
Gephyrin-Lacking PV Synapses on Neocortical Pyramidal Neurons

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Gephyrin as an Inhibitory Synapse Marker in Cortical Pyramidal Neurons
2.2. Gephyrin Colocalization at PV Synapses onto Pyr Neurons
2.3. Automated Alignment and Quantitation
2.4. Conjugate Immunofluorescence and Electron Array Tomography Confirm Gephyrin-Lacking Inhibitory Synapses
2.5. Quantitative Analysis of Inhibitory Synapses on Pyr Neurons
2.6. PV-Mediated Inhibition Is Functionally Stronger for L5 Pyr Neurons
2.7. PV Synapses onto PV Neurons Also Lack Postsynaptic Gephyrin
3. Discussion
3.1. Universal Marker for Inhibitory Synapses
3.2. Methodological Limitations
3.3. Pyr Neurons Show Heterogeneous Levels of Inhibition
3.4. Properties of Gephyrin-Lacking Synapses
4. Materials and Methods
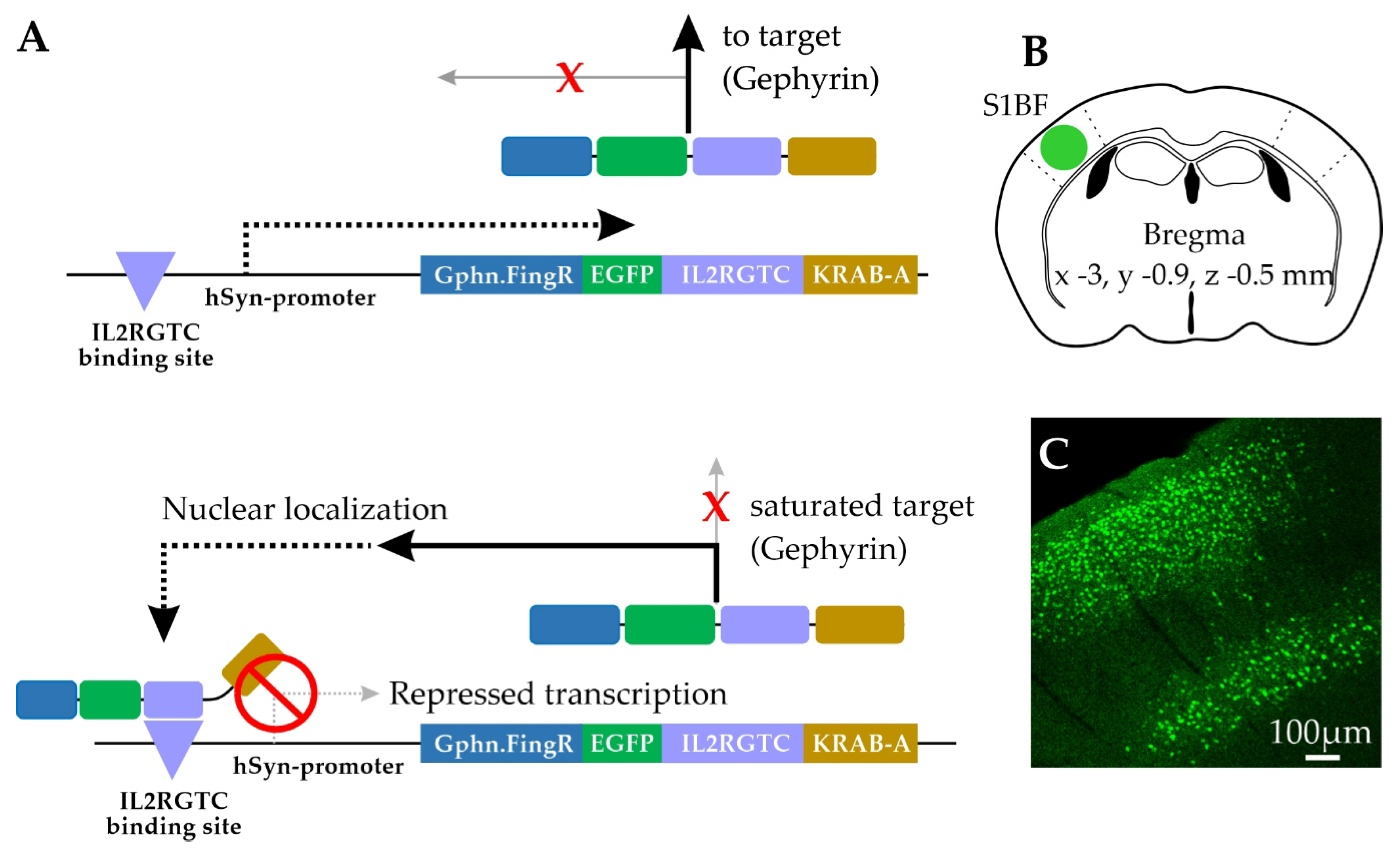
4.1. AAV Generation
4.2. Virus Production
4.3. Animals for Gephyrin.FingR Analysis
4.4. Cell Selection for Gephyrin.FingR Analysis
4.5. Gephyrin.FingR Immunolabeling Validation
4.6. Gephyrin.FingR Imaging
4.7. Gephyrin.FingR Image Analysis
4.8. Conjugate Immunofluorescence—SEM
4.9. Immunofluorescent Array Tomography
4.10. Image Processing and Analysis of Array Tomography
4.11. Antibodies
4.12. Electrophysiology
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kannan, M.; Gross, G.G.; Arnold, D.B.; Higley, M.J. Visual Deprivation During the Critical Period Enhances Layer 2/3 GABAergic Inhibition in Mouse V1. J. Neurosci. 2016, 36, 5914–5919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Oh, W.C.; Seong, J.; Kim, J. Advanced fluorescence protein-based synapse-detectors. Front. Synaptic Neurosci. 2016, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Kuljis, D.A.; Park, E.; Telmer, C.A.; Lee, J.; Ackerman, D.S.; Bruchez, M.P.; Barth, A.L. Fluorescence-based quantitative synapse analysis for cell-type specific connectomics. eNeuro 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Khayenko, V.; Maric, H.M. Targeting GABAAR-Associated proteins: New modulators, labels and concepts. Front. Mol. Neurosci. 2019, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Reshetniak, S.; Rizzoli, S.O. Interrogating Synaptic Architecture: Approaches for Labeling Organelles and Cytoskeleton Components. Front. Synaptic Neurosci. 2019, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Villa, K.L.; Berry, K.P.; Subramanian, J.; Cha, J.W.; Oh, W.C.; Kwon, H.B.; Kubota, Y.; So, P.T.C.; Nedivi, E. Inhibitory Synapses Are Repeatedly Assembled and Removed at Persistent Sites In Vivo. Neuron 2016, 89, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjulson, L.; Cassataro, D.; Dasgupta, S.; Miesenböck, G. Cell-Specific Targeting of Genetically Encoded Tools for Neuroscience. Annu. Rev. Genet. 2016, 50, 571–594. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Zhao, T.; Petralia, R.S.; Yu, Y.; Peng, H.; Myers, E.; Magee, J.C. mGRASP enables mapping mammalian synaptic connectivity with light microscopy. Nat. Methods 2011, 9, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Barth, A.L.; Ray, A. Progressive Circuit Changes during Learning and Disease. Neuron 2019, 104, 37–46. [Google Scholar] [CrossRef]
- Mishchenko, Y. On optical detection of densely labeled synapses in neuropil and mapping connectivity with combinatorially multiplexed fluorescent synaptic markers. PLoS ONE 2010, 5, e8853. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Shen, S.; Cadwell, C.R.; Berens, P.; Sinz, F.; Ecker, A.S.; Patel, S.; Tolias, A.S. Principles of connectivity among morphologically defined cell types in adult neocortex. Science (80-) 2015, 350, aac9462. [Google Scholar] [CrossRef] [Green Version]
- Fortin, D.A.; Tillo, S.E.; Yang, G.; Rah, J.; Melander, J.B.; Bai, S.; Soler-ceden, O.; Qin, M.; Zemelman, X.B.V.; Guo, C.; et al. Live Imaging of Endogenous PSD-95 Using ENABLED: A Conditional Strategy to Fluorescently Label Endogenous Proteins. J. Neurosci. 2014, 34, 16698–16712. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, A.; Li, H. CRASP: CFP reconstitution across synaptic partners. Biochem. Biophys. Res. Commun. 2016, 469, 352–356. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Zaharieva, E.E.; Kearney, P.J.; Alpert, M.H.; Lin, T.Y.; Turan, Z.; Lee, C.H.; Gallio, M. Dynamic labelling of neural connections in multiple colours by trans-synaptic fluorescence complementation. Nat. Commun. 2015, 6, 10024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.W.; Harris, J.A.; Ng, L.; Winslow, B.; Cain, N.; Mihalas, S.; Wang, Q.; Lau, C.; Kuan, L.; Henry, A.M.; et al. A mesoscale connectome of the mouse brain. Nature 2014, 508, 207–214. [Google Scholar] [CrossRef]
- Druckmann, S.; Feng, L.; Lee, B.; Yook, C.; Zhao, T.; Magee, J.C.; Kim, J. Structured Synaptic Connectivity between Hippocampal Regions. Neuron 2014, 81, 629–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassi, J.J.; Cepko, C.L.; Born, R.T.; Beier, K.T. Neuroanatomy goes viral! Front. Neuroanat. 2015, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Akin, O.; Nern, A.; Tsui, C.Y.K.; Pecot, M.Y.; Zipursky, S.L. Cell-type-specific labeling of synapses in vivo through synaptic tagging with recombination. Neuron 2014, 81, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Bemben, M.A.; Shipman, S.L.; Nicoll, R.A.; Roche, K.W. The cellular and molecular landscape of neuroligins. Trends Neurosci. 2015, 38, 496–505. [Google Scholar] [CrossRef]
- Kubota, Y.; Karube, F.; Nomura, M.; Kawaguchi, Y. The Diversity of Cortical Inhibitory Synapses. Front. Neural Circuits 2016, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Barberis, A. Postsynaptic plasticity of GABAergic synapses. Neuropharmacology 2020, 169, 107643. [Google Scholar] [CrossRef]
- Tyagarajan, S.K.; Fritschy, J.M. Gephyrin: A master regulator of neuronal function? Nat. Rev. Neurosci. 2014, 15, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Tretter, V.; Mukherjee, J.; Maric, H.M.; Schindelin, H.; Sieghart, W.; Moss, S.J. Gephyrin, the enigmatic organizer at GABAergic synapses. Front. Cell. Neurosci. 2012, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Kasaragod, V.B.; Schindelin, H. Structure–Function Relationships of Glycine and GABAA Receptors and Their Interplay With the Scaffolding Protein Gephyrin. Front. Mol. Neurosci. 2018, 11, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essrich, C.; Lorez, M.; Benson, J.A.; Fritschy, J.M.; Lüscher, B. Postsynaptic clustering of major GABAA receptor subtypes requires the γ2 subunit and gephyrin. Nat. Neurosci. 1998, 1, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Maric, H.M.; Mukherjee, J.; Tretter, V.; Moss, S.J.; Schindelin, H. Gephyrin-mediated γ-aminobutyric acid type A and glycine receptor clustering relies on a common binding site. J. Biol. Chem. 2011, 286, 42105–42114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choii, G.; Ko, J. Gephyrin: A central GABAergic synapse organizer. Exp. Mol. Med. 2015, 47, e158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groeneweg, F.L.; Trattnig, C.; Kuhse, J.; Nawrotzki, R.A.; Kirsch, J. Gephyrin: A key regulatory protein of inhibitory synapses and beyond. Histochem. Cell Biol. 2018, 150, 489–508. [Google Scholar] [CrossRef]
- Pizzarelli, R.; Griguoli, M.; Zacchi, P.; Petrini, E.M.; Barberis, A.; Cattaneo, A.; Cherubini, E. Tuning GABAergic Inhibition: Gephyrin Molecular Organization and Functions. Neuroscience 2020, 439, 125–136. [Google Scholar] [CrossRef]
- Chen, J.L.; Villa, K.L.; Cha, J.W.; So, P.T.C.; Kubota, Y.; Nedivi, E. Clustered Dynamics of Inhibitory Synapses and Dendritic Spines in the Adult Neocortex. Neuron 2012, 74, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Flores, C.E.; Nikonenko, I.; Mendez, P.; Fritschy, J.-M.; Tyagarajan, S.K.; Muller, D. Activity-dependent inhibitory synapse remodeling through gephyrin phosphorylation. Proc. Natl. Acad. Sci. USA 2015, 112, E65–E72. [Google Scholar] [CrossRef] [Green Version]
- Pennacchietti, F.; Vascon, S.; Nieus, T.; Rosillo, C.; Das, S.; Tyagarajan, S.K.; Diaspro, A.; del Bue, A.; Petrini, E.M.; Barberis, A.; et al. Nanoscale molecular reorganization of the inhibitory postsynaptic density is a determinant of gabaergic synaptic potentiation. J. Neurosci. 2017, 37, 1747–1756. [Google Scholar] [CrossRef] [PubMed]
- Gross, G.G.; Junge, J.A.; Mora, R.J.; Kwon, H.-B.B.; Olson, C.A.; Takahashi, T.T.; Liman, E.R.; Ellis-Davies, G.C.R.; McGee, A.W.; Sabatini, B.L.; et al. Recombinant Probes for Visualizing Endogenous Synaptic Proteins in Living Neurons. Neuron 2013, 78, 971–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensussen, S.; Shankar, S.; Ching, K.H.; Zemel, D.; Ta, T.L.; Mount, R.A.; Shroff, S.N.; Gritton, H.J.; Fabris, P.; Vanbenschoten, H.; et al. A Viral Toolbox of Genetically Encoded Fluorescent Synaptic Tags. iScience 2020, 23, 101330. [Google Scholar] [CrossRef]
- Son, J.H.; Keefe, M.D.; Stevenson, T.J.; Barrios, J.P.; Anjewierden, S.; Newton, J.B.; Douglass, A.D.; Bonkowsky, J.L. Transgenic FingRs for Live Mapping of Synaptic Dynamics in Genetically-Defined Neurons. Sci. Rep. 2016, 6, 18734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, C.K.; Xue, M.; He, M.; Huang, Z.J.; Scanziani, M. Inhibition of inhibition in visual cortex: The logic of connections between molecularly distinct interneurons. Nat. Neurosci. 2013, 16, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y.; Kondo, S.; Nomura, M.; Hatada, S.; Yamaguchi, N.; Mohamed, A.A.; Karube, F.; Lubke, J.; Kawaguchi, Y. Functional effects of distinct innervation styles of pyramidal cells by fast spiking cortical interneurons. eLife 2015, 4, e07919. [Google Scholar] [CrossRef] [Green Version]
- Hefft, S.; Jonas, P. Asynchronous GABA release generates long-lasting inhibition at a hippocampal interneuron-principal neuron synapse. Nat. Neurosci. 2005, 8, 1319–1328. [Google Scholar] [CrossRef]
- Maffei, A.; Nataraj, K.; Nelson, S.B.; Turrigiano, G.G. Potentiation of cortical inhibition by visual deprivation. Nature 2006, 443, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, C.D.; Zilberter, Y. Coincident spiking activity induces long-term changes in inhibition of neocortical pyramidal cells. J. Neurosci. 2001, 21, 8270–8277. [Google Scholar] [CrossRef]
- Vickers, E.D.; Clark, C.; Osypenko, D.; Fratzl, A.; Kochubey, O.; Bettler, B.; Schneggenburger, R. Parvalbumin-Interneuron Output Synapses Show Spike-Timing-Dependent Plasticity that Contributes to Auditory Map Remodeling. Neuron 2018, 99, 720–735.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daigle, T.L.; Madisen, L.; Hage, T.A.; Valley, M.T.; Knoblich, U.; Larsen, R.S.; Takeno, M.M.; Huang, L.; Gu, H.; Larsen, R.; et al. A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality. Cell 2018, 174, 465–480.e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouhanneau, J.S.; Kremkow, J.; Poulet, J.F.A. Single synaptic inputs drive high-precision action potentials in parvalbumin expressing GABA-ergic cortical neurons in vivo. Nat. Commun. 2018, 9, 1540. [Google Scholar] [CrossRef] [PubMed]
- Kuhlman, S.J.; Olivas, N.D.; Tring, E.; Ikrar, T.; Xu, X.; Trachtenberg, J.T. A disinhibitory microcircuit initiates critical-period plasticity in the visual cortex. Nature 2013, 501, 543–546. [Google Scholar] [CrossRef] [Green Version]
- Donato, F.; Rompani, S.B.; Caroni, P. Parvalbumin-expressing basket-cell network plasticity induced by experience regulates adult learning. Nature 2013, 504, 272–276. [Google Scholar] [CrossRef]
- Hippenmeyer, S.; Vrieseling, E.; Sigrist, M.; Portmann, T.; Laengle, C.; Ladle, D.R.; Arber, S. A developmental switch in the response of DRG neurons to ETS transcription factor signaling. PLoS Biol. 2005, 3, e159. [Google Scholar] [CrossRef] [Green Version]
- Rudy, B.; Fishell, G.; Lee, S.H.; Hjerling-Leffler, J. Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons. Dev. Neurobiol. 2011, 71, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Micheva, K.D.; Busse, B.; Weiler, N.C.; O’rourke, N.; Smith, S.J. Neurotechnique Single-Synapse Analysis of a Diverse Synapse Population: Proteomic Imaging Methods and Markers. Neuron 2010, 68, 639–653. [Google Scholar] [CrossRef] [Green Version]
- Takács, V.T.; Szőnyi, A.; Freund, T.F.; Nyiri, G.; Gulyás, A.I. Quantitative ultrastructural analysis of basket and axo-axonic cell terminals in the mouse hippocampus. Brain Struct. Funct. 2015, 220, 919–940. [Google Scholar] [CrossRef] [Green Version]
- Petrini, E.M.; Ravasenga, T.; Hausrat, T.J.; Iurilli, G.; Olcese, U.; Racine, V.; Sibarita, J.B.; Jacob, T.C.; Moss, S.J.; Benfenati, F.; et al. Synaptic recruitment of gephyrin regulates surface GABA A receptor dynamics for the expression of inhibitory LTP. Nat. Commun. 2014, 5, 3921. [Google Scholar] [CrossRef] [Green Version]
- Specht, C.G.; Izeddin, I.; Rodriguez, P.C.; ElBeheiry, M.; Rostaing, P.; Darzacq, X.; Dahan, M.; Triller, A. Quantitative nanoscopy of inhibitory synapses: Counting gephyrin molecules and receptor binding sites. Neuron 2013, 79, 308–321. [Google Scholar] [CrossRef] [Green Version]
- Crosby, K.C.; Gookin, S.E.; Garcia, J.D.; Hahm, K.M.; Dell’Acqua, M.L.; Smith, K.R. Nanoscale Subsynaptic Domains Underlie the Organization of the Inhibitory Synapse. Cell Rep. 2019, 26, 3284–3297.e3. [Google Scholar] [CrossRef] [Green Version]
- Sassoè-Pognetto, M.; Panzanelli, P.; Sieghart, W.; Fritschy, J.M. Colocalization of multiple GABA(A) receptor subtypes with gephyrin at postsynaptic sites. J. Comp. Neurol. 2000, 420, 481–498. [Google Scholar] [CrossRef]
- Freund, T.F.; Katona, I. Perisomatic Inhibition. Neuron 2007, 56, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, C.; Soltesz, I. Basket cell dichotomy in microcircuit function. J. Physiol. 2012, 590, 683–694. [Google Scholar] [CrossRef] [Green Version]
- Bartos, M.; Elgueta, C. Functional characteristics of parvalbumin- and cholecystokinin-expressing basket cells. J. Physiol. 2012, 590, 669–681. [Google Scholar] [CrossRef]
- Marlin, J.J.; Carter, A.G. GABA-A Receptor Inhibition of Local Calcium Signaling in Spines and Dendrites. J. Neurosci. 2014, 34, 15898–15911. [Google Scholar] [CrossRef] [PubMed]
- Jinno, S.; Kosaka, T. Parvalbumin is expressed in glutamatergic and GABAergic corticostriatal pathway in mice. J. Comp. Neurol. 2004, 477, 188–201. [Google Scholar] [CrossRef]
- Tanahira, C.; Higo, S.; Watanabe, K.; Tomioka, R.; Ebihara, S.; Kaneko, T.; Tamamaki, N. Parvalbumin neurons in the forebrain as revealed by parvalbumin-Cre transgenic mice. Neurosci. Res. 2009, 63, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Kosaka, T.; Singer, W.; Galuske, R.A.W. Gap junctions among dendrites of cortical GABAergic neurons establish a dense and widespread intercolumnar network. J. Neurosci. 2006, 26, 3434–3443. [Google Scholar] [CrossRef] [PubMed]
- Galarreta, M.; Hestrin, S. A network of fast-spiking cells in the neocortex connected by electrical synapses. Nature 1999, 402, 72–75. [Google Scholar] [CrossRef]
- Galarreta, M.; Hestrin, S. Electrical and chemical synapses among parvalbumin fast-spiking GABAergic interneurons in adult mouse neocortex. Proc. Natl. Acad. Sci. USA 2002, 99, 12438–12443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, J.R.; Belerlein, M.; Connors, B.W. Two networks of electrically coupled inhibitory neurons in neocortex. Nature 1999, 402, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, N.; Nishi, A.; Fukuda, T. Gap Junctions Interconnect Different Subtypes of Parvalbumin-Positive Interneurons in Barrels and Septa with Connectivity Unique to Each Subtype. Cereb. Cortex 2019, 29, 1414–1429. [Google Scholar] [CrossRef] [PubMed]
- Tamás, G.; Buhl, E.H.; Lörincz, A.; Somogyi, P. Proximally targeted GABAergic synapses and gap junctions synchronize cortical interneurons. Nat. Neurosci. 2000, 3, 366–371. [Google Scholar] [CrossRef]
- Hioki, H.; Okamoto, S.; Konno, M.; Kameda, H.; Sohn, J.; Kuramoto, E.; Fujiyama, F.; Kaneko, T. Cell type-specific inhibitory inputs to dendritic and somatic compartments of parvalbumin-expressing neocortical interneuron. J. Neurosci. 2013, 33, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Hioki, H. Compartmental organization of synaptic inputs to parvalbumin-expressing GABAergic neurons in mouse primary somatosensory cortex. Anat. Sci. Int. 2015, 90, 7–21. [Google Scholar] [CrossRef]
- Hioki, H.; Sohn, J.; Nakamura, H.; Okamoto, S.; Hwang, J.; Ishida, Y.; Takahashi, M.; Kameda, H. Preferential inputs from cholecystokinin-positive neurons to the somatic compartment of parvalbumin-expressing neurons in the mouse primary somatosensory cortex. Brain Res. 2018, 1695, 18–30. [Google Scholar] [CrossRef]
- Chiu, C.Q.; Barberis, A.; Higley, M.J. Preserving the balance: Diverse forms of long-term GABAergic synaptic plasticity. Nat. Rev. Neurosci. 2019, 20, 272–281. [Google Scholar] [CrossRef]
- Artinian, J.; Lacaille, J.C. Disinhibition in learning and memory circuits: New vistas for somatostatin interneurons and long-term synaptic plasticity. Brain Res. Bull. 2018, 141, 20–26. [Google Scholar] [CrossRef]
- Sheng, M.; Lin, J.W. Glutamatergic Synapses: Molecular Organization. eLS 2001. [Google Scholar] [CrossRef]
- Newpher, T.M.; Ehlers, M.D. Glutamate Receptor Dynamics in Dendritic Microdomains. Neuron 2008, 58, 472–497. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Levy, J.M.; Hou, A.; Winters, C.; Azzam, R.; Sousa, A.A.; Leapman, R.D.; Nicoll, R.A.; Reese, T.S. PSD-95 family MAGUKs are essential for anchoring AMPA and NMDA receptor complexes at the postsynaptic density. Proc. Natl. Acad. Sci. USA 2015, 112, E6983–E6992. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Cizeron, M.; Qiu, Z.; Benavides-Piccione, R.; Kopanitsa, M.V.; Skene, N.G.; Koniaris, B.; DeFelipe, J.; Fransén, E.; Komiyama, N.H.; et al. Architecture of the Mouse Brain Synaptome. Neuron 2018, 99, 781–799.e10. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W.; Sieghart, W. International Union of Pharmacology. LXX. Subtypes of γ-aminobutyric acidA receptors: Classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol. Rev. 2008, 60, 243–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engin, E.; Benham, R.S.; Rudolph, U. An Emerging Circuit Pharmacology of GABAA Receptors. Trends Pharmacol. Sci. 2018, 39, 710–732. [Google Scholar] [CrossRef] [PubMed]
- Tretter, V.; Jacob, T.C.; Mukherjee, J.; Fritschy, J.M.; Pangalos, M.N.; Moss, S.J. The clustering of GABAA receptor subtypes at inhibitory synapses is facilitated via the direct binding of receptor α2 subunits to gephyrin. J. Neurosci. 2008, 28, 1356–1365. [Google Scholar] [CrossRef] [Green Version]
- Tretter, V.; Kerschner, B.; Milenkovic, I.; Ramsden, S.L.; Ramerstorfer, J.; Saiepour, L.; Maric, H.M.; Moss, S.J.; Schindelin, H.; Harvey, R.J.; et al. Molecular basis of the γ-aminobutyric acid a receptor α3 subunit interaction with the clustering protein gephyrin. J. Biol. Chem. 2011, 286, 37702–37711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, J.; Kretschmannova, K.; Gouzer, G.; Maric, H.M.; Ramsden, S.; Tretter, V.; Harvey, K.; Davies, P.A.; Triller, A.; Schindelin, H.; et al. The residence time of GABA ARs at inhibitory synapses is determined by direct binding of the receptor α1 subunit to gephyrin. J. Neurosci. 2011, 31, 14677–14687. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, S.; Winkelmann, A.; Smolinsky, B.; Förstera, B.; Neundorf, I.; Schwarz, G.; Meier, J.C. Direct binding of GABAA receptor β2 and β3 subunits to gephyrin. Eur. J. Neurosci. 2013, 37, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Poulopoulos, A.; Aramuni, G.; Meyer, G.; Soykan, T.; Hoon, M.; Papadopoulos, T.; Zhang, M.; Paarmann, I.; Fuchs, C.; Harvey, K.; et al. Neuroligin 2 Drives Postsynaptic Assembly at Perisomatic Inhibitory Synapses through Gephyrin and Collybistin. Neuron 2009, 63, 628–642. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, T.; Soykan, T. The role of collybistin in gephyrin clustering at inhibitory synapses: Facts and open questions. Front. Cell. Neurosci. 2011, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kins, S.; Betz, H.; Kirsch, J. Collybistin, a newly identified brain-specific GEF, induces submembrane clustering of gephyrin. Nat. Neurosci. 2000, 3, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kneussel, M.; Helmut Brandstätter, J.; Gasnier, B.; Feng, G.; Sanes, J.R.; Betz, H. Gephyrin-independent clustering of postsynaptic GABAA receptor subtypes. Mol. Cell. Neurosci. 2001, 17, 973–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévi, S.; Logan, S.M.; Tovar, K.R.; Craig, A.M. Gephyrin Is Critical for Glycine Receptor Clustering but Not for the Formation of Functional GABAergic Synapses in Hippocampal Neurons. J. Neurosci. 2004, 24, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Viltono, L.; Patrizi, A.; Fritschy, J.M.; Sassoe-Pognetto, M.; Sassoè-Pognetto, M. Synaptogenesis in The Cerebellar Cortex: Differential Regulation of Gephyrin and GABAA Receptors At Somatic and Dendritic Synapses of Purkinje Cells. J. Comp. Neurol. 2008, 591, 579–591. [Google Scholar] [CrossRef]
- Ichikawa, R.; Yamasaki, M.; Miyazaki, T.; Konno, K.; Hashimoto, K.; Tatsumi, H.; Inoue, Y.; Kano, M.; Watanabe, M. Developmental switching of perisomatic innervation from climbing fibers to basket cell fibers in cerebellar Purkinje cells. J. Neurosci. 2011, 31, 16916–16927. [Google Scholar] [CrossRef] [Green Version]
- Briatore, F.; Pregno, G.; Di Angelantonio, S.; Frola, E.; De Stefano, M.E.; Vaillend, C.; Sassoè-Pognetto, M.; Patrizi, A. Dystroglycan Mediates Clustering of Essential GABAergic Components in Cerebellar Purkinje Cells. Front. Mol. Neurosci. 2020, 13, 164. [Google Scholar] [CrossRef]
- Papadopoulos, T.; Korte, M.; Eulenburg, V.; Kubota, H.; Retiounskaia, M.; Harvey, R.J.; Harvey, K.; O’Sullivan, G.A.; Laube, B.; Hülsmann, S.; et al. Impaired GABAergic transmission and altered hippocampal synaptic plasticity in collybistin-deficient mice. EMBO J. 2007, 26, 3888–3899. [Google Scholar] [CrossRef]
- Panzanelli, P.; Früh, S.; Fritschy, J.M. Differential role of GABAA receptors and neuroligin 2 for perisomatic GABAergic synapse formation in the hippocampus. Brain Struct. Funct. 2017, 222, 4149–4161. [Google Scholar] [CrossRef] [Green Version]
- Fritschy, J.M.; Panzanelli, P.; Tyagarajan, S.K. Molecular and functional heterogeneity of GABAergic synapses. Cell. Mol. Life Sci. 2012, 69, 2485–2499. [Google Scholar] [CrossRef] [Green Version]
- Panzanelli, P.; Gunn, B.G.; Schlatter, M.C.; Benke, D.; Tyagarajan, S.K.; Scheiffele, P.; Belelli, D.; Lambert, J.J.; Rudolph, U.; Fritschy, J.M. Distinct mechanisms regulate GABA A receptor and gephyrin clustering at perisomatic and axo-axonic synapses on CA1 pyramidal cells. J. Physiol. 2011, 589, 4959–4980. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, L.E.; Godin, A.G.; Wang, F.; St-Louis, M.; Carbonetto, S.; Wiseman, P.W.; Ribeiro-da-Silva, A.; De Koninck, Y. Gephyrin clusters are absent from small diameter primary afferent terminals despite the presence of GABAA receptors. J. Neurosci. 2014, 34, 8300–8317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Husseini, A.; Schnell, E.; Chetkovich, D. PSD-95 involvement in maturation of excitatory synapses. Science (80-) 2000, 290, 1364–1368. [Google Scholar] [CrossRef]
- Tsetsenis, T.; Boucard, A.A.; Arac, D.; Brunger, A.T.; Sudhof, T.C. Direct Visualization of Trans-Synaptic Neurexin-Neuroligin Interactions during Synapse Formation. J. Neurosci. 2014, 34, 15083–15096. [Google Scholar] [CrossRef] [Green Version]
- Takesian, A.E.; Bogart, L.J.; Lichtman, J.W.; Hensch, T.K. Inhibitory circuit gating of auditory critical-period plasticity. Nat. Neurosci. 2018, 21, 218–227. [Google Scholar] [CrossRef]
- Kameda, H.; Hioki, H.; Tanaka, Y.H.; Tanaka, T.; Sohn, J.; Sonomura, T.; Furuta, T.; Fujiyama, F.; Kaneko, T. Parvalbumin-producing cortical interneurons receive inhibitory inputs on proximal portions and cortical excitatory inputs on distal dendrites. Eur. J. Neurosci. 2012, 35, 838–854. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.S.; Maclachlan, C.; Blanc, J.; Dubois, A.; Petersen, C.C.H.; Knott, G.; Lee, S.H. 3D Ultrastructure of Synaptic Inputs to Distinct GABAergic Neurons in the Mouse Primary Visual Cortex. Cereb. Cortex 2021, 31, 2610–2624. [Google Scholar] [CrossRef]
- Micheva, K.D.; Beaulieu, C. An anatomical substrate for experience-dependent plasticity of the rat barrel field cortex. Proc. Natl. Acad. Sci. USA 1995, 92, 11834–11838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tixier-Vidal, A.; Faivre-Bauman, A.; Picart, R.; Wiedenmann, B. Immunoelectron microscopic localization of synaptophysin in a golgi subcompartment of developing hypothalamic neurons. Neuroscience 1988, 26, 847–861. [Google Scholar] [CrossRef]
- Leclerc, N.; Beesley, P.W.; Brown, I.; Colonnier, M.; Gurd, J.W.; Paladino, T.; Hawkes, R. Synaptophysin expression during synaptogenesis in the rat cerebellar cortex. J. Comp. Neurol. 1989, 280, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, T.; Paarmann, I.; Schmitt, B.; Haeger, S.; Sola, M.; Schmalzing, G.; Weissenhorn, W.; Betz, H. Molecular basis of gephyrin clustering at inhibitory synapses: Role of G- and E-domain interactions. J. Biol. Chem. 2007, 282, 5625–5632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kang, M.; Park, D.; Lee, A.R.; Betz, H.; Ko, J.; Chang, I.; Um, J.W. Impaired formation of high-order gephyrin oligomers underlies gephyrin dysfunction-associated pathologies. iScience 2021, 24, 102037. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Atallah, B.V.; Scanziani, M. Equalizing excitation-inhibition ratios across visual cortical neurons. Nature 2014, 511, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Tucciarone, J.; Padilla-coreano, N.; He, M.; Gordon, J.A.; Huang, Z.J. Selective inhibitory control of pyramidal neuron ensembles and cortical subnetworks by chandelier cells. Nat. Neurosci. 2017, 20, 1377–1383. [Google Scholar] [CrossRef] [Green Version]
- Tasic, B.; Yao, Z.; Graybuck, L.T.; Smith, K.A.; Nguyen, T.N.; Bertagnolli, D.; Goldy, J.; Garren, E.; Economo, M.N.; Viswanathan, S.; et al. Shared and distinct transcriptomic cell types across neocortical areas. Nature 2018, 563, 72–78. [Google Scholar] [CrossRef]
- Groh, A.; Bokor, H.; Mease, R.A.; Plattner, V.M.; Hangya, B.; Stroh, A.; Deschenes, M.; Acsády, L. Convergence of cortical and sensory driver inputs on single thalamocortical cells. Cereb. Cortex 2014, 24, 3167–3179. [Google Scholar] [CrossRef] [Green Version]
- Okabe, S.; Miwa, A.; Okado, H. Spine formation and correlated assembly of presynaptic and postsynaptic molecules. J. Neurosci. 2001, 21, 6105–6114. [Google Scholar] [CrossRef] [Green Version]
- Karunakaran, S.; Chowdhury, A.; Donato, F.; Quairiaux, C.; Michel, C.M.; Caroni, P. PV plasticity sustained through D1/5 dopamine signaling required for long-term memory consolidation. Nat. Neurosci. 2016, 19, 454–464. [Google Scholar] [CrossRef]
- Li, L.; Gainey, M.A.; Goldbeck, J.E.; Feldman, D.E. Rapid homeostasis by disinhibition during whisker map plasticity. Proc. Natl. Acad. Sci. USA 2014, 111, 1616–1621. [Google Scholar] [CrossRef] [Green Version]
- Wegner, W.; Ilgen, P.; Gregor, C.; Van Dort, J.; Mott, A.C.; Steffens, H.; Willig, K.I. In vivo mouse and live cell STED microscopy of neuronal actin plasticity using far-red emitting fluorescent proteins. Sci. Rep. 2017, 7, 11781. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.S.; Vandemark, K.; Mezey, D.; Shultz, N.; Fitzpatrick, D. Functional Synaptic Architecture of Callosal Inputs in Mouse Primary Visual Cortex. Neuron 2019, 101, 421–428.e5. [Google Scholar] [CrossRef] [Green Version]
- Collman, F.; Buchanan, J.; Phend, K.D.; Micheva, K.D.; Weinberg, R.J.; Smith, S.J. Mapping Synapses by Conjugate Light-Electron Array Tomography. J. Neurosci. 2015, 35, 5792–5807. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Madisen, L.; Mao, T.; Koch, H.; Zhuo, J.; Berenyi, A.; Fujisawa, S.; Hsu, Y.-W.A.; Garcia, A.J.; Gu, X.; Zanella, S.; et al. A toolbox of Cre-dependent optogenetic transgenic mice for light-induced activation and silencing. Nat. Neurosci. 2012, 15, 793–802. [Google Scholar] [CrossRef]
- Clem, R.L.; Barth, A. Pathway-specific trafficking of native AMPARs by in vivo experience. Neuron 2006, 49, 663–670. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Steps | Plasmid | Primer | DNA Insert | Endonuclease |
|---|---|---|---|---|
| Start | pAAV-hSyn-LA-mNeptune2 1 | |||
| 1. | pAAV-ZFN-LA-mNeptune2 | P-5′-CGCGTCTTCCACAGAGTGTGT-3′ | ZFN 2 | MluI |
| P-5′-CTAGACACACTCTGTGGAAGA-3′ | XbaI | |||
| 2. | pAAV-ZFN-hSyn-Gephyrin.FingR-EGFP-IL2RGTC-KRAB-A | 5′-TCATAGATCTGTGAGCAAGGGCGAGGAGC-3′ | EGFP | BglII |
| 5′-GACTAGATCTCTTGTACAGCTCGTCCATGC-3′ | ||||
| 5′-GAATTCGGTACCGCGGGCCCGGGA-3′ | GPHN-FingR-mKate2-IL2RGTC-KRAB-A 3 | KpnI BglII HindIII | ||
| 5′-GCGCCAAGCTTGCTTTACTTGTACGCTAA-3′ |
| Antigen | Host | Antibody Source | Dilution | RRID |
|---|---|---|---|---|
| Gephyrin | mouse | BD Biosciences 612632 | 1:100 | AB_399669 |
| Gephyrin | mouse | NeuroMab 75–443 | 1:100 | AB_2636851 |
| Gephyrin | mouse | Synaptic Systems 147 011C3 | 1:250 | AB_887716 |
| GABA | guinea pig | Millipore AB175 | 1:5000 | AB_91011 |
| GABAARα1 | Mouse | NeuroMab 75–136 | 1:100 | AB_2108811 |
| GAD2 | rabbit | Cell Signaling 5843 | 1:200 | AB_10835855 |
| Parvalbumin | rabbit | SWANT PV27 | 1:300 | AB_2631173 |
| Synapsin | rabbit | Cell Signaling 5297 | 1:200 | AB_ 2616578 |
| VGAT | mouse | Synaptic Systems 131 011 | 1:100 | AB_887868 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuljis, D.A.; Micheva, K.D.; Ray, A.; Wegner, W.; Bowman, R.; Madison, D.V.; Willig, K.I.; Barth, A.L. Gephyrin-Lacking PV Synapses on Neocortical Pyramidal Neurons. Int. J. Mol. Sci. 2021, 22, 10032. https://doi.org/10.3390/ijms221810032
Kuljis DA, Micheva KD, Ray A, Wegner W, Bowman R, Madison DV, Willig KI, Barth AL. Gephyrin-Lacking PV Synapses on Neocortical Pyramidal Neurons. International Journal of Molecular Sciences. 2021; 22(18):10032. https://doi.org/10.3390/ijms221810032
Chicago/Turabian StyleKuljis, Dika A., Kristina D. Micheva, Ajit Ray, Waja Wegner, Ryan Bowman, Daniel V. Madison, Katrin I. Willig, and Alison L. Barth. 2021. "Gephyrin-Lacking PV Synapses on Neocortical Pyramidal Neurons" International Journal of Molecular Sciences 22, no. 18: 10032. https://doi.org/10.3390/ijms221810032