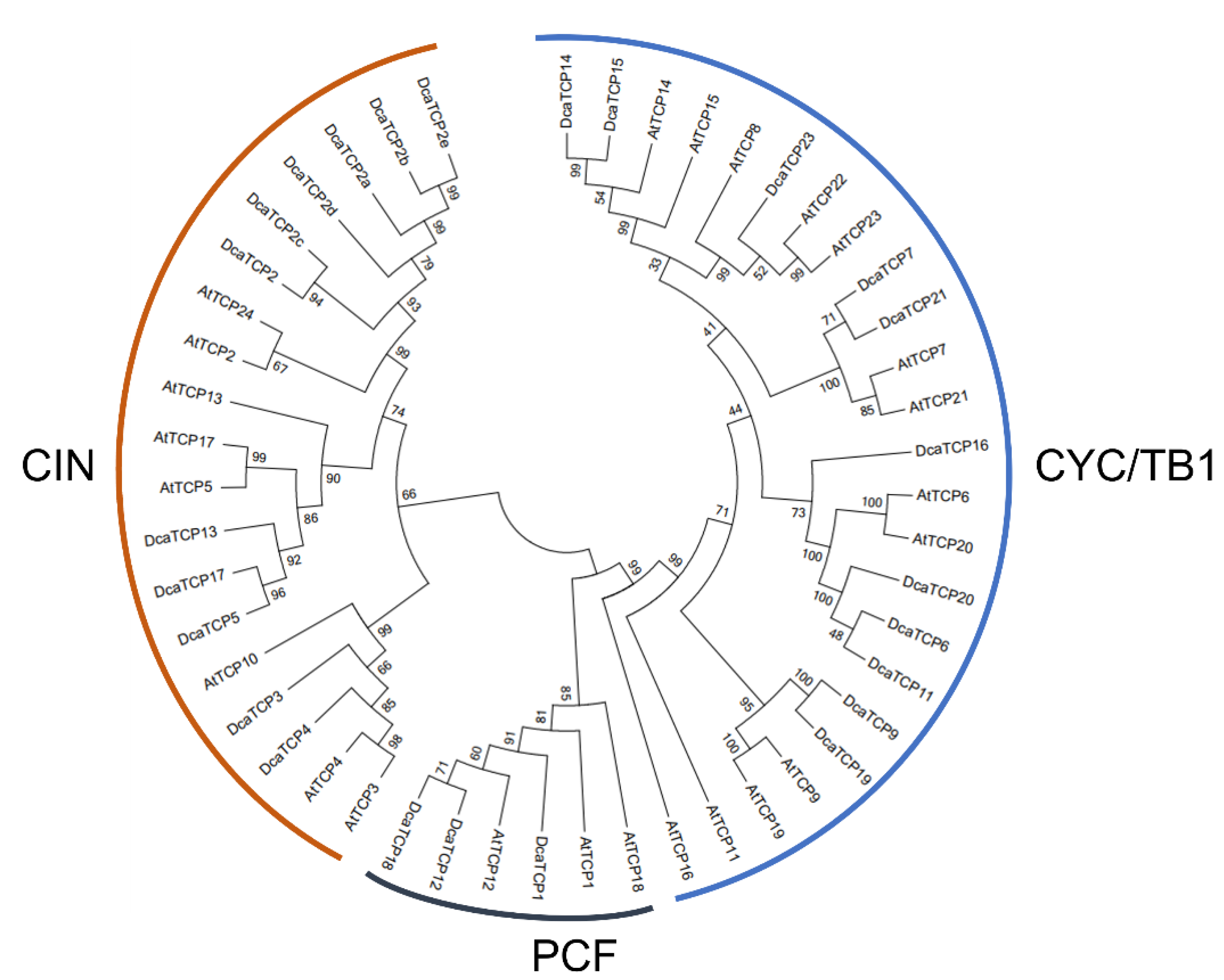
2.1. Identification of TCP Family Genes in D. catenatum
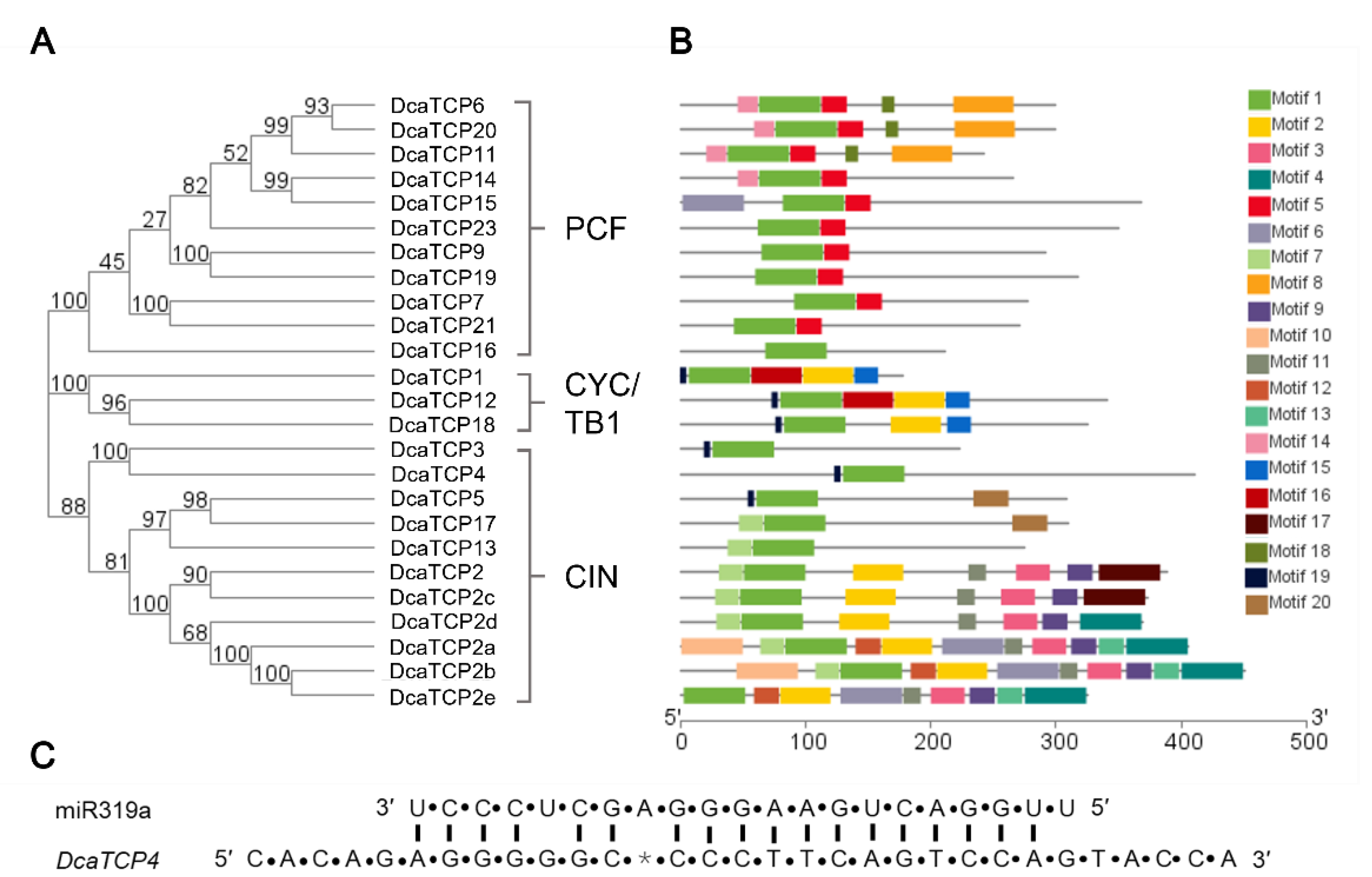
2.2. Conserved Domain and Motif Analysis
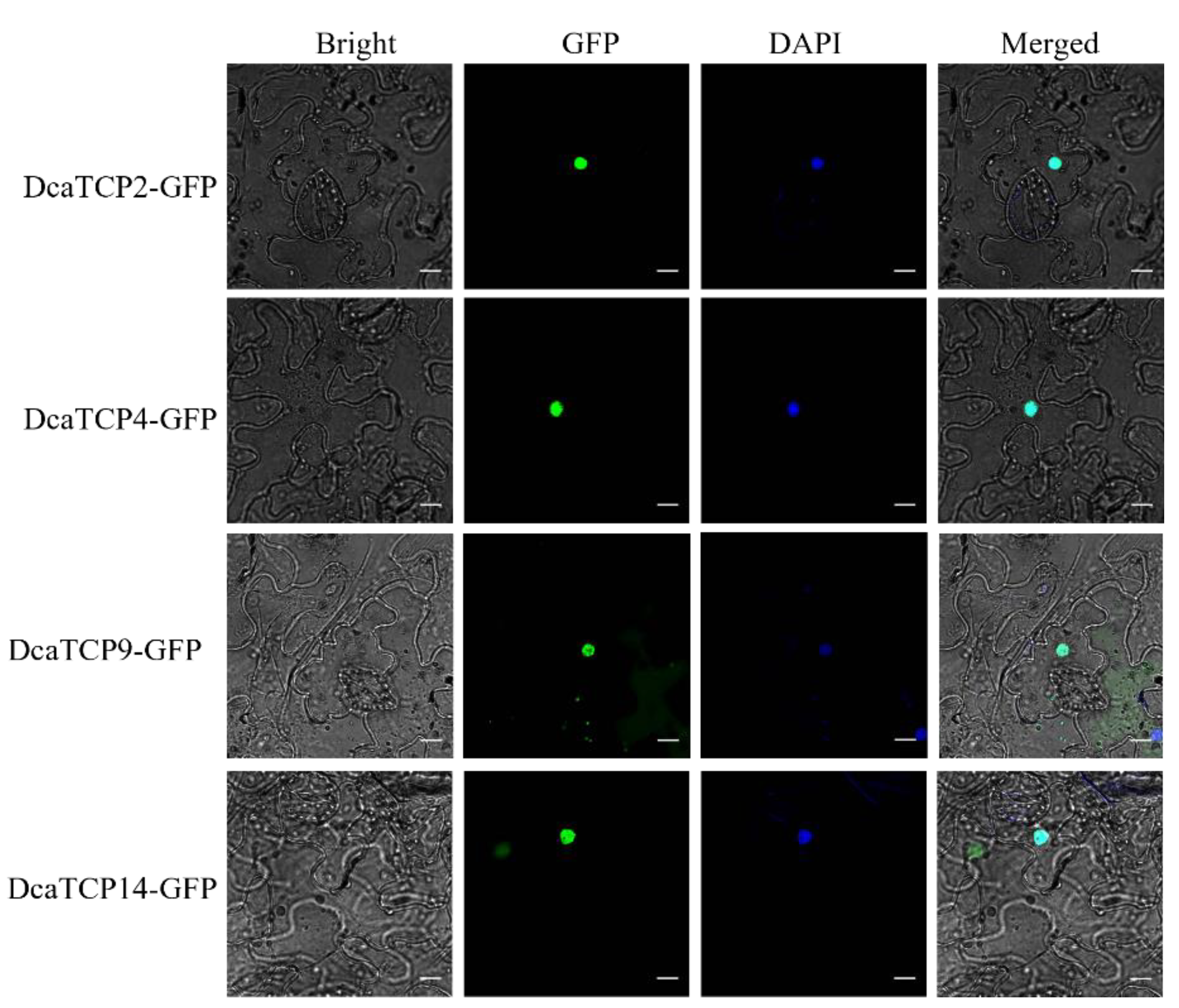
2.3. Subcellular Localization
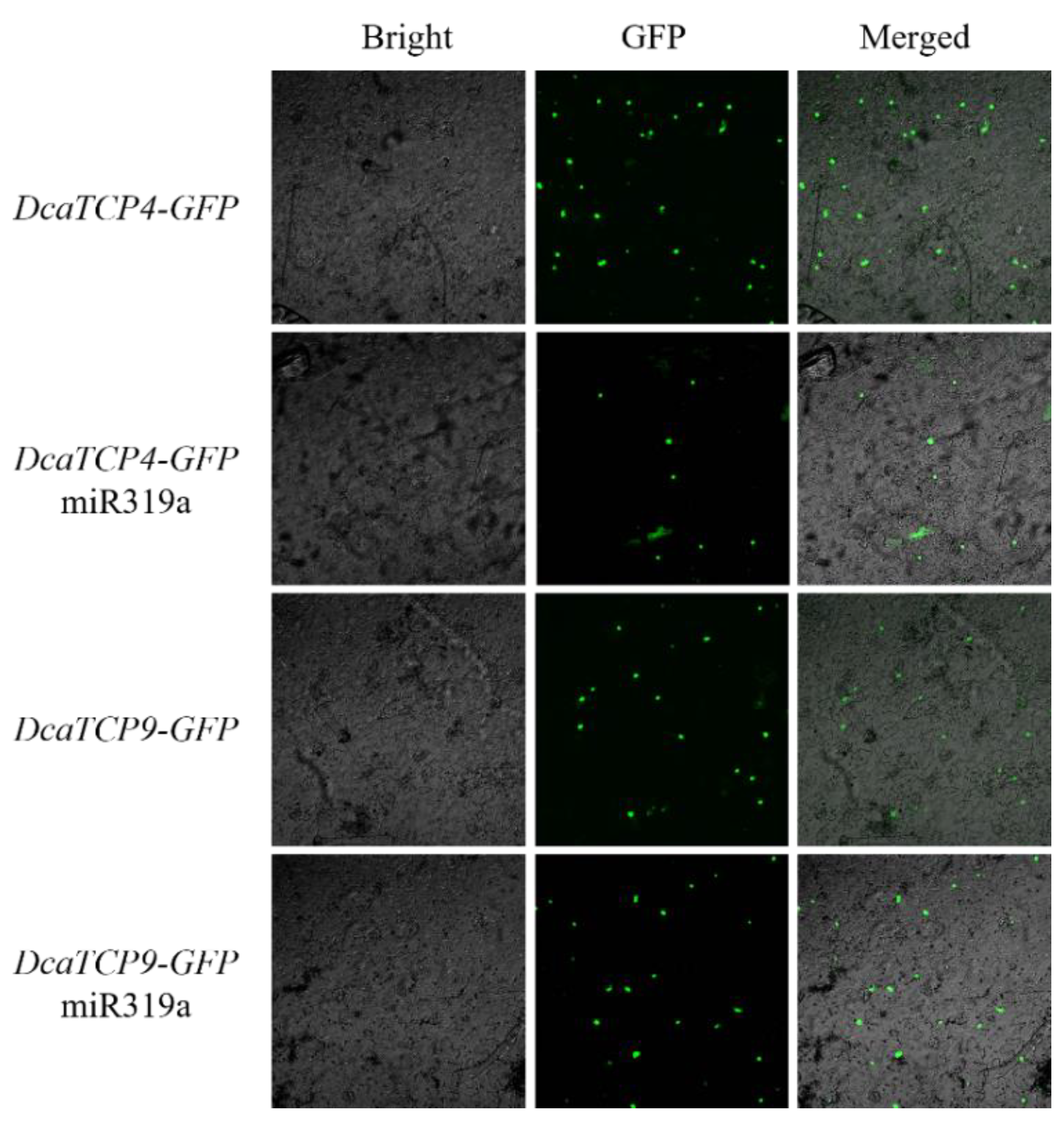
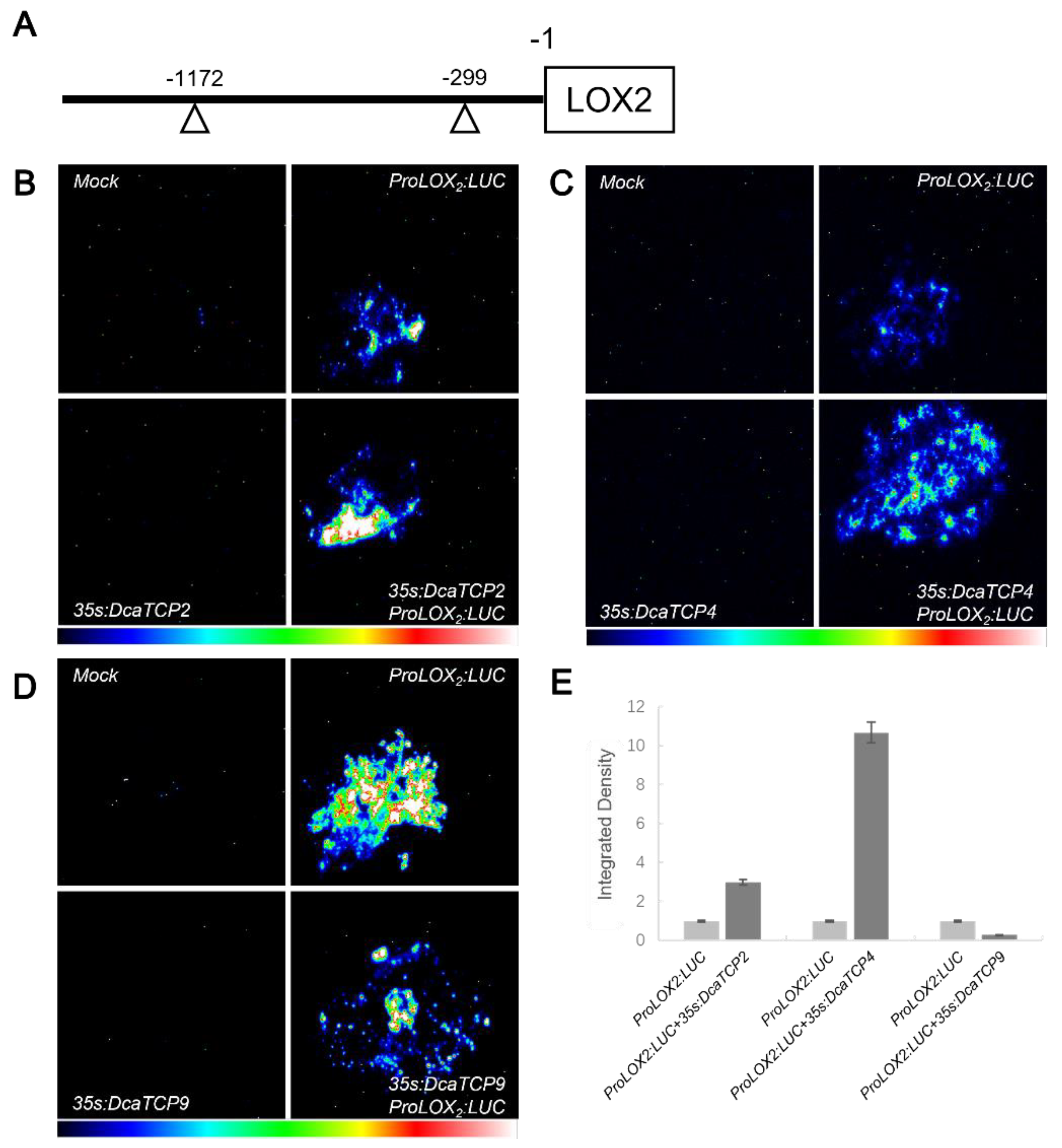
2.4. Class I and II TCPs Antagonistically Regulate LOX2 Expression
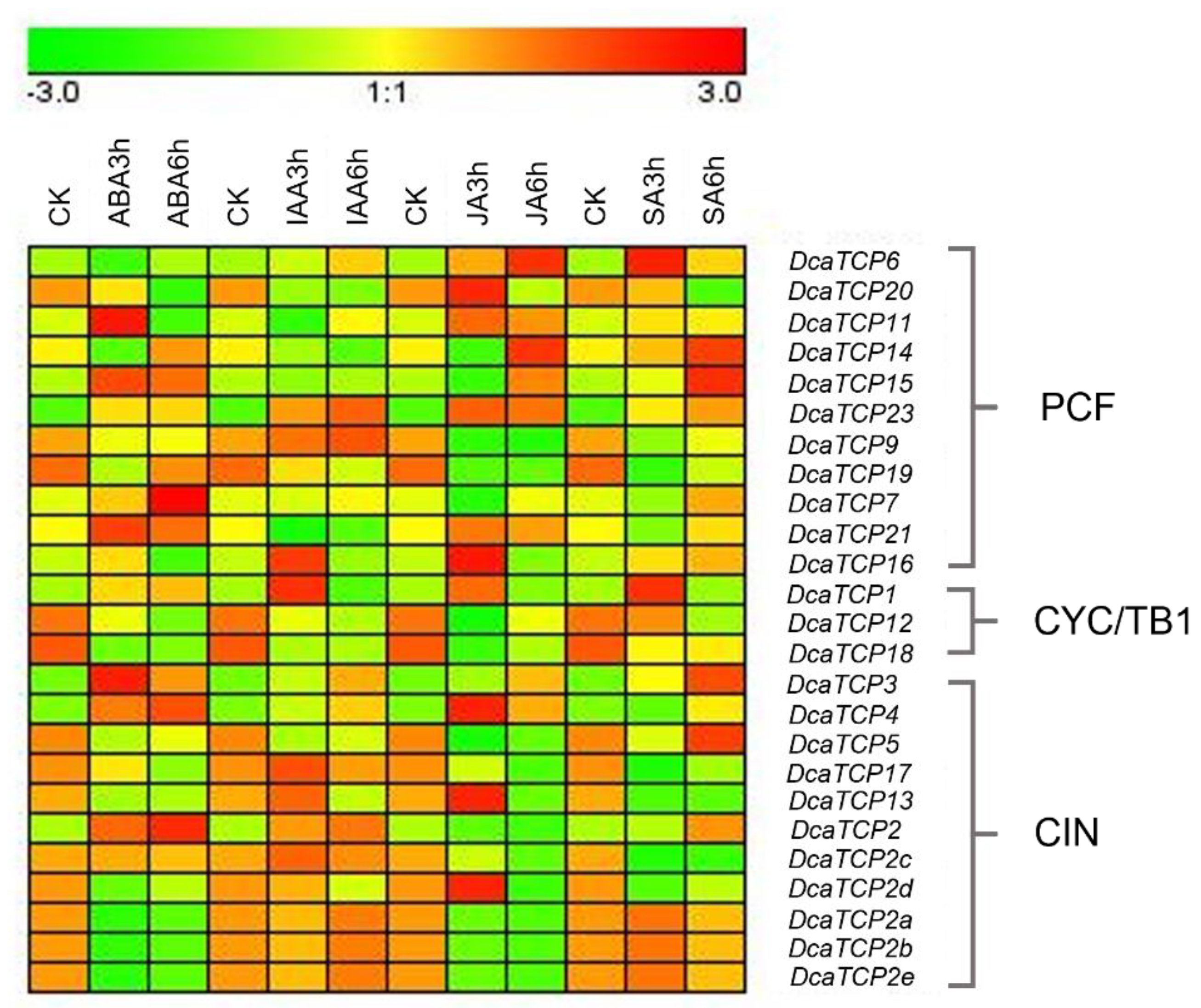
2.5. Expression Profiles of TCP Genes in D. catenatum
3. Discussion
4. Materials and Methods
4.1. Plant Material and Phytohormone Treatments
4.2. Identification of TCP Family Genes in D. catenatum
4.3. Conserved Motif Analysis
4.4. Sequence Characterization and Phylogenetic Analysis
4.5. Subcellular Localization and miRNA-Target Interactions
4.6. Transient Expression in Nicotiana benthamiana
4.7. Transcriptomic Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Luo, D.; Carpenter, R.; Vincent, C.; Copsey, L.; Coen, E. Origin of floral asymmetry in Antirrhinum. Nature 1996, 383, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: A motif found in proteins regulating plant growth and development. Plant J. 1999, 18, 215–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosugi, S.; Ohashi, Y. PCF1 and PCF2 specifically bind to cis elements in the rice proliferating cell nuclear antigen gene. Plant Cell 1997, 9, 1607–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Trillo, M.; Cubas, P. TCP genes: A family snapshot ten years later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Ohashi, Y. DNA binding and dimerization specificity and potential targets for the TCP protein family. Plant J. 2002, 30, 337–348. [Google Scholar] [CrossRef]
- Navaud, O.; Dabos, P.; Carnus, E.; Tremousaygue, D.; Herve, C. TCP transcription factors predate the emergence of land plants. J. Mol. Evol. 2007, 65, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Parapunova, V.; Busscher, M.; Busscher-Lange, J.; Lammers, M.; Karlova, R.; Bovy, A.G.; Angenent, G.C.; de Maagd, R.A. Identification, cloning and characterization of the tomato TCP transcription factor family. BMC Plant Biol. 2014, 14, 157. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Ma, J.; Fan, D.; Li, C.; Jiang, Y.; Luo, K. Genome-wide Identification of TCP Family Transcription Factors from Populus euphratica and Their Involvement in Leaf Shape Regulation. Sci. Rep. 2016, 6, 32795. [Google Scholar] [CrossRef]
- Shi, P.; Guy, K.M.; Wu, W.; Fang, B.; Yang, J.; Zhang, M.; Hu, Z. Genome-wide identification and expression analysis of the ClTCP transcription factors in Citrullus lanatus. BMC Plant Biol. 2016, 16, 85. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xu, Z.; Zhao, K.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. Genome-Wide Identification, Characterization and Expression Analysis of the TCP Gene Family in Prunus mume. Front. Plant Sci. 2016, 7, 1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.T.; Zhou, Q.; Chen, F.; Wu, L.; Liu, B.J.; Li, F.; Zhang, J.Q.; Bao, M.Z.; Liu, G.F. Genome-Wide Identification, Characterization and Expression Analysis of TCP Transcription Factors in Petunia. Int. J. Mol. Sci. 2020, 21, 6594. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Zhang, W.; An, Y.; Du, B.; Wang, D.; Guo, C. Genome-wide analysis of the TCP transcription factor genes in five legume genomes and their response to salt and drought stresses. Funct. Integr. Genom. 2020, 20, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Nag, A.; King, S.; Jack, T. miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 22534–22539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Martínez, J.A.; Poza-Carrión, C.; Cubas, P. Arabidopsis BRANCHED1 Acts as an Integrator of Branching Signals within Axillary Buds. Plant Cell 2007, 19, 458–472. [Google Scholar] [CrossRef]
- Poza-Carrión, C.; Aguilar-Martínez, J.A.; Cubas, P. Role of TCP Gene BRANCHED1 in the Control of Shoot Branching in Arabidopsis. Plant Signal. Behav. 2007, 2, 551–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efroni, I.; Blum, E.; Goldshmidt, A.; Eshed, Y. A protracted and dynamic maturation schedule underlies Arabidopsis leaf development. Plant Cell 2008, 20, 2293–2306. [Google Scholar] [CrossRef] [Green Version]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nat. Genet. 2003, 425, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Mitsuda, N.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. TCP Transcription Factors Regulate the Activities of ASYMMETRIC LEAVES1 and miR164, as Well as the Auxin Response, during Differentiation of Leaves in Arabidopsis. Plant Cell 2010, 22, 3574–3588. [Google Scholar] [CrossRef] [Green Version]
- Schommer, C.; Palatnik, J.F.; Aggarwal, P.; Chetelat, A.; Cubas, P.; Farmer, E.E.; Nath, U.; Weigel, D. Control of jasmonate biosynthesis and senescence by miR319 targets. PLoS Biol. 2008, 6, e230. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Wang, C.; Han, J.; Zhu, X.; Li, X.; Wang, X.; Fang, J. Characterization of miRNAs responsive to exogenous ethylene in grapevine berries at whole genome level. Funct. Integr. Genom. 2017, 17, 213–235. [Google Scholar] [CrossRef] [PubMed]
- Welchen, E.; Gonzalez, D.H. Overrepresentation of elements recognized by TCP-domain transcription factors in the upstream regions of nuclear genes encoding components of the mitochondrial oxidative phosphorylation Machinery. Plant Physiol. 2006, 141, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Qu, Y.; Gao, R.; Jiang, J.; Fang, W.; Guan, Z.; Zhang, F.; Zhao, S.; Chen, S.; Chen, F.; et al. The Heterologous Expression of a Chrysanthemum nankingense TCP Transcription Factor Blocks Cell Division in Yeast and Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 4848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, J.Q.; Qin, G.J. The Regulation of CIN-like TCP Transcription Factors. Int. J. Mol. Sci. 2020, 21, 4498. [Google Scholar] [CrossRef]
- Hou, B.; Luo, J.; Zhang, Y.; Niu, Z.; Xue, Q.; Ding, X. Iteration expansion and regional evolution: Phylogeography of Dendrobium officinale and four related taxa in southern China. Sci. Rep. 2017, 7, 43525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Z.; Zhou, C.; Ji, X.; Wei, G.; Huang, Y.; Yu, W.; Luo, Y.; Qiu, Y. Transcriptome Analysis Reveals genes involved in flavonoid biosynthesis and accumulation in Dendrobium catenatum From Different Locations. Sci. Rep. 2018, 8, 6373. [Google Scholar] [CrossRef]
- Palatnik, J.F.; Wollmann, H.; Schommer, C.; Schwab, R.; Boisbouvier, J.; Rodriguez, R.; Warthmann, N.; Allen, E.; Dezulian, T.; Huson, D.; et al. Sequence and expression differences underlie functional specialization of Arabidopsis microRNAs miR159 and miR319. Dev. Cell 2007, 13, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, F.; Axtell, M.J. Analysis of Complementarity Requirements for Plant MicroRNA Targeting Using a Nicotiana benthamiana Quantitative Transient Assay. Plant Cell 2014, 26, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Danisman, S.; van der Wal, F.; Dhondt, S.; Waites, R.; de Folter, S.; Bimbo, A.; van Dijk, A.-J.; Muino, J.M.; Cutri, L.; Dornelas, M.C.; et al. Arabidopsis Class I and Class II TCP Transcription Factors Regulate Jasmonic Acid Metabolism and Leaf Development Antagonistically. Plant Physiol. 2012, 159, 1511–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.D.; Sun, L.D.; Zhou, Y.Z.; Yang, W.R.; Cheng, T.R.; Wang, J.; Zhang, Q.X. Identification and expression analysis of the SQUAMOSA promoter-binding protein (SBP)-box gene family in Prunus mume. Mol. Genet. Genom. 2015, 290, 1701–1715. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Wang, X.; Liu, H.; Tian, Y.; Lian, J.; Yang, R.; Hao, S.; Wang, X.; Yang, S.; Li, Q.; et al. The Genome of Dendrobium officinale Illuminates the Biology of the Important Traditional Chinese Orchid Herb. Mol. Plant 2015, 8, 922–934. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Zhao, T.; Sheng, Y.; Zheng, T.; Fu, L.; Zhang, Y. Dendrobium officinale Kimura et Migo: A Review on Its Ethnopharmacology, Phytochemistry, Pharmacology, and Industrialization. Evid. Based Complement. Altern. Med. 2017, 2017, 7436259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vick, B.A.; Zimmerman, D.C. The biosynthesis of jasmonic acid—A physiological-role for plant lipoxygenase. Biochem. Biophys. Res. Commun. 1983, 111, 470–477. [Google Scholar] [CrossRef]
- Feng, Z.J.; Xu, S.C.; Liu, N.; Zhang, G.W.; Hu, Q.Z.; Gong, Y.M. Soybean TCP transcription factors: Evolution, classification, protein interaction and stress and hormone responsiveness. Plant Physiol. 2018, 127, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; He, H.; Li, Y.; Yu, D.Q. A new strategy for construction of artificial miRNA vectors in Arabidopsis. Planta 2012, 235, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.Z.; Yan, L.H.; Tan, D.; Chen, R.; Sun, J.Q.; Gao, L.Y.; Dong, M.Q.; Wang, Y.C.; Li, C.Y. Phosphorylation-Coupled Proteolysis of the Transcription Factor MYC2 Is Important for Jasmonate-Signaled Plant Immunity. PLoS Genet. 2013, 9, e1003422. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.P.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.C.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Alex, B.; Jody, C.; Penelope, C.; Eberhardt, R.Y.; Eddy, S.R.; Andreas, H.; Kirstie, H.; Liisa, H.; Jaina, M. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Hu, S.; Yu, Q.; Wang, C.; Yang, Y.; Sun, H.; Yang, Y.; Sun, X. Genome-Wide Identification and Characterization of BrrTCP Transcription Factors in Brassica rapa ssp. rapa. Front. Plant Sci. 2017, 8, 1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zou, Y.; Shang, Y.; Lin, H.; Wang, Y.; Cai, R.; Tang, X.; Zhou, J.-M. Firefly luciferase complementation imaging assay for protein-protein interactions in plants. Plant Physiol. 2008, 146, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | CDS Length (bp) | Protein Size (aa) | MW (kD) | PI | GRAVY |
|---|---|---|---|---|---|---|
| DenTCP1 | XP_028548590 | 537 | 178 | 20.03 | 9.809 | 12.969 |
| DenTCP2 | XP_020673786 | 1170 | 389 | 42.07 | 8.417 | 5.644 |
| DenTCP2a | XP_028551220 | 1221 | 406 | 44.45 | 6.723 | −1.370 |
| DenTCP2b | XP_020679251 | 1353 | 450 | 49.60 | 6.875 | −0.761 |
| DenTCP2c | XP_020703327 | 1122 | 373 | 40.56 | 8.798 | 7.145 |
| DenTCP2d | XP_020675099 | 1110 | 369 | 40.64 | 8.402 | 3.993 |
| DenTCP2e | XP_028551222 | 978 | 325 | 35.19 | 4.808 | −13.867 |
| DenTCP3 | XP_028553615 | 672 | 223 | 24.98 | 9.531 | 6.732 |
| DenTCP4 | XP_020674146 | 1236 | 411 | 45.23 | 9.346 | 9.242 |
| DenTCP5 | XP_028553819 | 927 | 308 | 34.67 | 6.765 | −0.807 |
| DenTCP6 | XP_028550088 | 903 | 300 | 31.82 | 6.740 | −1.591 |
| DenTCP7 | XP_020688226 | 837 | 278 | 29.78 | 10.017 | 9.840 |
| DenTCP9 | XP_020701159 | 879 | 292 | 30.26 | 9.460 | 5.574 |
| DenTCP11 | XP_020699509 | 732 | 243 | 25.64 | 7.132 | 0.397 |
| DenTCP12 | XP_028556330 | 1026 | 341 | 37.73 | 7.472 | 1.493 |
| DenTCP13 | XP_020702002 | 828 | 275 | 30.77 | 6.019 | −3.960 |
| DenTCP14 | XP_020695302 | 801 | 266 | 28.62 | 8.720 | 3.578 |
| DenTCP15 | XP_020697754 | 1107 | 368 | 38.87 | 8.131 | 4.875 |
| DenTCP16 | XP_020686130 | 636 | 211 | 22.32 | 8.154 | 1.389 |
| DenTCP17 | XP_020702494 | 933 | 310 | 35.05 | 7.097 | 0.265 |
| DenTCP18 | XP_028550169 | 978 | 325 | 36.22 | 7.172 | 0.662 |
| DenTCP19 | XP_020673638 | 954 | 317 | 33.20 | 8.933 | 3.916 |
| DenTCP20 | XP_020693581 | 900 | 299 | 31.79 | 8.204 | 2.740 |
| DenTCP21 | XP_020682551 | 816 | 271 | 27.21 | 9.723 | 6.533 |
| DenTCP23 | XP_020696150 | 1053 | 350 | 35.96 | 9.514 | 6.902 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Li, C.; Yang, D.; Wang, Y.; Yang, Y.; Sun, X. Genome-Wide Analysis of the TCP Transcription Factor Genes in Dendrobium catenatum Lindl. Int. J. Mol. Sci. 2021, 22, 10269. https://doi.org/10.3390/ijms221910269
Zhang L, Li C, Yang D, Wang Y, Yang Y, Sun X. Genome-Wide Analysis of the TCP Transcription Factor Genes in Dendrobium catenatum Lindl. International Journal of Molecular Sciences. 2021; 22(19):10269. https://doi.org/10.3390/ijms221910269
Chicago/Turabian StyleZhang, Li, Cheng Li, Danni Yang, Yuhua Wang, Yongping Yang, and Xudong Sun. 2021. "Genome-Wide Analysis of the TCP Transcription Factor Genes in Dendrobium catenatum Lindl." International Journal of Molecular Sciences 22, no. 19: 10269. https://doi.org/10.3390/ijms221910269