
The Jacalin-Related Lectin HvHorcH Is Involved in the Physiological Response of Barley Roots to Salt Stress


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Screening for Salt Stress-Related Genes from Barley cv. Morex Roots in Salt Sensitive Yeast
2.2. In Silico Gene Expression Pattern of HvHorcH in Barley Roots
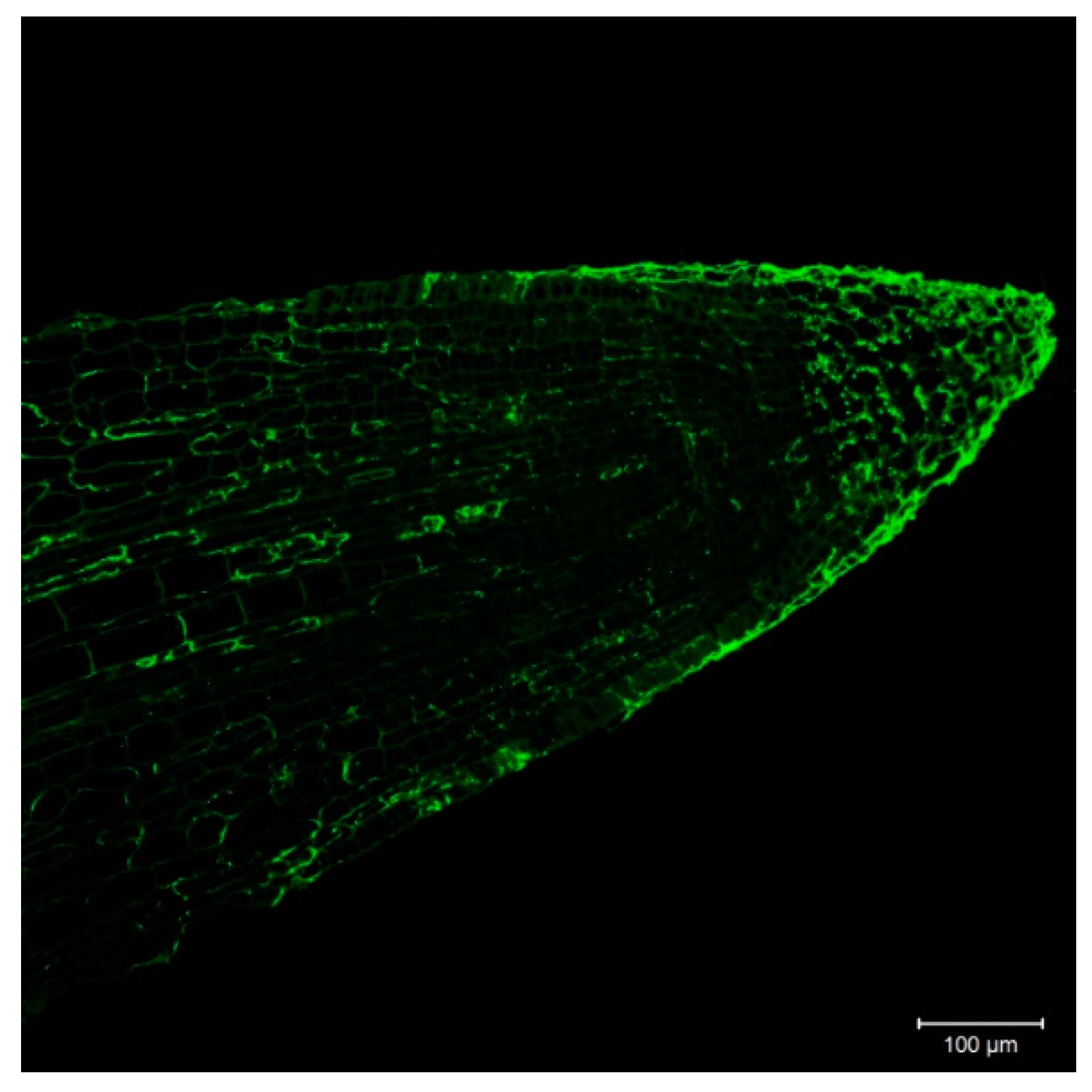
2.3. Cellular and Subcellular Localization of HvHorcH
2.4. Mannose-Binding of HvHorcH Protein
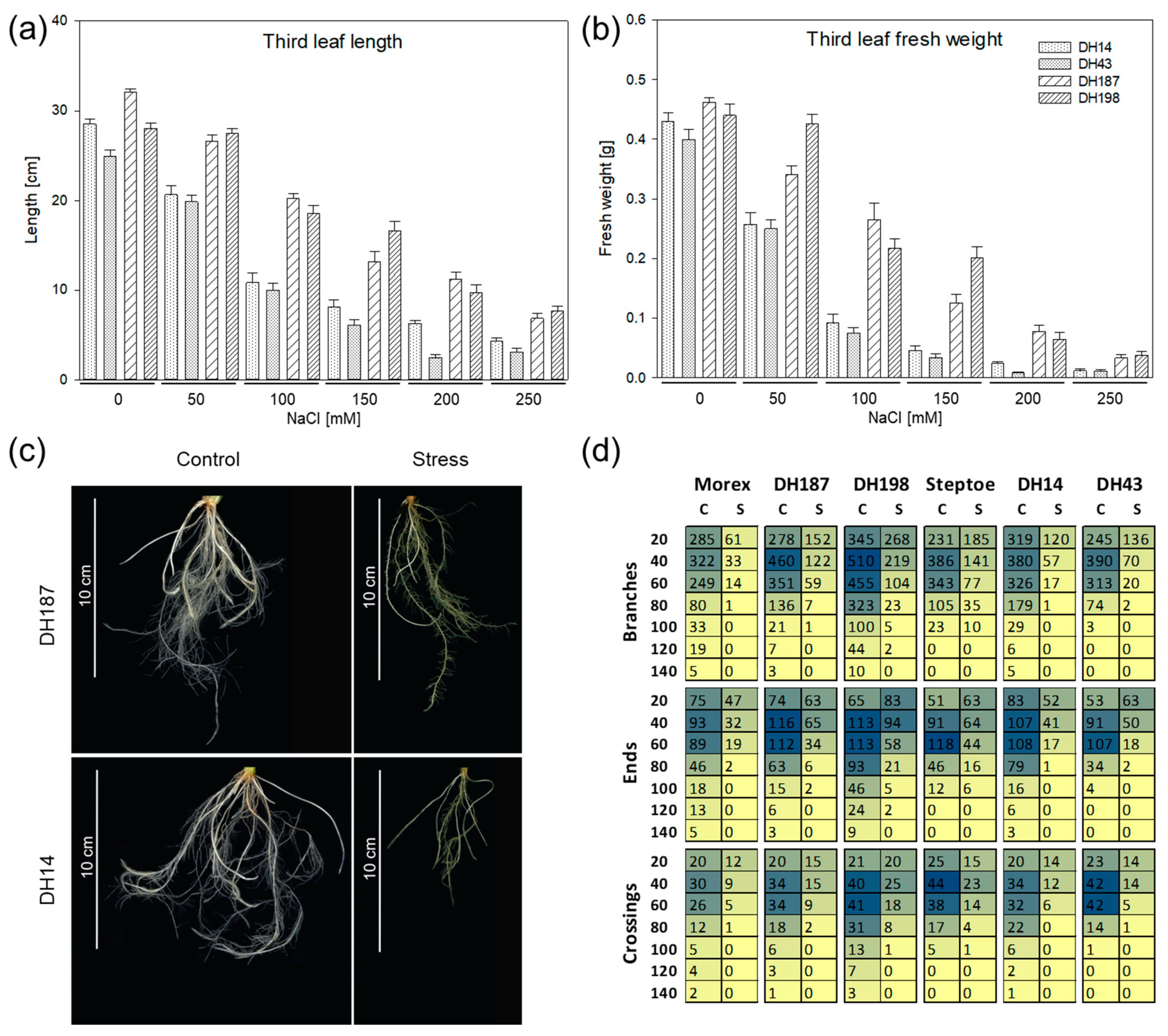
2.5. Selection of Barley Lines with Different Salt Stress Tolerance Levels
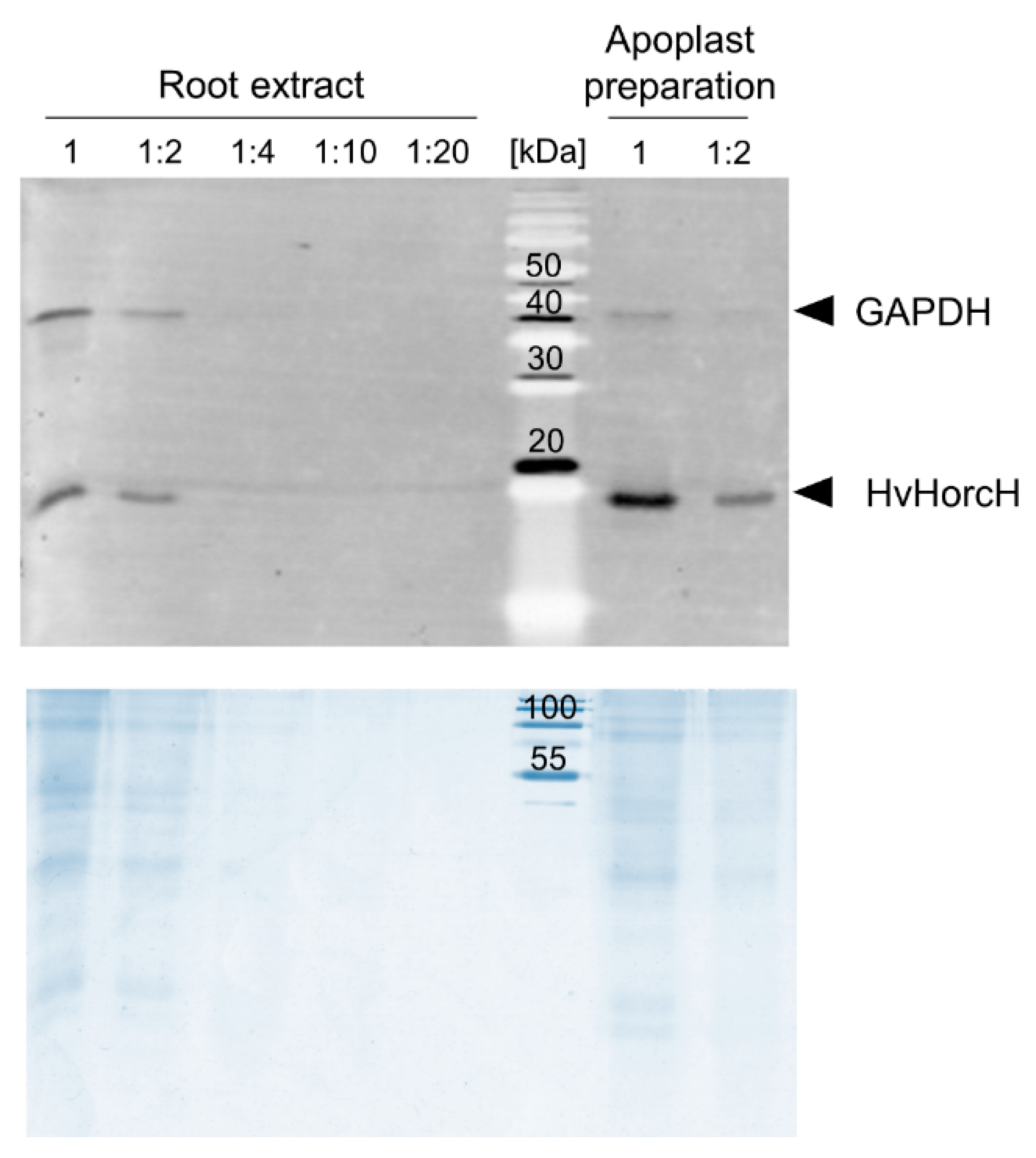
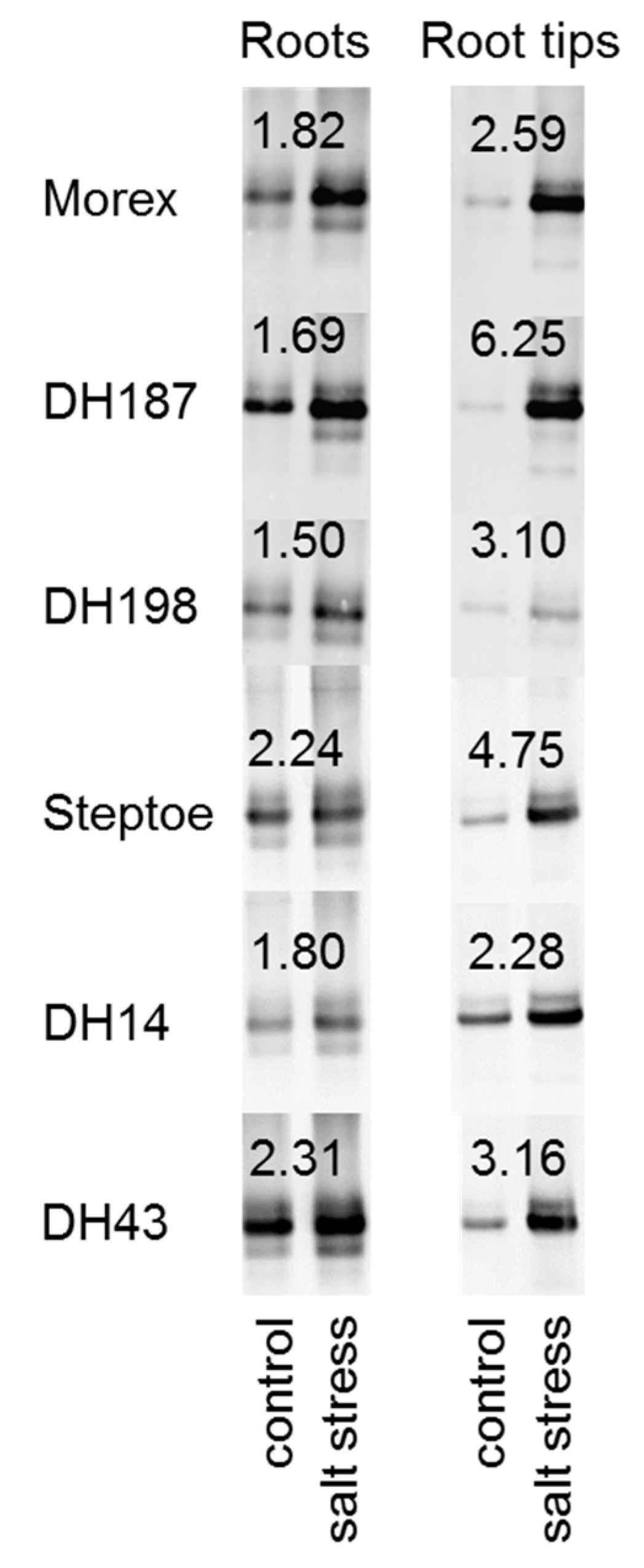
2.6. HvHorcH Protein Abundance Is Highly Increased in Root Tips under Salt Stress
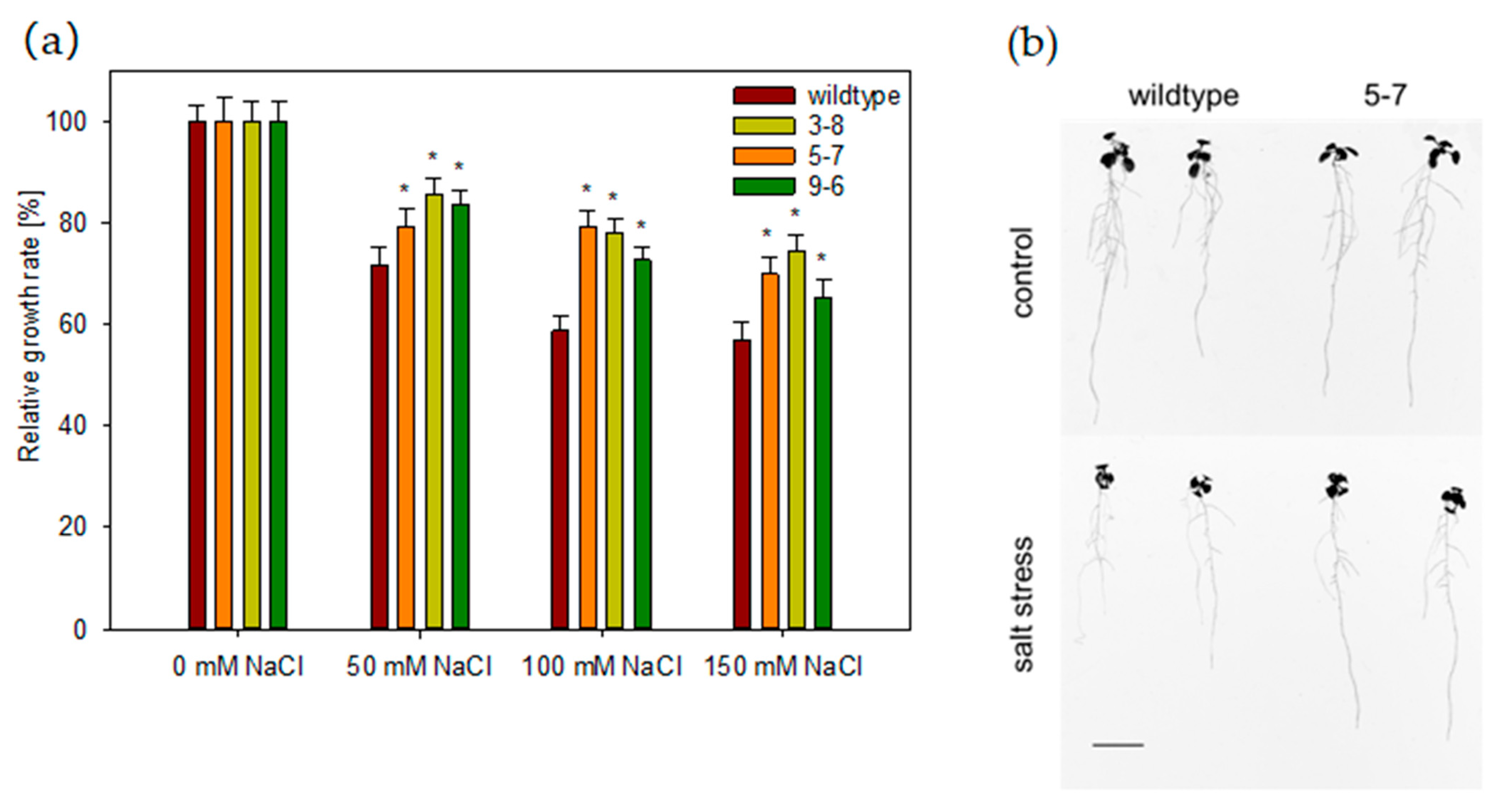
2.7. HvHorcH Enhances Salt Tolerance in Transgenic Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Salt Stress Treatment
4.2. Yeast Strain and Cultivation
4.3. CDNA Library Construction and cDNA Isolation
4.4. Database Search and Sequence Alignments
4.5. Root Morphological Analysis
4.6. Expression of Recombinant HvHorcH in Escherichia coli
4.7. Protein Extraction, SDS-Polyacrylamide Gel Electrophoresis (PAGE), and Gel Blot Analysis
4.8. Microscopy
4.9. Collection of Apoplastic Fluid
4.10. Affinity Enrichment of HvHorcH by Mannose Binding
4.11. Generation of Transgenic Arabidopsis Plants
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butcher, K.; Wick, A.F.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil salinity: A threat to global food security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrao, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Francois, L.E.; Maas, E.V. Crop Response and Management of Salt-Affected Soils. In Handbook of Plant and Crop Stress; Marcel Dekker Press Inc.: New York, NY, USA, 1999; pp. 169–201. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Witzel, K.; Matros, A.; Møller, A.L.B.; Ramireddy, E.; Finnie, C.; Peukert, M.; Rutten, T.; Herzog, A.; Kunze, G.; Melzer, M.; et al. Plasma membrane proteome analysis identifies a role of barley membrane steroid binding protein in root architecture response to salinity. Plant Cell Environ. 2018, 41, 1311–1330. [Google Scholar] [CrossRef]
- Witzel, K.; Matros, A.; Strickert, M.; Kaspar, S.; Peukert, M.; Mühling, K.H.; Börner, A.; Mock, H.-P. Salinity stress in roots of contrasting barley genotypes reveals time-distinct and genotype-specific patterns for defined proteins. Mol. Plant 2014, 7, 336–355. [Google Scholar] [CrossRef] [Green Version]
- Witzel, K.; Weidner, A.; Surabhi, G.-K.; Börner, A.; Mock, H.-P. Salt stress-induced alterations in the root proteome of barley genotypes with contrasting response towards salinity. J. Exp. Bot. 2009, 60, 3545–3557. [Google Scholar] [CrossRef] [Green Version]
- Witzel, K.; Weidner, A.; Surabhi, G.-K.; Varshney, R.K.; Kunze, G.; Buck-Sorlin, G.H.; Börner, A.; Mock, H.-P. Comparative analysis of the grain proteome fraction in barley genotypes with contrasting salinity tolerance during germination. Plant Cell Environ. 2010, 33, 211–222. [Google Scholar] [CrossRef]
- Garciadeblas, B.; Haro, R.; Benito, B. Cloning of two SOS1 transporters from the seagrass Cymodocea nodosa. SOS1 transporters from Cymodocea and Arabidopsis mediate potassium uptake in bacteria. Plant Mol. Biol. 2007, 63, 479–490. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [Green Version]
- Nozawa, A.; Miwa, K.; Kobayashi, M.; Fujiwara, T. Isolation of Arabidopsis thaliana cDNAs that confer yeast boric acid tolerance. Biosci. Biotechnol. Biochem. 2006, 70, 1724–1730. [Google Scholar] [CrossRef]
- Quintero, F.J.; Ohta, M.; Shi, H.Z.; Zhu, J.K.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, R.; Culianz-Macia, F.A.; Moreno, V. Genetic engineering of salt and drought tolerance with yeast regulatory genes. Sci. Hortic. 1999, 78, 261–269. [Google Scholar] [CrossRef]
- Mulet, J.M.; Alemany, B.; Ros, R.; Calvete, J.J.; Serrano, R. Expression of a plant serine O-acetyltransferase in Saccharomyces cerevisiae confers osmotic tolerance and creates an alternative pathway for cysteine biosynthesis. Yeast 2004, 21, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patankar, H.V.; Al-Harrasi, I.; Al-Yahyai, R.; Yaish, M.W. Identification of candidate genes involved in the salt tolerance of date palm (Phoenix dactylifera L.) based on a yeast functional bioassay. DNA Cell Biol. 2018, 37, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Chen, A.-P.; Zhong, N.-Q.; Wang, F.; Wang, H.-Y.; Xia, G.-X. Functional screening of salt stress-related genes from Thellungiella halophila using fission yeast system. Physiol. Plant. 2007, 129, 671–678. [Google Scholar] [CrossRef]
- Batelli, G.; Massarelli, I.; van Oosten, M.; Nurcato, R.; Vannini, C.; Raimondi, G.; Leone, A.; Zhu, J.K.; Maggio, A.; Grillo, S. Asg1 is a stress-inducible gene which increases stomatal resistance in salt stressed potato. J. Plant Physiol. 2012, 169, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Massarelli, I.; Cioffi, R.; Batelli, G.; de Palma, M.; Costa, A.; Grillo, S.; Leone, A. Functional screening of plant stress-related cDNAs by random over-expression in Escherichia coli. Plant Sci. 2006, 170, 880–888. [Google Scholar] [CrossRef]
- Joshi, A.; Dang, H.Q.; Vaid, N.; Tuteja, N. Pea lectin receptor-like kinase promotes high salinity stress tolerance in bacteria and expresses in response to stress in planta. Glycoconj. J. 2010, 27, 133–150. [Google Scholar] [CrossRef]
- Grunwald, I.; Heinig, I.; Thole, H.H.; Neumann, D.; Kahmann, U.; Kloppstech, K.; Gau, A.E. Purification and characterisation of a jacalin-related, coleoptile specific lectin from Hordeum vulgare. Planta 2007, 226, 225–234. [Google Scholar] [CrossRef]
- Pinedo, M.; Orts, F.; de Oliveira Carvalho, A.; Regente, M.; Soares, J.R.; Gomes, V.M.; de la Canal, L. Molecular characterization of Helja, an extracellular jacalin-related protein from Helianthus annuus: Insights into the relationship of this protein with unconventionally secreted lectins. J. Plant Physiol. 2015, 183, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Weidner, A.; Varshney, R.K.; Buck-Sorlin, G.H.; Stein, N.; Graner, A.; Börner, A. QTLs for salt tolerance in three different barley mapping populations. In Proceedings of the 13th International EWAC Conference, Prague, Czech Republic, 27 June–1 July 2005; Börner, A., Pánková, K., Snape, J.W., Eds.; [Google Scholar]
- Bellande, K.; Bono, J.-J.; Savelli, B.; Jamet, E.; Canut, H. Plant lectins and lectin receptor-like kinases: How do they sense the outside? Int. J. Mol. Sci. 2017, 18, 1164. [Google Scholar] [CrossRef] [PubMed]
- Bunn-Moreno, M.M.; Campos-Neto, A. Lectin(s) extracted from seeds of Artocarpus integrifolia (jackfruit): Potent and selective stimulator(s) of distinct human T and B cell functions. J. Immunol. 1981, 127, 427–429. [Google Scholar]
- Esch, L.; Schaffrath, U. An update on jacalin-like lectins and their role in plant defense. Int. J. Mol. Sci. 2017, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Song, M.; Wei, Z.; Tong, J.; Zhang, L.; Xiao, L.; Ma, Z.; Wang, Y. A jacalin-related lectin-like gene in wheat is a component of the plant defence system. J. Exp. Bot. 2011, 62, 5471–5483. [Google Scholar] [CrossRef] [Green Version]
- Al Atalah, B.; Smagghe, G.; van Damme, E.J.M. Orysata, a jacalin-related lectin from rice, could protect plants against biting-chewing and piercing-sucking insects. Plant Sci. 2014, 221–222, 21–28. [Google Scholar] [CrossRef]
- Xiao, J.; Li, C.H.; Xu, S.J.; Xing, L.J.; Xu, Y.Y.; Chong, K. JACALIN-LECTIN LIKE1 Regulates the Nuclear Accumulation of GLYCINE-RICH RNA-BINDING PROTEIN7, Influencing the RNA Processing of FLOWERING LOCUS C Antisense Transcripts and Flowering Time in Arabidopsis. Plant Physiol. 2015, 169, 2102–2117. [Google Scholar]
- Menhaj, A.R.; Mishra, S.K.; Bezhani, S.; Kloppstech, K. Posttranscriptional control in the expression of the genes coding for high-light-regulated HL#2 proteins. Planta 1999, 209, 406–413. [Google Scholar]
- Zhang, W.; Peumans, W.J.; Barre, A.; Astoul, C.H.; Rovira, P.; Rougé, P.; Proost, P.; Truffa-Bachi, P.; Jalali, A.A.; Van Damme, E.J. Isolation and characterization of a jacalin-related mannose-binding lectin from salt-stressed rice (Oryza sativa) plants. Planta 2000, 210, 970–978. [Google Scholar] [PubMed]
- Song, M.; Xu, W.Q.; Xiang, Y.; Jia, H.Y.; Zhang, L.X.; Ma, Z.Q. Association of jacalin-related lectins with wheat responses to stresses revealed by transcriptional profiling. Plant Mol. Biol. 2014, 84, 95–110. [Google Scholar] [CrossRef]
- Abebe, T.; Skadsen, R.W.; Kaeppler, H.F. A proximal upstream sequence controls tissue-specific expression of Lem2, a salicylate-inducible barley lectin-like gene. Planta 2005, 221, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Parthier, B.; Löbler, M. Jasmonate signalling can be uncoupled from abscisic acid signalling in barley: Identification of jasmonate-regulated transcripts which are not induced by abscisic acid. Planta 1996, 199, 625–632. [Google Scholar] [CrossRef]
- Lerner, D.R.; Raikhel, N.V. Cloning and characterization of root-specific barley lectin. Plant Physiol. 1989, 91, 124–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regente, M.; Taveira, G.B.; Pinedo, M.; Elizalde, M.M.; Ticchi, A.J.; Diz, M.S.S.; Carvalho, A.O.; de la Canal, L.; Gomes, V.M. A sunflower lectin with antifungal properties and putative medical mycology applications. Curr. Microbiol. 2014, 69, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Pinedo, M.; Regente, M.; Elizalde, M.; Quiroga, I.Y.; Pagnussat, L.A.; Jorrin-Novo, J.; Maldonado, A.; de la Canal, L. Extracellular sunflower proteins: Evidence on non-classical secretion of a jacalin-related lectin. Protein Pept. Lett. 2012, 19, 270–276. [Google Scholar] [CrossRef]
- Olbrich, A.; Hillmer, S.; Hinz, G.; Oliviusson, P.; Robinson, D.G. Newly formed vacuoles in root meristems of barley and pea seedlings have characteristics of both protein storage and lytic vacuoles. Plant Physiol. 2007, 145, 1383–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cammue, B.; Stinissen, H.M.; Peumans, W.J. Lectin in vegetative tissues of adult barley plants grown under field conditions. Plant Physiol. 1985, 78, 384–387. [Google Scholar] [CrossRef]
- Cammue, B.P.A.; Broekaert, W.F.; Kellens, J.T.C.; Raikhel, N.V.; Peumans, W.J. Stress-induced accumulation of wheat germ agglutinin and abscisic acid in roots of wheat seedlings. Plant Physiol. 1989, 91, 1432–1435. [Google Scholar] [CrossRef]
- Shakirova, F.M.; Bezrukova, M.V.; Aval’baev, A.M.; Fatkhutdinova, R.A. Control mechanisms of lectin accumulation in wheat seedlings under salinity. Russ. J. Plant Physiol. 2003, 50, 301–304. [Google Scholar] [CrossRef]
- Mattos-Moreira, L.A.; Ferreira, C.F.; Amorim, E.P.; Pirovani, C.P.; de Andrade, E.M.; Filho, M.A.C.; da Silva Ledo, C.A. Differentially expressed proteins associated with drought tolerance in bananas (Musa spp.). Acta Physiol. Plant. 2018, 40, 60. [Google Scholar] [CrossRef]
- Claes, B.; Dekeyser, R.; Villarroel, R.; Vandenbulcke, M.; Bauw, G.; Vanmontagu, M.; Caplan, A. Characterization of a rice gene showing organ-specific expression in response to salt stress and drought. Plant Cell 1990, 2, 19–27. [Google Scholar] [PubMed] [Green Version]
- Garcia, A.B.; Engler, J.A.; Claes, B.; Villarroel, R.; van Montagu, M.; Gerats, T.; Caplan, A. The expression of the salt-responsive gene sal T from rice is regulated by hormonal and developmental cues. Planta 1998, 207, 172–180. [Google Scholar] [CrossRef] [PubMed]
- de Souza, G.A.; Ferreira, B.S.; Dias, J.M.; Queiroz, K.S.; Branco, A.T.; Bressan-Smith, R.E.; Oliveira, J.G.; Garcia, A.B. Accumulation of SALT protein in rice plants as a response to environmental stresses. Plant Sci. 2003, 164, 623–628. [Google Scholar]
- Hirano, K.; Teraoka, T.; Yamanaka, H.; Harashima, A.; Kunisaki, A.; Takahashi, H.; Hosokawa, D. Novel mannose-binding rice lectin composed of some isolectins and its relation to a stress-inducible salT gene. Plant Cell Physiol. 2000, 41, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Bezrukova, M.V.; Fatkhutdinova, R.A.; Shakirova, F.M. Protective effect of wheat germ agglutinin on the course of mitosis in the roots of Triticum aestivum seedlings exposed to cadmium. Russ. J. Plant Physiol. 2016, 63, 358–364. [Google Scholar] [CrossRef]
- Bezrukova, M.V.; Lubyanova, A.R.; Fatkhutdinova, R.A. The involvement of wheat and common bean lectins in the control of cell division in the root apical meristems of various plant species. Russ. J. Plant Physiol. 2011, 58, 174–180. [Google Scholar] [CrossRef]
- Rutter, B.D.; Innes, R.W. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017, 173, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-F. Carbohydrate Microarray. In Carbohydrates; Chang, C.-F., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Wilson, S.M.; Burton, R.A.; Collins, H.M.; Doblin, M.S.; Pettolino, F.A.; Shirley, N.; Fincher, G.B.; Bacic, A. Pattern of deposition of cell wall polysaccharides and transcript abundance of related cell wall synthesis genes during differentiation in barley endosperm. Plant Physiol. 2012, 159, 655–670. [Google Scholar] [CrossRef] [Green Version]
- Little, A.; Lahnstein, J.; Jeffery, D.W.; Khor, S.F.; Schwerdt, J.G.; Shirley, N.J.; Hooi, M.; Xing, X.; Burton, R.A.; Bulone, V. A novel (1,4)-β-linked glucoxylan is synthesized by members of the cellulose synthase-like F gene family in land plants. ACS Cent. Sci 2019, 5, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.A.; Jobling, S.A.; Harvey, A.J.; Shirley, N.J.; Mather, D.E.; Bacic, A.; Fincher, G.B. The genetics and transcriptional profiles of the cellulose synthase-like HvCslF gene family in barley. Plant Physiol. 2008, 146, 1821–1833. [Google Scholar] [CrossRef] [Green Version]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Byrt, C.S.; Munns, R.; Burton, R.A.; Gilliham, M.; Wege, S. Root cell wall solutions for crop plants in saline soils. Plant Sci. 2018, 269, 47–55. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legentil, L.; Paris, F.; Ballet, C.; Trouvelot, S.; Daire, X.; Vetvicka, V.; Ferrieres, V. Molecular interactions of β-(1->3)-glucans with their receptors. Molecules 2015, 20, 9745–9766. [Google Scholar] [CrossRef]
- Melida, H.; Sopena-Torres, S.; Bacete, L.; Garrido-Arandia, M.; Jorda, L.; Lopez, G.; Munoz-Barrios, A.; Pacios, L.F.; Molina, A. Non-branched β-1,3-glucan oligosaccharides trigger immune responses in Arabidopsis. Plant J. 2018, 93, 34–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaprakash, N.G.; Singh, A.; Vivek, R.; Yadav, S.; Pathak, S.; Trivedi, J.; Jayaraman, N.; Nandi, D.; Mitra, D.; Surolia, A. The barley lectin, horcolin, binds high-mannose glycans in a multivalent fashion, enabling high-affinity, specific inhibition of cellular HIV infection. J. Biol. Chem. 2020, 295, 12111–12129. [Google Scholar] [CrossRef] [PubMed]
- Albertyn, J.; Hohmann, S.; Thevelein, J.M.; Prior, B.A. Gpd1, which encodes glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic-stress in Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity glycerol response pathway. Mol. Cell. Biol. 1994, 14, 4135–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmsee, C.; Beier, S.; Himmelbach, A.; Schmutzer, T.; Stein, N.; Scholz, U.; Mascher, M. BARLEX–The Barley Draft Genome Explorer. Mol. Plant 2015, 8, 964–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Tommaso, P.; Moretti, S.; Xenarios, I.; Orobitg, M.; Montanyola, A.; Chang, J.M.; Taly, J.F.; Notredame, C. T-Coffee: A web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucleic Acids Res. 2011, 39, W13–W17. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [Green Version]
- Sholl, D.A. Dendritic organization in the neurons of the visual and motor cortices of the cat. J. Anat. 1953, 87, 387–406. [Google Scholar] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotation | Gene | Score (Bits) | E Value | Function | Number of Independent Yeast Transformants |
|---|---|---|---|---|---|
| jacalin-related lectin 31 (HvHorcH) | HORVU7Hr1G059330 | 296 | 7 × 10−103 | carbohydrate binding | 10 |
| cellulose-synthase like D2 | HORVU2Hr1G042350 | 502 | 2 × 10−174 | cellulose biosynthesis | 6 |
| peptide-N(4)-(N-acetyl-β-glucosaminyl)asparagine amidase | HORVU2Hr1G048680 | 507 | 3 × 10−177 | protein deglycosylation | 3 |
| WD-40 repeat family protein | HORVU3Hr1G115170 | 376 | 4 × 10−128 | translation | 3 |
| phytanoyl-CoA dioxygenase domain-containing protein 1 | HORVU4Hr1G007050 | 439 | 3 × 10−156 | fatty acid metabolism | 3 |
| unknown function | HORVU4Hr1G070280 | 64 | 8 × 10−14 | unknown | 3 |
| histone H2A 6 | HORVU7Hr1G100110 | 171 | 1 × 10−52 | DNA binding | 3 |
| 30S ribosomal protein S11 | HORVU7Hr1G104220 | 256 | 8 × 10−88 | translation | 3 |
| exocyst complex component 6 | HORVU0Hr1G006630 | 367 | 2 × 10−122 | vesicle transport | 2 |
| membrane steroid binding protein 1 | HORVU1Hr1G045630 | 276 | 8 × 10−93 | vesicle transport | 2 |
| histone H2A 2 | HORVU2Hr1G043860 | 145 | 1 × 10−44 | DNA binding | 2 |
| unknown function | HORVU7Hr1G032340 | 159 | 2 × 10−48 | unknown | 2 |
| cytochrome P450 superfamily protein | HORVU1Hr1G069310 | 124 | 2 × 10−43 | gibberellin catabolic process | 1 |
| GDSL esterase/lipase | HORVU2Hr1G025800 | 280 | 4 × 10−93 | fatty acid metabolism | 1 |
| adenine nucleotide alpha hydrolases-like superfamily protein | HORVU2Hr1G042480 | 69 | 4 × 10−16 | stress response | 1 |
| ribosomal protein S24e family protein | HORVU4Hr1G058010 | 207 | 1 × 10−67 | unknown | 1 |
| potassium transporter 26 | HORVU4Hr1G058080 | 39 | 7 × 10−04 | ion transport | 1 |
| 26S proteasome non-ATPase regulatory subunit 4 homolog | HORVU4Hr1G063820 | 370 | 3 × 10−126 | proteolysis | 1 |
| ATP-dependent zinc metalloprotease FtsH 2 | HORVU5Hr1G063340 | 68 | 2 × 10−15 | proteolysis | 1 |
| peptidyl-prolyl cis-trans isomerase | HORVU6Hr1G012570 | 295 | 3 × 10−102 | protein folding | 1 |
| Rad23 UV excision repair protein family | HORVU7Hr1G042100 | 221 | 4 × 10−70 | mRNA catabolism | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witzel, K.; Matros, A.; Bertsch, U.; Aftab, T.; Rutten, T.; Ramireddy, E.; Melzer, M.; Kunze, G.; Mock, H.-P. The Jacalin-Related Lectin HvHorcH Is Involved in the Physiological Response of Barley Roots to Salt Stress. Int. J. Mol. Sci. 2021, 22, 10248. https://doi.org/10.3390/ijms221910248
Witzel K, Matros A, Bertsch U, Aftab T, Rutten T, Ramireddy E, Melzer M, Kunze G, Mock H-P. The Jacalin-Related Lectin HvHorcH Is Involved in the Physiological Response of Barley Roots to Salt Stress. International Journal of Molecular Sciences. 2021; 22(19):10248. https://doi.org/10.3390/ijms221910248
Chicago/Turabian StyleWitzel, Katja, Andrea Matros, Uwe Bertsch, Tariq Aftab, Twan Rutten, Eswarayya Ramireddy, Michael Melzer, Gotthard Kunze, and Hans-Peter Mock. 2021. "The Jacalin-Related Lectin HvHorcH Is Involved in the Physiological Response of Barley Roots to Salt Stress" International Journal of Molecular Sciences 22, no. 19: 10248. https://doi.org/10.3390/ijms221910248