
Regulation of Ribosomal Protein Synthesis in Mycobacteria: The Autogenous Control of rpsO

Abstract
:1. Introduction
2. Results and Discussion
2.1. A Strategy for Using Escherichia coli as a Host for Studying the Autogenous Regulation of Mycobacterial r-Proteins
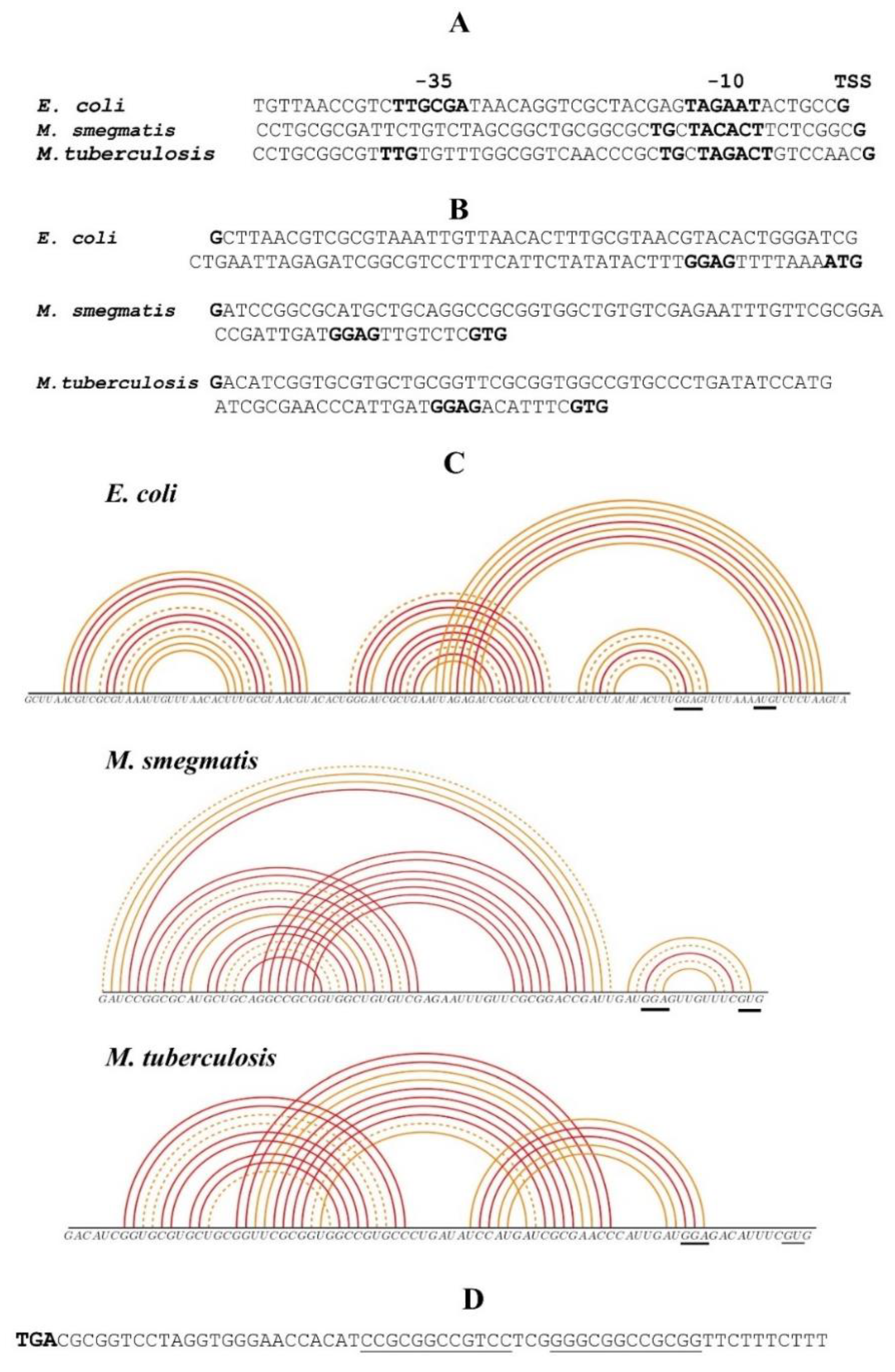
2.2. Comparison of Regulatory Regions of Mycobacterial and E. coli rpsO Genes
2.3. The rpsO Promoter from M. smegmatis Is Inoperative in E. coli
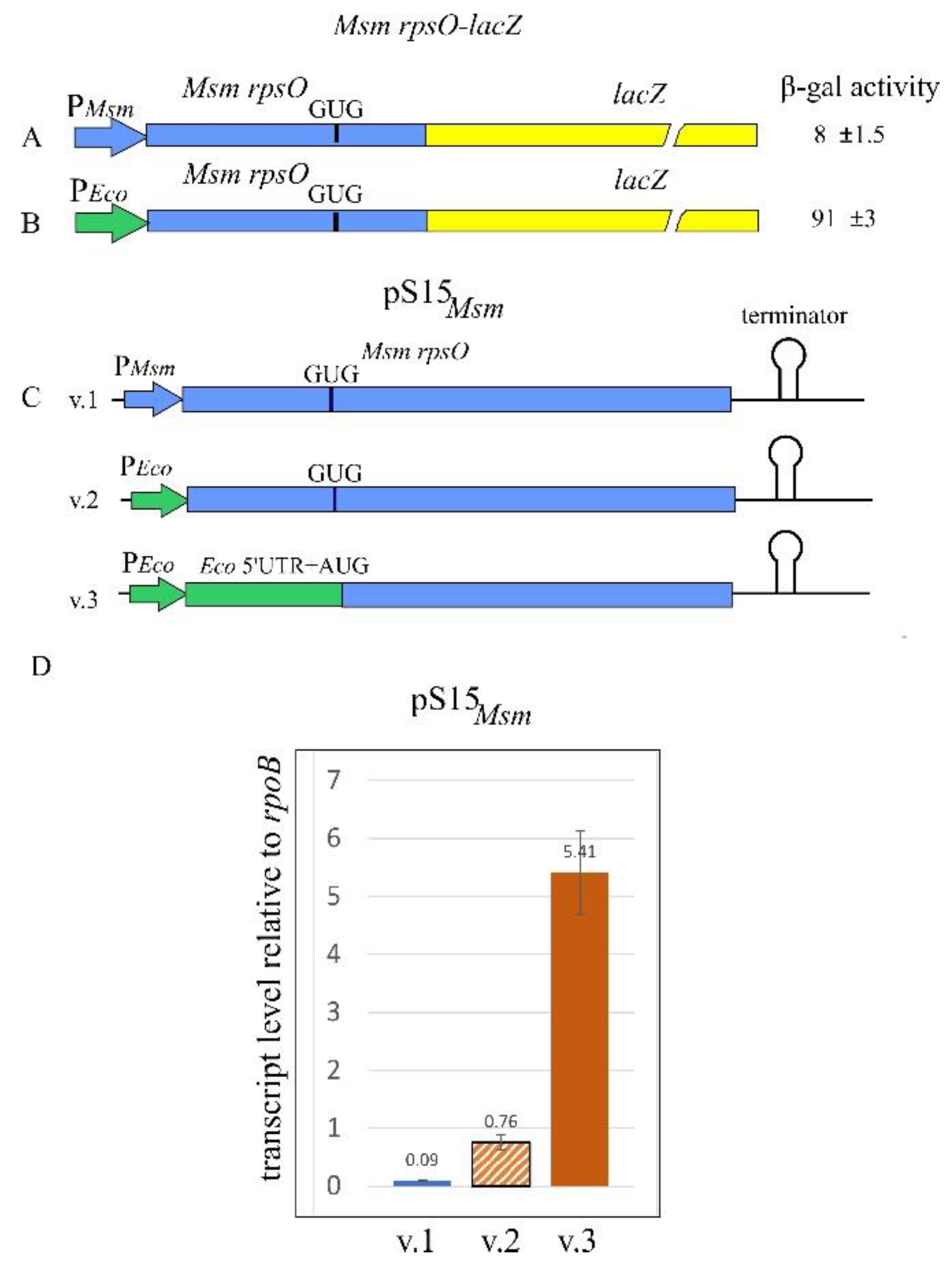
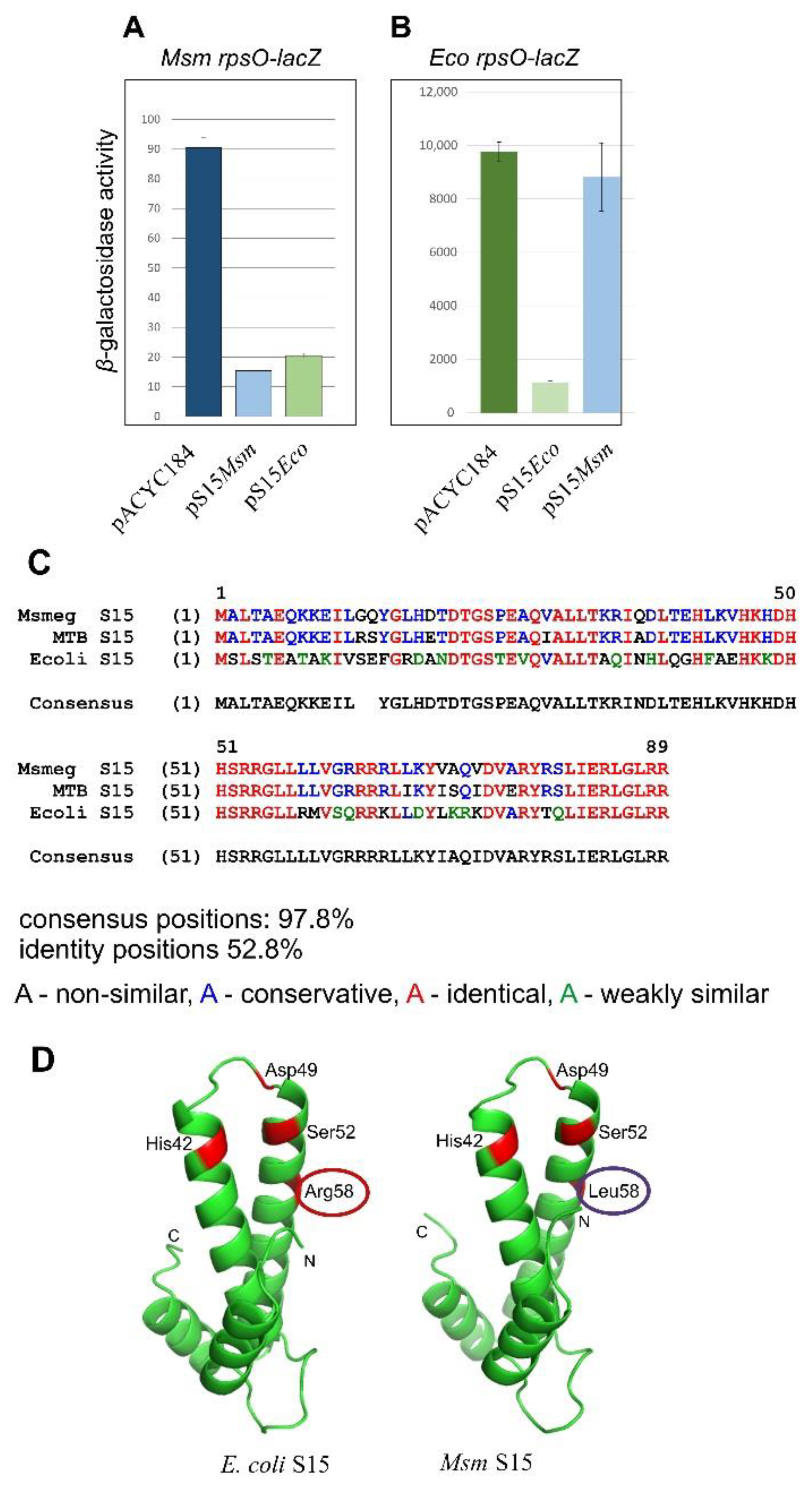
2.4. The Msm rpsO’-‘lacZ Reporter Is Regulated by Both Msm and E. coli S15
2.5. Creation of the Cognate System for Studying the Autogenous Control of r-Protein Synthesis in Mycobacteria
2.6. The Msm rpsO Core Promoter Requires an Upstream Region to Enhance the Transcription Yield
2.7. Generation of the Novel Replicative Shuttle Vector, pAMYC
2.8. Mycobacterial rpsO Expression Is Feedback-Regulated at the Translation Level
2.9. The Pseudoknot in the Msm rpsO 5′UTR Is Essential for the Autogenous Control
2.10. Concluding Remarks
3. Materials and Methods
3.1. Strains and Plasmids
3.2. Construction of Expression Plasmids for Use in E. coli as a Surrogate Host
3.3. Quantification of the In Vivo Transcripts by RT-qPCR with an Internal Standard
3.4. Construction of the Msm_rpsO’-‘lacZ Fusions Integrated into the E. coli Chromosome
3.5. Cell Growth and β-Galactosidase Assay
3.6. Creation of the Novel Escherichia coli-Mycobacteria Shuttle Vector pAMYC, a Derivative of pACYC184
3.7. Modification of the Integrative Plasmid pMV306hsp to Provide Insertion of the rpsO-egfp Reporter Construct into the Chromosome of M. smegmatis
3.8. Mutagenesis of the Msm rpsO 5′UTR to Prevent Pseudoknot Formation
3.9. Creating Plasmids for Ectopic Expression of the Msm (or E. coli) rpsO Gene in M. smegmatis
3.10. Cell Growth and eGFP Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nomura, M.; Gourse, R.; Baughman, G. Regulation of the synthesis of ribosomes and ribosomal components. Ann. Rev. Biochem. 1984, 53, 75–117. [Google Scholar] [CrossRef]
- Zengel, J.M.; Lindahl, L. Diverse mechanisms for regulating ribosomal protein synthesis in Escherichia coli. Prog. Nucleic Acid Res. Mol. Biol. 1994, 47, 331–370. [Google Scholar] [CrossRef]
- Fu, Y.; Deiorio-Haggar, K.; Anthony, J.; Meyer, M.M. Most RNAs regulating ribosomal protein biosynthesis in Escherichia coli are narrowly distributed to Gammaproteobacteria. Nucleic Acids Res. 2013, 41, 3491–3503. [Google Scholar] [CrossRef] [Green Version]
- Babina, A.M.; Parker, D.J.; Li, G.W.; Meyer, M.M. Fitness advantages conferred by the L20-interacting RNA cis-regulator of ribosomal protein synthesis in Bacillus subtilis. RNA 2018, 24, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Babina, A.M.; Soo, M.W.; Fu, Y.; Meyer, M.M. An S6:S18 complex inhibits translation of E. coli rpsF. RNA 2015, 21, 2039–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aseev, L.V.; Bylinkina, N.S.; Boni, I.V. Regulation of the rplY gene encoding 5S rRNA binding protein L25 in Escherichia coli and related bacteria. RNA 2015, 21, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Aseev, L.V.; Koledinskaya, L.S.; Boni, I.V. Regulation of ribosomal protein operons rplM-rpsI, rpmB-rpmG, and rplU-rpmA at the transcriptional and translational levels. J. Bacteriol. 2016, 198, 2494–2502. [Google Scholar] [CrossRef] [Green Version]
- Aseev, L.V.; Koledinskaya, L.S.; Boni, I.V. Autogenous regulation in vivo of the rpmE gene encoding ribosomal protein L31 (bL31), a key component of the protein-protein intersubunit bridge B1b. RNA 2020, 26, 814–826. [Google Scholar] [CrossRef]
- Philippe, C.; Eyermann, F.; Benard, L.; Portier, C.; Ehresmann, B.; Ehresmann, C. Ribosomal protein S15 from Escherichia coli modulates its own translation by trapping the ribosome on the mRNA initiation loading site. Proc. Natl. Acad. Sci. USA 1993, 90, 4394–4398. [Google Scholar] [CrossRef] [Green Version]
- Ehresmann, C.; Philippe, C.; Westhof, E.; Benard, L.; Portier, C.; Ehresmann, B. A pseudoknot is required for efficient translation initiation and regulation of the E. coli rpsO gene coding for ribosomal protein S15. Biochem. Cell Biol. 1995, 73, 1131–1140. [Google Scholar] [CrossRef]
- Bénard, L.; Mathy, N.; Grunberg-Manago, M.; Ehresmann, B.; Ehresmann, C.; Portier, C. Identification in a pseudoknot of a U.G motif essential for the regulation of the expression of ribosomal protein S15. Proc. Natl. Acad. Sci. USA 1998, 95, 2564–2567. [Google Scholar] [CrossRef] [Green Version]
- Serganov, A.; Ennifar, E.; Portier, C.; Ehresmann, B.; Ehresmann, C. Do mRNA and rRNA binding sites of E. coli ribosomal protein S15 share common structural determinants? J. Mol. Biol. 2002, 320, 963–978. [Google Scholar] [CrossRef]
- Mathy, N.; Pellegrini, O.; Serganov, A.; Patel, D.J.; Ehresmann, C.; Portier, C. Specific recognition of rpsO mRNA and 16S rRNA by Escherichia coli ribosomal protein S15 relies on both mimicry and site differentiation. Mol. Microbiol. 2004, 52, 661–675. [Google Scholar] [CrossRef]
- Marzi, S.; Myasnikov, A.G.; Serganov, A.; Ehresmann, C.; Romby, P.; Yusupov, M.; Klaholz, B.P. Structured mRNAs regulate translation initiation by binding to the platform of the ribosome. Cell 2007, 130, 1019–1031. [Google Scholar] [CrossRef]
- Scott, L.G.; Williamson, J.R. Interaction of the Bacillus stearothermophilus ribosomal protein S15 with its 5′-translational operator mRNA. J. Mol. Biol. 2001, 314, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.G.; Williamson, J.R. The binding interface between Bacillus stearothermophilus ribosomal protein S15 and its 5’-translational operator mRNA. J. Mol. Biol. 2005, 351, 280–290. [Google Scholar] [CrossRef]
- Slinger, B.L.; Meyer, M.M. RNA regulators responding to ribosomal protein S15 are frequent in sequence space. Nucleic Acids Res. 2016, 44, 9331–9341. [Google Scholar] [CrossRef] [Green Version]
- Serganov, A.; Polonskaia, A.; Ehresmann, B.; Ehresmann, C.; Patel, D.J. Ribosomal protein S15 represses its own translation via adaptation of an rRNA-like fold within its mRNA. EMBO J. 2003, 22, 1898–1908. [Google Scholar] [CrossRef] [Green Version]
- Slinger, B.L.; Deiorio-Haggar, K.; Anthony, J.S.; Gilligan, M.M.; Meyer, M.M. Discovery and validation of novel and distinct RNA regulators for ribosomal protein S15 in diverse bacterial phyla. BMC Genom. 2014, 15, 657. [Google Scholar] [CrossRef] [Green Version]
- Slinger, B.L.; Newman, H.; Lee, Y.; Pei, S.; Meyer, M.M. Coevolution of bacterial ribosomal protein S15 with diverse mRNA regulatory structures. PLoS Genet. 2015, 11, e1005720. [Google Scholar] [CrossRef] [Green Version]
- Bashiri, G.; Baker, E.N. Production of recombinant proteins in Mycobacterium smegmatis for structural and functional studies. Protein Sci. 2015, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; de la Cruz, V.F.; Fuerst, T.R.; Burlein, J.E.; Benson, L.A.; Bennett, L.T.; Bansal, G.P.; Young, J.F.; Lee, M.H.; Hatfull, G.F.; et al. New use of BCG for recombinant vaccines. Nature 1991, 351, 456–460. [Google Scholar] [CrossRef]
- Dreyfus, M. What constitutes the signal for the initiation of protein synthesis on Escherichia coli mRNAs? J. Mol. Biol. 1988, 204, 79–94. [Google Scholar] [CrossRef]
- Tchufistova, L.S.; Komarova, A.V.; Boni, I.V. A key role for the mRNA leader structure in translational control of ribosomal protein S1 synthesis in γ-proteobacteria. Nucleic Acids Res. 2003, 31, 6996–7002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aseev, L.V.; Levandovskaya, A.A.; Skaptsova, N.V.; Boni, I.V. Conservation of regulatory elements controlling the expression of the rpsB-tsf operon in γ-proteobacteria. Mol. Biol. 2009, 43, 101–107. [Google Scholar] [CrossRef]
- Bashyam, M.D.; Tyagi, A.K. Identification and analysis of “extended -10” promoters from mycobacteria. J. Bacteriol. 1998, 180, 2568–2573. [Google Scholar] [CrossRef] [Green Version]
- Arnvig, K.B.; Gopal, B.; Papavinasasundaram, K.G.; Cox, R.A.; Colston, M.J. The mechanism of upstream activation in the rrnB operon of Mycobacterium smegmatis is different from the Escherichia coli paradigm. Microbiology 2005, 151, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, N.; Tyagi, A.K. Mycobacterial transcriptional signals: Requirements for recognition by RNA polymerase and optimal transcriptional activity. Nucleic Acids Res. 2006, 34, 4245–4257. [Google Scholar] [CrossRef]
- Satchidanandam, V.; Amara, R.R.; Uchil, P.D.; Singh, V. The regulatory elements of the Mycobacterium tuberculosis gene Rv3881c function efficiently in Escherichia coli. FEMS Microbiol. Lett. 2003, 218, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Cortes, T.; Cox, R.A. Transcription and translation of the rpsJ, rplN and rRNA operons of the tubercle bacillus. Microbiology 2015, 161, 719–728. [Google Scholar] [CrossRef]
- Sawyer, E.B.; Grabowska, A.D.; Cortes, T. Translation regulation in mycobacteria and its implications for pathogenicity. Nucleic Acids Res. 2018, 46, 6950–6961. [Google Scholar] [CrossRef]
- Sawyer, E.B.; Phelan, J.E.; Clark, T.G.; Cortes, T. A snapshot of translation in Mycobacterium tuberculosis during exponential growth and nutrient starvation revealed by ribosome profiling. Cell Rep. 2021, 34, 108695. [Google Scholar] [CrossRef]
- Srivastava, A.; Asahara, H.; Zhang, M.; Zhang, W.; Liu, H.; Cui, S.; Jin, Q.; Chong, S. Reconstitution of protein translation of Mycobacterium reveals functional conservation and divergence with the Gram-negative bacterium Escherichia coli. PLoS ONE 2016, 11, e0162020. [Google Scholar] [CrossRef]
- Newton-Foot, M.; Gey van Pittius, N.C. The complex architecture of mycobacterial promoters. Tuberculosis 2013, 93, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Cortes, T.; Schubert, O.T.; Rose, G.; Arnvig, K.B.; Comas, I.; Aebersold, R.; Douglas, B. Genome-wide mapping of transcriptional start sites defines an extensive leaderless transcriptome in Mycobacterium tuberculosis. Cell Rep. 2013, 5, 1121–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aseev, L.V.; Koledinskaya, L.S.; Boni, I.V. Dissecting the extended “-10” Escherichia coli rpsB promoter activity and regulation in vivo. Biochemistry 2014, 79, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Bon, M.; Micheletti, C.; Orland, H. McGenus: A Monte Carlo algorithm to predict RNA secondary structures with pseudoknots. Nucleic Acids Res. 2013, 41, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Conn, A.B.; Diggs, S.; Tam, T.K.; Blaha, G.M. Two old dogs, one new trick: A review of RNA polymerase and ribosome interactions during transcription-translation coupling. Int. J. Mol. Sci. 2019, 20, 2595. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Fredrick, K. RNA polymerase’s relationship with the ribosome: Not so physical, most of the time. J. Mol. Biol. 2020, 432, 3981–3986. [Google Scholar] [CrossRef] [PubMed]
- Boni, I.V.; Isaeva, D.M.; Musychenko, M.L.; Tzareva, N.V. Ribosome-messenger recognition: mRNA target sites for ribosomal protein S1. Nucleic Acids Res. 1991, 19, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Komarova, A.V.; Tchufistova, L.S.; Supina, E.V.; Boni, I.V. Protein S1 counteracts the inhibitory effect of the extended Shine-Dalgarno sequence on translation. RNA 2002, 8, 1137–1147. [Google Scholar] [CrossRef] [Green Version]
- Duval, M.; Korepanov, A.; Fuchsbauer, O.; Fechter, P.; Haller, A.; Fabbretti, A.; Choulier, L.; Micura, R.; Klaholz, B.P.; Romby, P.; et al. Escherichia coli ribosomal protein S1 unfolds structured mRNAs onto the ribosome for active translation initiation. PLoS Biol. 2013, 11, e1001731. [Google Scholar] [CrossRef] [Green Version]
- Lund, P.E.; Chatterjee, S.; Daher, M.; Walter, N.G. Protein unties the pseudoknot: S1-mediated unfolding of RNA higher order structure. Nucleic Acids Res. 2020, 48, 2107–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, J.R.; Bulter, T.; Shen, C.R.; Liao, J.C. Directed evolution of ribosomal protein S1 for enhanced translational efficiency of high GC Rhodopseudomonas palustris DNA in Escherichia coli. J. Biol. Chem. 2007, 282, 18929–18936. [Google Scholar] [CrossRef] [Green Version]
- McGuffin, L.J.; Adiyaman, R.; Maghrabi, A.H.A.; Shuid, A.N.; Brackenridge, D.A.; Nealon, J.O.; Philomina, L.S. IntFOLD: An integrated web resource for high performance protein structure and function prediction. Nucleic Acids Res. 2019, 47, W408–W413. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.J.; Wu, C.H.; Yeh, A.Y.; Wen, J.D. Folding a stable RNA pseudoknot through rearrangement of two hairpin structures. Nucleic Acids Res. 2014, 42, 4505–4515. [Google Scholar] [CrossRef] [PubMed]
- Šiková, M.; Janoušková, M.; Ramaniuk, O.; Páleníková, P.; Pospíšil, J.; Bartl, P.; Suder, A.; Pajer, P.; Kubičková, P.; Pavliš, O.; et al. Ms1 RNA increases the amount of RNA polymerase in Mycobacterium smegmatis. Mol. Microbiol. 2019, 11, 354–372. [Google Scholar] [CrossRef]
- Gerrick, E.R.; Barbier, T.; Chase, M.R.; Xu, R.; François, J.; Lin, V.H.; Szucs, M.J.; Rock, J.M.; Ahmad, R.; Tjaden, B.; et al. Small RNA profiling in Mycobacterium tuberculosis identifies MrsI as necessary for an anticipatory iron sparing response. Proc. Natl. Acad. Sci. USA 2018, 115, 6464–6469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Mei, H.; Chen, F.; Tang, Q.; Yu, Z.; Cao, X.; Andongma, B.T.; Chou, S.H.; He, J. Transcriptome landscape of Mycobacterium smegmatis. Front. Microbiol. 2017, 8, 2505. [Google Scholar] [CrossRef] [Green Version]
- Kapopoulou, A.; Lew, J.M.; Cole, S.T. The MycoBrowser portal: A comprehensive and manually annotated resource for mycobacterial genomes. Tuberculosis 2011, 91, 8–13. [Google Scholar] [CrossRef]
- Snapper, S.B.; Melton, R.E.; Mustafa, S.; Kieser, T.; Jacobs, J.W.R. Isolation and characterization of efficient plasmid transformation mutants of Mycobacterium smegmatis. Mol. Microbiol. 1990, 4, 1911–1919. [Google Scholar] [CrossRef] [PubMed]
- Belisle, J.T.; Mahaffey, S.B.; Hill, P.J. Isolation of mycobacterium species genomic DNA. Methods Mol. Biol. 2009, 465, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Le Derout, J.; Boni, I.V.; Regnier, P.; Hajnsdorf, E. Hfq affects mRNA levels independently of degradation. BMC Mol. Biol. 2010, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, A.C.; Cohen, S.N. Construction and characterization of amplifiable multicopy DNA cloning vehicles derived from the P15A cryptic miniplasmid. J. Bacteriol. 1978, 134, 1141–1156. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Goude, R.; Roberts, D.M.; Parish, T. Electroporation in mycobacteria. In Mycobacteria Protocols. Methods in Molecular Biology; Parish, T., Roberts, D.M., Eds.; Springer Science+ Business Media: New York, NY, USA, 2015; Volume 1285. [Google Scholar] [CrossRef] [Green Version]
- Rabodoarivelo, M.S.; Aerts, M.; Vandamme, P.; Palomino, J.C.; Rasolofo, V.; Martin, A. Optimizing of a protein extraction method for Mycobacterium tuberculosis proteome analysis using mass spectrometry. J. Microbiol. Methods 2016, 131, 144–147. [Google Scholar] [CrossRef]
- Cormack, B.P.; Valdiva, R.H.; Falkow, S. FACS-optimized mutants of the green fluorescent protein (GFP) . Gene 1996, 173, 33–38. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Plasmid | Relevant Characteristics | Reference or Source |
|---|---|---|
| Strains | ||
| M. smegmatis mc2155 | [51] | |
| DH5a | E. coli cloning host | Laboratory stock |
| ENS0 | E. coli strain, his, formerly HfrG6D12 | [23] |
| IBrpsO188:lacZ | ENS0 bearing E. coli rpsO’-‘lacZ | [53] |
| LAB_PEcorpsOMsm:lacZ | ENS0 bearing Msm rpsO’-‘lacZ under E. coli rpsO promoter | This paper |
| Msm_PrpsOMsm:egfp | M. smegmatis bearing Msm rpsO’-egfp | This paper |
| Msm_PrpsOMsm:egfp | M. smegmatis bearing Msm rpsO’-egfp in the chromosome (Kanr) under the 5’ extended rpsO promoter (−231) | This paper |
| Msm_PrpsOMtb:egfp | M. smegmatis bearing Mtb rpsO’-egfp reporter in the chromosome (Kanr) under the Mtb rpsO promoter (−158) | This paper |
| Plasmids | ||
| pEMBL 46 | pEMBL8+derivative lacking lacZ RBS | [23] |
| pES15MsmTIR(v1) | pEMBL 46 derivative bearing rpsOMsm’-‘lacZ reporter under the Msm core rpsO promoter | This paper |
| pES15MsmTIR(v2) | pEMBL 46 derivative bearing rpsOMsm’-‘lacZ reporter under the E. coli rpsO promoter | This paper |
| pACYC184 | Tetr, Cmr, cloning vector | [54] |
| pQE30_egfp | derivative of pQE30 (Qiagen) expressing the egfp gene | Lukyanov KA# |
| pS15 (pS15Eco) | pACYC184 derivative expressing E. coli rpsO | [52] |
| pS15Msm(v1) | pACYC184 derivative expressing Msm rpsO under Msm rpsO core promoter and 5’UTR | This paper |
| pS15Msm(v2) | pACYC184 derivative expressing Msm rpsO under E. coli rpsO promoter and Msm 5’UTR | This paper |
| pS15Msm(v3) | pACYC184 derivative expressing Msm rpsO under E. coli rpsO promoter and 5’UTR -AUG | This paper |
| pAMYC | pACYC184 derivative carrying oriM | This paper |
| pAMS15Msm | pAMYC expressing Msm rpsO underthe Msm rpsO 5’-extended promoter (−231) | This paper |
| pAMS15Eco | pAMYC expressing E. coli rpsO | This paper |
| pMV261 | replicative shuttle vector, Kanr | [22] |
| pMV306hsp | integrative shuttle vector bearing hsp60 promoter and rrnB terminator, Kanr | [22] |
| pMVrpsOMsm:egfp | pMV306 derivatives bearing egfp fused with Msm rpsO 5’UTR under rpsO promoters differing in 5’ extensions (−47, −117 and −231 bp from TSS) | This paper |
| pMVrpsOMsmmut:egfp | pMV306 derivative bearing egfp fused with Msm rpsO 5’UTR harboring the mutated pseudoknot under the rpsO extended (−231) promoter | This paper |
| pMVrpsOMtb:egfp | pMV306 derivative bearing egfp fused with Mtb rpsO 5’UTR under the Mtb rpsO promoter (−158) | This paper |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aseev, L.V.; Koledinskaya, L.S.; Bychenko, O.S.; Boni, I.V. Regulation of Ribosomal Protein Synthesis in Mycobacteria: The Autogenous Control of rpsO. Int. J. Mol. Sci. 2021, 22, 9679. https://doi.org/10.3390/ijms22189679
Aseev LV, Koledinskaya LS, Bychenko OS, Boni IV. Regulation of Ribosomal Protein Synthesis in Mycobacteria: The Autogenous Control of rpsO. International Journal of Molecular Sciences. 2021; 22(18):9679. https://doi.org/10.3390/ijms22189679
Chicago/Turabian StyleAseev, Leonid V., Ludmila S. Koledinskaya, Oksana S. Bychenko, and Irina V. Boni. 2021. "Regulation of Ribosomal Protein Synthesis in Mycobacteria: The Autogenous Control of rpsO" International Journal of Molecular Sciences 22, no. 18: 9679. https://doi.org/10.3390/ijms22189679