
Early Embryo Exposure to Assisted Reproductive Manipulation Induced Subtle Changes in Liver Epigenetics with No Apparent Negative Health Consequences in Rabbit

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Normal Values for Hematological and Serum Metabolites Were Obtained for the F3 Generation Animals Ancestrally Exposed to Embryo Technologies
2.2. Liver Metabolite Profile Changes for the F3 Generation Animals Ancestrally Exposed to Embryo Technologies
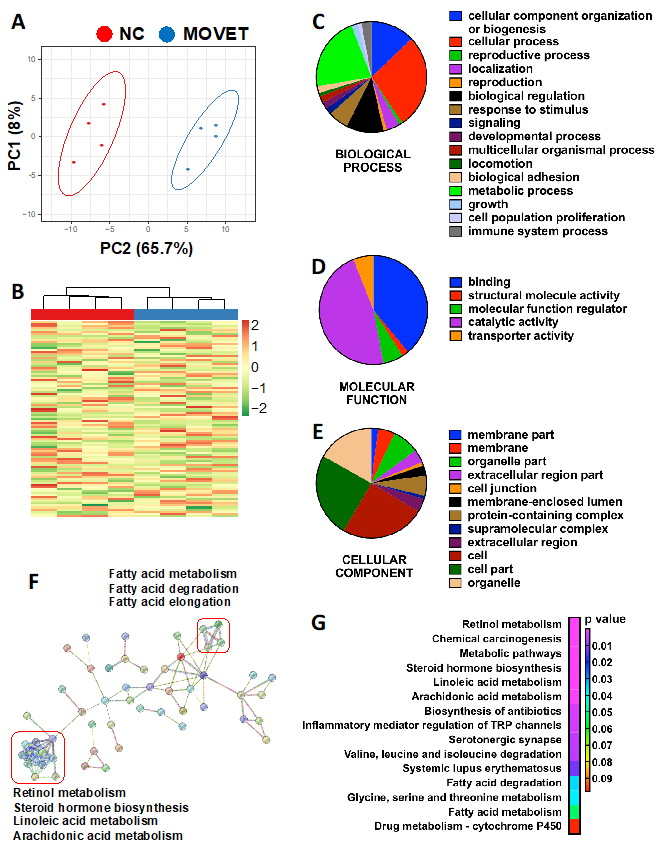
2.3. Liver Protein Profile Changes for the F3 Generation Animals Ancestrally Exposed to Embryo Technologies
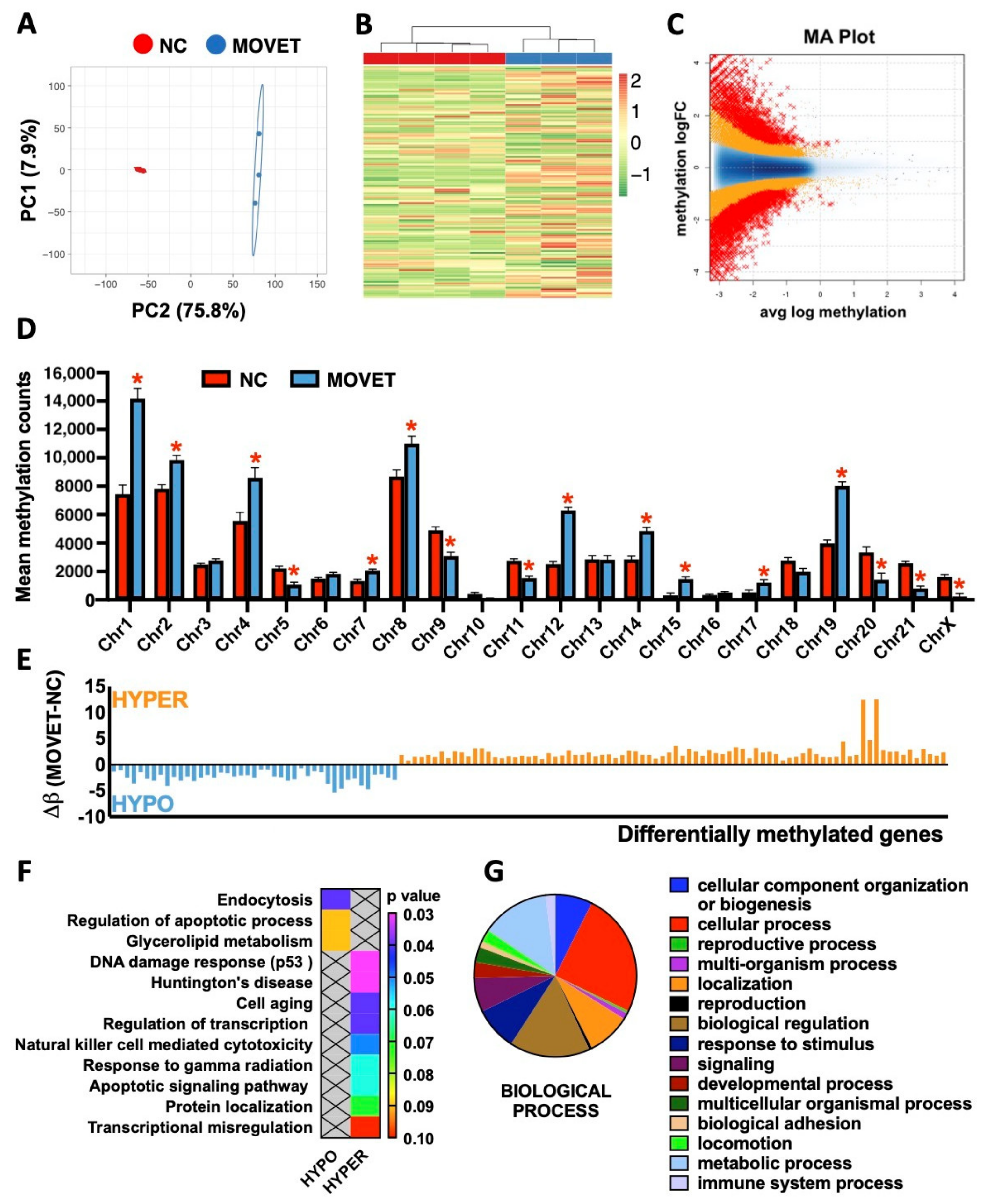
2.4. Genome-Wide Liver Methylation Changes for the F3 Generation Animals Ancestrally Exposed to Embryo Technologies
3. Discussion
4. Materials and Methods
4.1. Animals and Ethical Statements
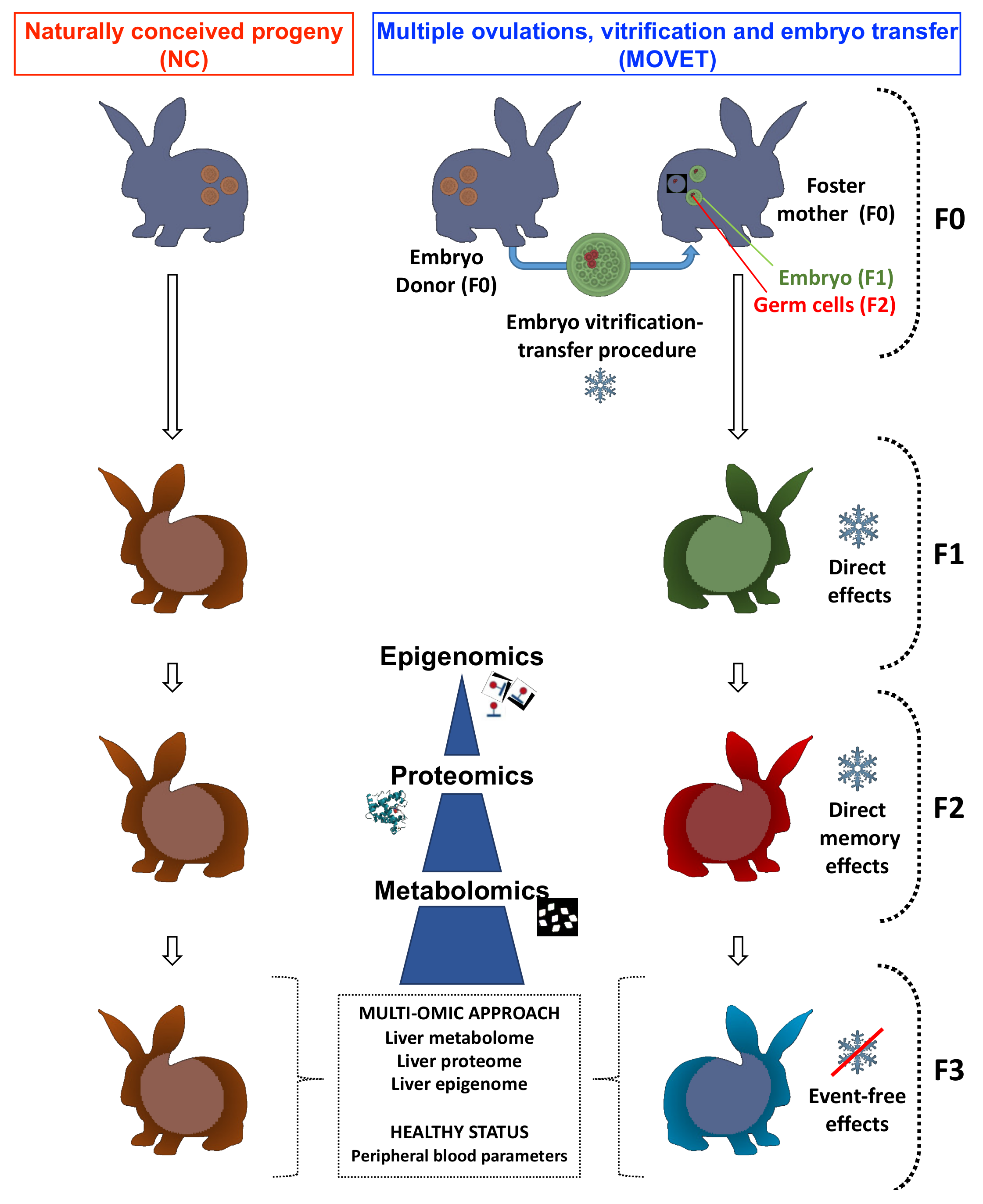
4.2. Experimental Design
4.3. Determination of Hematological and Biochemical Parameters on Peripheral Blood
4.4. Sample Collection for Molecular Study
4.5. Semi-Polar and Non-Polar Metabolome Analysis
4.6. Comparative Proteomic Analysis
4.7. Genome-Wide DNA Methylation Profiling by MBD-Seq
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Biosynthesis of amino acids |
| ANOVA | Analysis of variance |
| ARA | Arachidonic acid metabolism |
| ARTs | Assisted reproductive technologies |
| BUFA | Biosynthesis of unsaturated fatty acids |
| DAMs | Differentially accumulated metabolites |
| DEPs | Differentially expressed proteins |
| DMGs | Differentially methylated genes |
| DNA | Deoxyribonucleic acid |
| DOHaD | Developmental origins of health and disease |
| FDR | False discovery rate |
| GLY | Glycolysis–gluconeogenesis |
| GO | Gene Ontology |
| HM | Heat map |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LC- APCI-HRMS | Liquid chromatography–atmospheric pressure chemical ionization–high resolution mass spectrometry |
| LC-ESI-HRMS | Liquid chromatography–electrospray ionization–high resolution mass spectrometry |
| LCPUFA | Long-chain polyunsaturated fatty acids |
| MOVET | Multiple ovulations, vitrification and embryo transfer |
| MS/MS | Tandem mass spectrometry |
| NC | Naturally conceived |
| PCA | Principal components analysis |
| RNA | Ribonucleic acid |
| SB | Steroid biosynthesis |
| SHB | Steroid hormone biosynthesis |
| STRING | Search Tool for the Retrieval of Interacting Genes/Proteins |
| TAG | Triglycerides |
| TCA | Citric acid cycle |
References
- Monk, D. Germline-derived DNA methylation and early embryo epigenetic reprogramming: The selected survival of imprints. Int. J. Biochem. Cell Biol. 2015, 67, 128–138. [Google Scholar] [CrossRef]
- Ventura-Juncá, P.; Irarrázaval, I.; Rolle, A.J.; Gutiérrez, J.I.; Moreno, R.D.; Santos, M.J. In vitro fertilization (IVF) in mammals: Epigenetic and developmental alterations. Scientific and bioethical implications for IVF in humans. Biol. Res. 2015, 48, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roseboom, T.J. Developmental plasticity and its relevance to assisted human reproduction. Hum. Reprod. 2018, 33, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Vrooman, L.A.; Bartolomei, M.S. Can assisted reproductive technologies cause adult-onset disease? Evidence from human and mouse. Reprod. Toxicol. 2017, 68, 72–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacchini, F.; Sampino, S.; Stankiewicz, A.M.; Haaf, T.; Ptak, G.E. Assessing the epigenetic risks of assisted reproductive technologies: A way forward. Int. J. Dev. Biol. 2019, 63, 217–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narapareddy, L.; Rhon-Calderon, E.A.; Vrooman, L.A.; Baeza, J.; Nguyen, D.K.; Mesaros, C.; Lan, Y.; Garcia, B.A.; Schultz, R.M.; Bartolomei, M.S. Sex-specific effects of in vitro fertilization on adult metabolic outcomes and hepatic transcriptome and proteome in mouse. FASEB J. 2021, 35, e21523. [Google Scholar] [CrossRef]
- Gosden, R.; Trasler, J.; Lucifero, D.; Faddy, M. Rare congenital disorders, imprinted genes, and assisted reproductive technology. Lancet 2003, 361, 1975–1977. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Marjani, S.L.; Jiang, Z. The Epigenetics of Gametes and Early Embryos and Potential Long-Range Consequences in Livestock Species-Filling in the Picture with Epigenomic Analyses. Front. Genet. 2021, 12, 557934. [Google Scholar] [CrossRef]
- Ramos-Ibeas, P.; Heras, S.; Gómez-Redondo, I.; Planells, B.; Fernández-González, R.; Pericuesta, E.; Laguna-Barraza, R.; Pérez-Cerezales, S.; Gutiérrez-Adán, A. Embryo responses to stress induced by assisted reproductive technologies. Mol. Reprod. Dev. 2019, 86, 1292–1306. [Google Scholar] [CrossRef] [Green Version]
- Feuer, S.K.; Rinaudo, P.F. Physiological, metabolic and transcriptional postnatal phenotypes of in vitro fertilization (IVF) in the mouse. J. Dev. Orig. Health Dis. 2017, 8, 403–410. [Google Scholar] [CrossRef]
- Cohen, M.E. The “brave new baby” and the law: Fashioning remedies for the victims of in vitro fertilization. Am. J. Law Med. 1978, 4, 319–336. [Google Scholar] [CrossRef]
- Uk, A.; Collardeau-Frachon, S.; Scanvion, Q.; Michon, L.; Amar, E. Assisted Reproductive Technologies and imprinting disorders: Results of a study from a French congenital malformations registry. Eur. J. Med. Genet. 2018, 61, 518–523. [Google Scholar] [CrossRef]
- Bergh, C.; Wennerholm, U.B. Long-term health of children conceived after assisted reproductive technology. Upsala J. Med. Sci. 2020, 125, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novakovic, B.; Lewis, S.; Halliday, J.; Kennedy, J.; Burgner, D.P.; Czajko, A.; Kim, B.; Sexton-Oates, A.; Juonala, M.; Hammarberg, K.; et al. Assisted reproductive technologies are associated with limited epigenetic variation at birth that largely resolves by adulthood. Nat. Commun. 2019, 10, 3922. [Google Scholar] [CrossRef]
- Duranthon, V.; Chavatte-Palmer, P. Long term effects of ART: What do animals tell us? Mol. Reprod. Dev. 2018, 85, 348–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Rovere, M.; Franzago, M.; Stuppia, L. Epigenetics and neurological disorders in ART. Int. J. Mol. Sci. 2019, 20, 4169. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Zhou, Z.; Zhao, W.; Zou, K.; Shi, W.; Yu, C.; Liu, X.; Dong, Z.; Mao, Y.; Liu, X.; et al. Abnormal Glucose Metabolism in Male Mice Offspring Conceived by in vitro Fertilization and Frozen-Thawed Embryo Transfer. Front. Cell Dev. Biol. 2021, 9, 637781. [Google Scholar] [CrossRef]
- Fiorello, C.V.; Divers, S.J. Rabbits. In Exotic Animal Formulary; Carpenter, J.W., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 546–595. ISBN 978-1-4377-2264-2. [Google Scholar]
- Garcia-Dominguez, X.; Marco-Jiménez, F.; Peñaranda, D.S.; Diretto, G.; García-Carpintero, V.; Cañizares, J.; Vicente, J.S. Long-term and transgenerational phenotypic, transcriptional and metabolic effects in rabbit males born following vitrified embryo transfer. Sci. Rep. 2020, 10, 11313. [Google Scholar] [CrossRef] [PubMed]
- Miles, H.L.; Hofman, P.L.; Peek, J.; Harris, M.; Wilson, D.; Robinson, E.M.; Gluckman, P.D.; Cutfield, W.S. In vitro fertilization improves childhood growth and metabolism. J. Clin. Endocrinol. Metab. 2007, 92, 3441–3445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Zhang, J.; Zheng, M.; Dong, G.; Xu, J.; Zhang, W.; Wu, Y.; Yang, Y.; Zhu, H. A potential high risk for fatty liver disease was found in mice generated after assisted reproductive techniques. J. Cell. Biochem. 2018, 119, 1899–1910. [Google Scholar] [CrossRef]
- Chen, M.; Wu, L.; Zhao, J.; Wu, F.; Davies, M.J.; Wittert, G.A.; Norman, R.J.; Robker, R.L.; Heilbronn, L.K. Altered glucose metabolism in mouse and humans conceived by IVF. Diabetes 2014, 63, 3189–3198. [Google Scholar] [CrossRef] [Green Version]
- Feuer, S.K.; Liu, X.; Donjacour, A.; Lin, W.; Simbulan, R.K.; Giritharan, G.; Piane, L.D.; Kolahi, K.; Ameri, K.; Maltepe, E.; et al. Use of a mouse in vitro fertilization model to understand the developmental origins of health and disease hypothesis. Endocrinology 2014, 155, 1956–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuer, S.K.; Donjacour, A.; Simbulan, R.K.; Lin, W.; Liu, X.; Maltepe, E.; Rinaudo, P.F. Sexually dimorphic effect of In Vitro Fertilization (IVF) on adult mouse fat and liver metabolomes. Endocrinology 2014, 155, 4554–4567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marco-Jiménez, F.; Garcia-dominguez, X.; Domínguez-martínez, M.; Viudes-de-castro, M.P.; Diretto, G.; Peñaranda, D.S.; Vicente, J.S. Effect of embryo vitrification on the steroid biosynthesis of liver tissue in rabbit offspring. Int. J. Mol. Sci. 2020, 21, 8642. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Dominguez, X.; Vicente, J.S.; Marco-Jiménez, F. Developmental Plasticity in Response to Embryo Cryopreservation: The Importance of the Vitrification Device in Rabbits. Animals 2020, 10, 804. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Dominguez, X.; Marco-Jiménez, F.; Peñaranda, D.S.; Vicente, J.S. Long-Term Phenotypic and Proteomic Changes Following Vitrified Embryo Transfer in the Rabbit Model. Animals 2020, 10, 1043. [Google Scholar] [CrossRef]
- Norrman, E.; Petzold, M.; Clausen, T.D.; Henningsen, A.K.; Opdahl, S.; Pinborg, A.; Rosengren, A.; Bergh, C.; Wennerholm, U.B. Type 1 diabetes in children born after assisted reproductive technology: A register-based national cohort study. Hum. Reprod. 2020, 35, 221–231. [Google Scholar] [CrossRef]
- Li, B.; Xiao, X.; Chen, S.; Huang, J.; Ma, Y.; Tang, N.; Sun, H.; Wang, X. Changes of Phospholipids in Fetal Liver of Mice Conceived by In Vitro Fertilization1. Biol. Reprod. 2016, 94, 105. [Google Scholar] [CrossRef]
- Wang, L.Y.; Le, F.; Wang, N.; Li, L.; Liu, X.Z.; Zheng, Y.M.; Lou, H.Y.; Xu, X.R.; Chen, Y.L.; Zhu, X.M.; et al. Alteration of fatty acid metabolism in the liver, adipose tissue, and testis of male mice conceived through assisted reproductive technologies: Fatty acid metabolism in ART mice. Lipids Health Dis. 2013, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.Y.; Liu, X.M.; Jin, L.; Wang, T.T.; Ullah, K.; Sheng, J.Z.; Huang, H.F. Cardiovascular and metabolic profiles of offspring conceived by assisted reproductive technologies: A systematic review and meta-analysis. Fertil. Steril. 2017, 107, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Mintjens, S.; Menting, M.D.; Gemke, R.J.B.J.; van Poppel, M.N.M.; van Wely, M.; Bensdorp, A.J.; Tjon Kon Fat, R.I.; Mol, B.W.J.; Painter, R.C.; van de Beek, C.; et al. The effects of intrauterine insemination and single embryo transfer or modified natural cycle in vitro fertilization on offspring’s health—Follow-up of a randomized clinical trial. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 242, 131–138. [Google Scholar] [CrossRef]
- Lee, J.H. Polyunsaturated fatty acids in children. Pediatr. Gastroenterol. Hepatol. Nutr. 2013, 16, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, J.; Fritsche, K. Linoleic Acid. Adv. Nutr. 2013, 4, 311–312. [Google Scholar] [CrossRef] [PubMed]
- Hadley, K.B.; Ryan, A.S.; Forsyth, S.; Gautier, S.; Salem, N. The essentiality of arachidonic acid in infant development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Czumaj, A.; Śledziński, T. Biological role of unsaturated fatty acid desaturases in health and disease. Nutrients 2020, 12, 356. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.; Jansson, I.; Stoilov, I.; Sarfarazi, M.; Schenkman, J.B. Metabolism of retinoids and arachidonic acid by human and mouse cytochrome P450 1B1. Drug Metab. Dispos. 2004, 32, 840–847. [Google Scholar] [CrossRef] [Green Version]
- Sewer, M.B.; Li, D. Regulation of steroid hormone biosynthesis by the cytoskeleton. Lipids 2008, 43, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Lidén, M.; Eriksson, U. Understanding retinol metabolism: Structure and function of retinol dehydrogenases. J. Biol. Chem. 2006, 281, 13001–13004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Pérez, L.; de Mirecki-Garrido, M.; Guerra, B.; Díaz, M.; Díaz-Chico, J.C. Sex steroids and growth hormone interactions. Endocrinol. Nutr. 2016, 63, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Sudano, M.J.; Rascado, T.D.S.; Tata, A.; Belaz, K.R.A.; Santos, V.G.; Valente, R.S.; Mesquita, F.S.; Ferreira, C.R.; Araújo, J.P.; Eberlin, M.N.; et al. Lipidome signatures in early bovine embryo development. Theriogenology 2016, 86, 472–484.e1. [Google Scholar] [CrossRef] [Green Version]
- Bereshchenko, O.; Bruscoli, S.; Riccardi, C. Glucocorticoids, sex hormones, and immunity. Front. Immunol. 2018, 9, 1332. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S. Role of Vitamin A in the Immune System. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavara, R.; Baselga, M.; Marco-Jiménez, F.; Vicente, J.S. Long-term and transgenerational effects of cryopreservation on rabbit embryos. Theriogenology 2014, 81, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Lavara, R.; Baselga, M.; Marco-Jiménez, F.; Vicente, J.S. Embryo vitrification in rabbits: Consequences for progeny growth. Theriogenology 2015, 84, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Dominguez, X.; Vicente, J.S.; Viudes-De-castro, M.P.; Marco-Jiménez, F. Long-term effects following fresh/vitrified embryo transfer are transmitted by paternal germline in a large size rabbit cohort. Animals 2020, 10, 1272. [Google Scholar] [CrossRef] [PubMed]
- Juarez, J.D.; Marco-Jiménez, F.; Lavara, R.; Vicente, J.S. Rederivation by cryopreservation of a paternal line of rabbits suggests exhaustion of selection for post-weaning daily weight gain after 37 generations. Animals 2020, 10, 1436. [Google Scholar] [CrossRef]
- Marco-Jiménez, F.; Baselga, M.; Vicente, J.S. Successful re-establishment of a rabbit population from embryos vitrified 15 years ago: The importance of biobanks in livestock conservation. PLoS ONE 2018, 13, e0199234. [Google Scholar] [CrossRef] [Green Version]
- Saenz-de-Juano, M.D.; Marco-Jiménez, F.; Vicente, J.S. Embryo transfer manipulation cause gene expression variation in blastocysts that disrupt implantation and offspring rates at birth in rabbit. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 207, 50–55. [Google Scholar] [CrossRef]
- Garcia-Dominguez, X.; Juarez, J.D.; Vicente, J.S.; Marco-Jiménez, F. Impact of embryo technologies on secondary sex ratio in rabbit. Cryobiology 2020, 97, 60–65. [Google Scholar] [CrossRef]
- Cheung, H.-H.; Liu, X.; Rennert, O.M. Apoptosis: Reprogramming and the Fate of Mature Cells. ISRN Cell Biol. 2012, 2012, 685852. [Google Scholar] [CrossRef] [Green Version]
- Mani, S.; Ghosh, J.; Coutifaris, C.; Sapienza, C.; Mainigi, M. Epigenetic changes and assisted reproductive technologies. Epigenetics 2020, 15, 12–25. [Google Scholar] [CrossRef]
- Xu, Q.; Xie, W. Epigenome in Early Mammalian Development: Inheritance, Reprogramming and Establishment. Trends Cell Biol. 2018, 28, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Tang, F. Human Germline Cell Development: From the Perspective of Single-Cell Sequencing. Mol. Cell 2019, 76, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Calle, A.; Fernandez-Gonzalez, R.; Ramos-Ibeas, P.; Laguna-Barraza, R.; Perez-Cerezales, S.; Bermejo-Alvarez, P.; Ramirez, M.A.; Gutierrez-Adan, A. Long-term and transgenerational effects of in vitro culture on mouse embryos. Theriogenology 2012, 77, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Horsthemke, B. A critical view on transgenerational epigenetic inheritance in humans. Nat. Commun. 2018, 9, 2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiken, C.E.; Ozanne, S.E. Transgenerational developmental programming. Hum. Reprod. Update 2014, 20, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Nie, D.; Gui, J.; Zhao, N.; Lin, Y.; Tang, H.; Cai, F.; Shen, G.; Liu, J.; Zhang, E.; Chen, M. Haematological and serum biochemical reference values in Chinese water deer (Hydropotes inermis): A preliminary study. BMC Vet. Res. 2020, 16, 395. [Google Scholar] [CrossRef]
- Estany, J.; Camacho, J.; Baselga, M.; Blasco, A. Selection response of growth rate in rabbits for meat production. Genet. Sel. Evol. 1992, 24, 527–537. [Google Scholar] [CrossRef]
- Garcia-Dominguez, X.; Marco-Jimenez, F.; Viudes-de-Castro, M.P.; Vicente, J.S. Minimally invasive embryo transfer and embryo vitrification at the optimal embryo stage in rabbit model. J. Vis. Exp. 2019, 147, e58055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, B.; Chavatte-Palmer, P.; Viebahn, C.; Navarrete Santos, A.; Duranthon, V. Rabbit as a reproductive model for human health. Reproduction 2012, 144, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K. What is an epigenetic transgenerational phenotype? F3 or F2. Reprod. Toxicol. 2008, 25, 2–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zucker, I.; Beery, A.K. Males still dominate animal studies. Nature 2010, 465, 690. [Google Scholar] [CrossRef]
- Ding, C.; Li, Y.; Guo, F.; Jiang, Y.; Ying, W.; Li, D.; Yang, D.; Xia, X.; Liu, W.; Zhao, Y.; et al. A cell-type-resolved liver proteome. Mol. Cell. Proteomics 2016, 15, 3190–3202. [Google Scholar] [CrossRef] [Green Version]
- Diretto, G.; Rubio-Moraga, A.; Argando, A.J.; Castillo, P.; Gómez-Gómez, L.; Ahrazem, O. Tissue-specific accumulation of sulfur compounds and saponins in different parts of garlic cloves from purple and white ecotypes. Molecules 2017, 22, 1359. [Google Scholar] [CrossRef]
- Cappelli, G.; Giovannini, D.; Basso, A.L.; Demurtas, O.C.; Diretto, G.; Santi, C.; Girelli, G.; Bacchetta, L.; Mariani, F. A Corylus avellana L. extract enhances human macrophage bactericidal response against Staphylococcus aureus by increasing the expression of anti-inflammatory and iron metabolism genes. J. Funct. Foods 2018, 45, 499–511. [Google Scholar] [CrossRef]
- Di Meo, F.; Aversano, R.; Diretto, G.; Demurtas, O.C.; Villano, C.; Cozzolino, S.; Filosa, S.; Carputo, D.; Crispi, S. Anti-cancer activity of grape seed semi-polar extracts in human mesothelioma cell lines. J. Funct. Foods 2019, 61, 103515. [Google Scholar] [CrossRef]
- Fiore, A.; Dall’Osto, L.; Cazzaniga, S.; Diretto, G.; Giuliano, G.; Bassi, R. A quadruple mutant of Arabidopsis reveals a β-carotene hydroxylation activity for LUT1/CYP97C1 and a regulatory role of xanthophylls on determination of the PSI/PSII ratio. BMC Plant Biol. 2012, 12, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambla, J.L.; Trapero-Mozos, A.; Diretto, G.; Moraga, A.R.; Granell, A.; Gómez, L.G.; Ahrazem, O. Gene-metabolite networks of volatile metabolism in Airen and Tempranillo grape cultivars revealed a distinct mechanism of aroma bouquet production. Front. Plant Sci. 2016, 7, 1619. [Google Scholar] [CrossRef] [Green Version]
- Sulli, M.; Mandolino, G.; Sturaro, M.; Onofri, C.; Diretto, G.; Parisi, B.; Giuliano, G. Molecular and biochemical characterization of a potato collection with contrasting tuber carotenoid content. PLoS ONE 2017, 12, e0184143. [Google Scholar] [CrossRef]
- Shilov, I.V.; Seymourt, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The paragon algorithm, a next generation search engine that uses sequence temperature values sequence temperature values and feature probabilities to identify peptides from tandem mass spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Peña-Llopis, S.; Brugarolas, J. Simultaneous isolation of high-quality DNA, RNA, miRNA and proteins from tissues for genomic applications. Nat. Protoc. 2013, 8, 2240–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lienhard, M.; Grimm, C.; Morkel, M.; Herwig, R.; Chavez, L. MEDIPS: Genome-wide differential coverage analysis of sequencing data derived from DNA enrichment experiments. Bioinformatics 2014, 30, 284–286. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Parameters | NC | MOVET |
|---|---|---|
| Hematology | ||
| White blood cells (103/mm3) | 9.1 ± 1.71 | 7.4 ± 1.71 |
| Lymphocytes (%) | 40.1 ± 2.75 | 36.0 ± 2.75 |
| Monocytes (%) | 7.3 ± 0.72 | 6.4 ± 0.72 |
| Granulocytes (%) | 46.7 ± 2.83 | 49.3 ± 2.83 |
| Red blood cells (106/mm3) | 6.1 ± 0.27 | 6.3 ± 0.27 |
| Hemoglobin (g/dL) | 12.6 ± 0.63 | 13.1 ± 0.63 |
| Hematocrit (%) | 43.8 ± 2.26 | 44.0 ± 2.26 |
| Serum metabolites + | ||
| Albumin (g/dL) | 4.4 ± 0.91 | 4.5 ± 0.91 |
| Bile acids (µmol/L) | 6.9 ± 1.08 | 7.1 ± 1.08 |
| Cholesterol (mg/dL) | 32.2 ± 1.94 a | 25.6 ± 1.94 b |
| Glucose (mg/dL) | 103.3 ± 10.39 b | 138.7 ± 10.39 a |
| Bilirubin total (mg/dL) | 0.1 ± 0.02 | 0.1 ± 0.02 |
| Number of animals | 10 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Domínguez, X.; Diretto, G.; Peñaranda, D.S.; Frusciante, S.; García-Carpintero, V.; Cañizares, J.; Vicente, J.S.; Marco-Jiménez, F. Early Embryo Exposure to Assisted Reproductive Manipulation Induced Subtle Changes in Liver Epigenetics with No Apparent Negative Health Consequences in Rabbit. Int. J. Mol. Sci. 2021, 22, 9716. https://doi.org/10.3390/ijms22189716
García-Domínguez X, Diretto G, Peñaranda DS, Frusciante S, García-Carpintero V, Cañizares J, Vicente JS, Marco-Jiménez F. Early Embryo Exposure to Assisted Reproductive Manipulation Induced Subtle Changes in Liver Epigenetics with No Apparent Negative Health Consequences in Rabbit. International Journal of Molecular Sciences. 2021; 22(18):9716. https://doi.org/10.3390/ijms22189716
Chicago/Turabian StyleGarcía-Domínguez, Ximo, Gianfranco Diretto, David S. Peñaranda, Sarah Frusciante, Victor García-Carpintero, Joaquín Cañizares, José S. Vicente, and Francisco Marco-Jiménez. 2021. "Early Embryo Exposure to Assisted Reproductive Manipulation Induced Subtle Changes in Liver Epigenetics with No Apparent Negative Health Consequences in Rabbit" International Journal of Molecular Sciences 22, no. 18: 9716. https://doi.org/10.3390/ijms22189716