
Influence of Vitamin D on the Vasoactive Effect of Estradiol in a Rat Model of Polycystic Ovary Syndrome

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PCOS Characteristics in Our Model
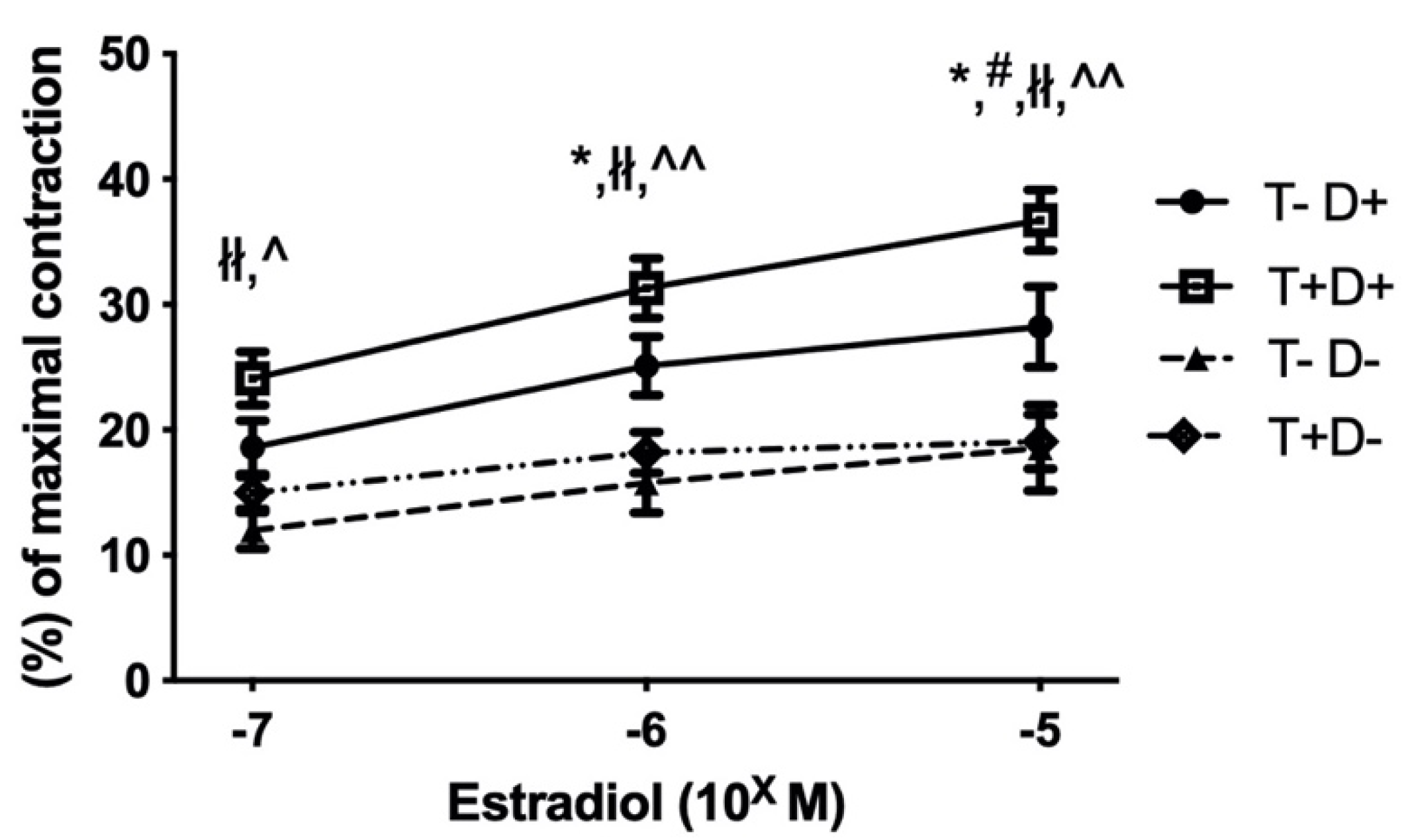
2.2. Estradiol Induced Relaxation of Isolated Thoracic Aorta Segments
2.3. Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Chronic Treatment of the Rats
4.4. Myography
4.5. Immunohistochemistry
4.6. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knochenhauer, E.S.; Key, T.J.; Kahsar-Miller, M.; Waggoner, W.; Boots, L.R.; Azziz, R. Prevalence of the polycystic ovary syndrome in unselected black and white women of the southeastern United States: A prospective study. J. Clin. Endocrinol. Metab. 1998, 83, 3078–3082. [Google Scholar] [CrossRef] [PubMed]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Dambala, K.; Paschou, S.A.; Michopoulos, A.; Siasos, G.; Goulis, D.G.; Vavilis, D.; Tarlatzis, B.C. Biomarkers of Endothelial Dysfunction in Women with Polycystic Ovary Syndrome. Angiology 2019, 70, 797–801. [Google Scholar] [CrossRef]
- Guastella, E.; Longo, R.A.; Carmina, E. Clinical and endocrine characteristics of the main polycystic ovary syndrome phenotypes. Fertil. Steril. 2010, 94, 2197–2201. [Google Scholar] [CrossRef] [PubMed]
- Sprung, V.; Atkinson, G.; Cuthbertson, D.; Pugh, C.J.A.; Aziz, N.; Green, D.J.; Cable, N.; Jones, H. Endothelial function measured using flow-mediated dilation in polycystic ovary syndrome: A meta-analysis of the observational studies. Clin. Endocrinol. 2013, 78, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Somani, Y.B.; Pawelczyk, J.; De Souza, M.J.; Kris-Etherton, P.; Proctor, D.N. Aging women and their endothelium: Probing the relative role of estrogen on vasodilator function. Am. J. Physiol. Circ. Physiol. 2019, 317, H395–H404. [Google Scholar] [CrossRef] [PubMed]
- Khalil, R.A. Estrogen, vascular estrogen receptor and hormone therapy in postmenopausal vascular disease. Biochem. Pharmacol. 2013, 86, 1627–1642. [Google Scholar] [CrossRef] [Green Version]
- Theodoratou, E.; Tzoulaki, I.; Zgaga, L.; Ioannidis, J.P.A. Vitamin D and multiple health outcomes: Umbrella review of systematic reviews and meta-analyses of observational studies and randomised trials. BMJ 2014, 348, g2035. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.; Peck, J.D.; Hansen, K.R.; Neas, B.; Craig, L.B. Associations between vitamin D levels and polycystic ovary syndrome phenotypes. Minerva Endocrinol. 2019, 44, 176–184. [Google Scholar] [CrossRef]
- Mogili, K.D.; Karuppusami, R.; Thomas, S.; Chandy, A.; Kamath, M.S.; Tk, A. Prevalence of vitamin D deficiency in infertile women with polycystic ovarian syndrome and its association with metabolic syndrome—A prospective observational study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 229, 15–19. [Google Scholar] [CrossRef]
- Yiu, Y.-F.; Chan, Y.-H.; Yiu, K.-H.; Siu, C.-W.; Li, S.-W.; Wong, L.-Y.; Lee, S.W.L.; Tam, S.; Wong, E.W.K.; Cheung, B.M.Y.; et al. Vitamin D Deficiency Is Associated with Depletion of Circulating Endothelial Progenitor Cells and Endothelial Dysfunction in Patients with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2011, 96, E830–E835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-H.; Meza, C.A.; Clarke, H.; Kim, J.-S.; Hickner, R.C. Vitamin D and Endothelial Function. Nutrients 2020, 12, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipos, M.; Péterffy, B.; Sziva, R.; Magyar, P.; Hadjadj, L.; Bányai, B.; Süli, A.; Soltész-Katona, E.; Gerszi, D.; Kiss, J.; et al. Vitamin D Deficiency Cause Gender Specific Alterations of Renal Arterial Function in a Rodent Model. Nutrients 2021, 13, 704. [Google Scholar] [CrossRef] [PubMed]
- Masszi, G.; Horváth, E.M.; Tarszabo, R.; Benko, R.; Novak, A.; Buday, A.; Tőkés, A.-M.; Nádasy, G.L.; Hamar, P.; Benyo, Z.; et al. Reduced Estradiol-Induced Vasodilation and Poly-(ADP-Ribose) Polymerase (PARP) Activity in the Aortas of Rats with Experimental Polycystic Ovary Syndrome (PCOS). PLoS ONE 2013, 8, e55589. [Google Scholar] [CrossRef] [PubMed]
- Lajtai, K.; Tarszabó, R.; Bányai, B.; Péterffy, B.; Gerszi, D.; Ruisanchez, É.; Sziva, R.E.; Korsós-Novák, Á.; Benkő, R.; Hadjadj, L.; et al. Effect of Vitamin D Status on Vascular Function of the Aorta in a Rat Model of PCOS. Oxid. Med. Cell. Longev. 2021, 2021, 8865979. [Google Scholar] [CrossRef]
- Hadjadj, L.; Várbíró, S.; Horváth, E.M.; Monori-Kiss, A.; Pál, E.; Karvaly, G.B.; Heinzlmann, A.; Magyar, A.; Szabo, I.; Sziva, R.E.; et al. Insulin resistance in an animal model of polycystic ovary disease is aggravated by vitamin D deficiency: Vascular consequences. Diabetes Vasc. Dis. Res. 2018, 15, 294–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pludowski, P.; Grant, W.; Bhattoa, H.P.; Bayer, M.; Povoroznyuk, V.; Rudenka, E.; Ramanau, H.; Varbiro, S.; Rudenka, A.; Karczmarewicz, E.; et al. Vitamin D Status in Central Europe. Int. J. Endocrinol. 2014, 2014, 589587. [Google Scholar] [CrossRef]
- Pludowski, P.; Holick, M.F.; Grant, W.B.; Konstantynowicz, J.; Mascarenhas, M.R.; Haq, A.; Povoroznyuk, V.; Balatska, N.; Barbosa, A.P.; Karonova, T.; et al. Vitamin D supplementation guidelines. J. Steroid Biochem. Mol. Biol. 2017, 175, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Masszi, G.; Buday, A.; Novak, A.; Horvath, E.M.; Tarszabo, R.; Sara, L.; Révész, C.; Benkő, R.; Nadasy, G.L.; Benyó, Z.; et al. Altered insulin-induced relaxation of aortic rings in a dihydrotestosterone-induced rodent model of polycystic ovary syndrome. Fertil. Steril. 2013, 99, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Masszi, G.; Novak, A.; Tarszabo, R.; Horvath, E.M.; Buday, A.; Ruisanchez, E.; Tokes, A.-M.; Sara, L.; Benko, R.; Nadasy, G.L.; et al. Effects of vitamin D3 derivative—Calcitriol on pharmacological reactivity of aortic rings in a rodent PCOS model. Pharmacol. Rep. 2013, 65, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Sara, L.; Antal, P.; Masszi, G.; Buday, A.; Horvath, E.M.; Hamar, P.; Monos, E.; Nadasy, G.L.; Varbiro, S. Arteriolar insulin resistance in a rat model of polycystic ovary syndrome. Fertil. Steril. 2012, 97, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sara, L.; Nádasy, G.; Antal, P.; Szekeres, M.; Monori-Kiss, A.; Horvath, E.M.; Tokes, A.-M.; Masszi, G.; Monos, E.; Varbiro, S. Arteriolar biomechanics in a rat polycystic ovary syndrome model—Effects of parallel vitamin D3 treatment. Acta Physiol. Hung. 2012, 99, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Sara, L.; Nadasy, G.L.; Antal, P.; Monori-Kiss, A.; Szekeres, M.; Masszi, G.; Monos, E.; Varbiro, S. Pharmacological reactivity of resistance vessels in a rat PCOS model—Vascular effects of parallel vitamin D3treatment. Gynecol. Endocrinol. 2012, 28, 961–964. [Google Scholar] [CrossRef]
- Hurliman, A.; Brown, J.K.; Maille, N.; Mandala, M.; Casson, P.; Osol, G. Hyperandrogenism and Insulin Resistance, Not Changes in Body Weight, Mediate the Development of Endothelial Dysfunction in a Female Rat Model of Polycystic Ovary Syndrome (PCOS). Endocrinology 2015, 156, 4071–4080. [Google Scholar] [CrossRef] [PubMed]
- Labruijere, S.; Houten, E.L.A.F.V.; De Vries, R.; Musterd-Bagghoe, U.M.; Garrelds, I.M.; Kramer, P.; Danser, A.H.J.; Villalón, C.M.; Visser, A.J.; Van Den Brink, M.A. Analysis of the vascular responses in a murine model of polycystic ovary syndrome. J. Endocrinol. 2013, 218, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannerås, L.; Cajander, S.; Holmäng, A.; Seleskovic, Z.; Lystig, T.; Lönn, M.; Stener-Victorin, E. A New Rat Model Exhibiting Both Ovarian and Metabolic Characteristics of Polycystic Ovary Syndrome. Endocrinology 2007, 148, 3781–3791. [Google Scholar] [CrossRef] [Green Version]
- Yanes, L.L.; Romero, D.G.; Moulana, M.; Lima, R.; Davis, D.D.; Zhang, H.; Lockhart, R.; Racusen, L.C.; Reckelhoff, J.F. Cardiovascular-Renal and Metabolic Characterization of a Rat Model of Polycystic Ovary Syndrome. Gend. Med. 2011, 8, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Pietro, M.; Pascuali, N.; Parborell, F.; Abramovich, D. Ovarian angiogenesis in polycystic ovary syndrome. Reproduction 2018, 155, R199–R209. [Google Scholar] [CrossRef] [Green Version]
- Miller, V.M.; Duckles, S.P. Vascular Actions of Estrogens: Functional Implications. Pharmacol. Rev. 2008, 60, 210–241. [Google Scholar] [CrossRef] [Green Version]
- Bucci, M.; Roviezzo, F.; Cicala, C.; Pinto, A.; Cirino, G. 17-beta-oestradiol-induced vasorelaxation in vitro is mediated by eNOS through hsp90 and akt/pkb dependent mechanism. Br. J. Pharmacol. 2002, 135, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.J.; Pencina, M.J.; Booth, S.L.; Jacques, P.F.; Ingelsson, E.; Lanier, K.; Benjamin, E.; D’Agostino, R.B.; Wolf, M.; Vasan, R.S. Vitamin D Deficiency and Risk of Cardiovascular Disease. Circulation 2008, 117, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Andrukhova, O.; Slavic, S.; Zeitz, U.; Riesen, S.C.; Heppelmann, M.S.; Ambrisko, T.D.; Markovic, M.; Kuebler, W.M.; Erben, R.G. Vitamin D Is a Regulator of Endothelial Nitric Oxide Synthase and Arterial Stiffness in Mice. Mol. Endocrinol. 2014, 28, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Xue, X.; Li, S.; Wang, Y.; Sun, Y.; Liu, W.; Yin, H.; Tao, T. Alterations of polyunsaturated fatty acid metabolism in ovarian tissues of polycystic ovary syndrome rats. J. Cell. Mol. Med. 2018, 22, 3388–3396. [Google Scholar] [CrossRef] [PubMed]
- Calkin, A.C.; Sudhir, K.; Honisett, S.; Williams, M.R.I.; Dawood, T.; Komesaroff, P.A. Rapid Potentiation of Endothelium-Dependent Vasodilation by Estradiol in Postmenopausal Women Is Mediated via Cyclooxygenase 2. J. Clin. Endocrinol. Metab. 2002, 87, 5072–5075. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.; Deb, S.; Sebastian, S.; Okamura, K.; Bulun, E.S. Estrogen up-regulates cyclooxygenase-2 via estrogen receptor in human uterine microvascular endothelial cells. Fertil. Steril. 2004, 81, 1351–1356. [Google Scholar] [CrossRef]
- Szymczak-Pajor, I.; Kleniewska, P.; Wieczfinska, J.; Pawliczak, R. Wide-Range Effects of 1,25(OH)2D3 on Group 4A Phospholipases Is Related to Nuclear Factor kappa-B and Phospholipase-A2 Activating Protein Activity in Mast Cells. Int. Arch. Allergy Immunol. 2020, 181, 56–70. [Google Scholar] [CrossRef]
- Luchetti, C.; Mikó, E.; Szekeres-Bartho, J.; Paz, D.; Motta, A. Dehydroepiandrosterone and metformin modulate progesterone-induced blocking factor (PIBF), cyclooxygenase 2 (COX2) and cytokines in early pregnant mice. J. Steroid Biochem. Mol. Biol. 2008, 111, 200–207. [Google Scholar] [CrossRef]
- Velez, L.M.; Heber, M.F.; Ferreira, S.R.; Abruzzese, A.G.; Reynoso, R.M.; Motta, A.B. Effect of hyperandrogenism on ovarian function. Reproduction 2015, 149, 577–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales, R.J.; Ghaffari, A.A.; Duckles, S.P.; Krause, D.N. Testosterone treatment increases thromboxane function in rat cerebral arteries. Am. J. Physiol. Circ. Physiol. 2005, 289, H578–H585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lajtai, K.; Nagy, C.T.; Tarszabó, R.; Benkő, R.; Hadjadj, L.; Sziva, R.E.; Gerszi, D.; Bányai, B.; Ferdinandy, P.; Nádasy, G.L.; et al. Effects of Vitamin D Deficiency on Proliferation and Autophagy of Ovarian and Liver Tissues in a Rat Model of Polycystic Ovary Syndrome. Biomolecules 2019, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Menichini, D.; Facchinetti, F. Effects of vitamin D supplementation in women with polycystic ovary syndrome: A review. Gynecol. Endocrinol. 2019, 36, 1–5. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarszabó, R.; Bányai, B.; Ruisanchez, É.; Péterffy, B.; Korsós-Novák, Á.; Lajtai, K.; Sziva, R.E.; Gerszi, D.; Hosszú, Á.; Benkő, R.; et al. Influence of Vitamin D on the Vasoactive Effect of Estradiol in a Rat Model of Polycystic Ovary Syndrome. Int. J. Mol. Sci. 2021, 22, 9404. https://doi.org/10.3390/ijms22179404
Tarszabó R, Bányai B, Ruisanchez É, Péterffy B, Korsós-Novák Á, Lajtai K, Sziva RE, Gerszi D, Hosszú Á, Benkő R, et al. Influence of Vitamin D on the Vasoactive Effect of Estradiol in a Rat Model of Polycystic Ovary Syndrome. International Journal of Molecular Sciences. 2021; 22(17):9404. https://doi.org/10.3390/ijms22179404
Chicago/Turabian StyleTarszabó, Róbert, Bálint Bányai, Éva Ruisanchez, Borbála Péterffy, Ágnes Korsós-Novák, Krisztina Lajtai, Réka Eszter Sziva, Dóra Gerszi, Ádám Hosszú, Rita Benkő, and et al. 2021. "Influence of Vitamin D on the Vasoactive Effect of Estradiol in a Rat Model of Polycystic Ovary Syndrome" International Journal of Molecular Sciences 22, no. 17: 9404. https://doi.org/10.3390/ijms22179404