
Muscimol Directly Activates the TREK-2 Channel Expressed in GABAergic Neurons through Its N-Terminus

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Expression of the GABA Receptor and K2P Channel mRNA in B35 Cells
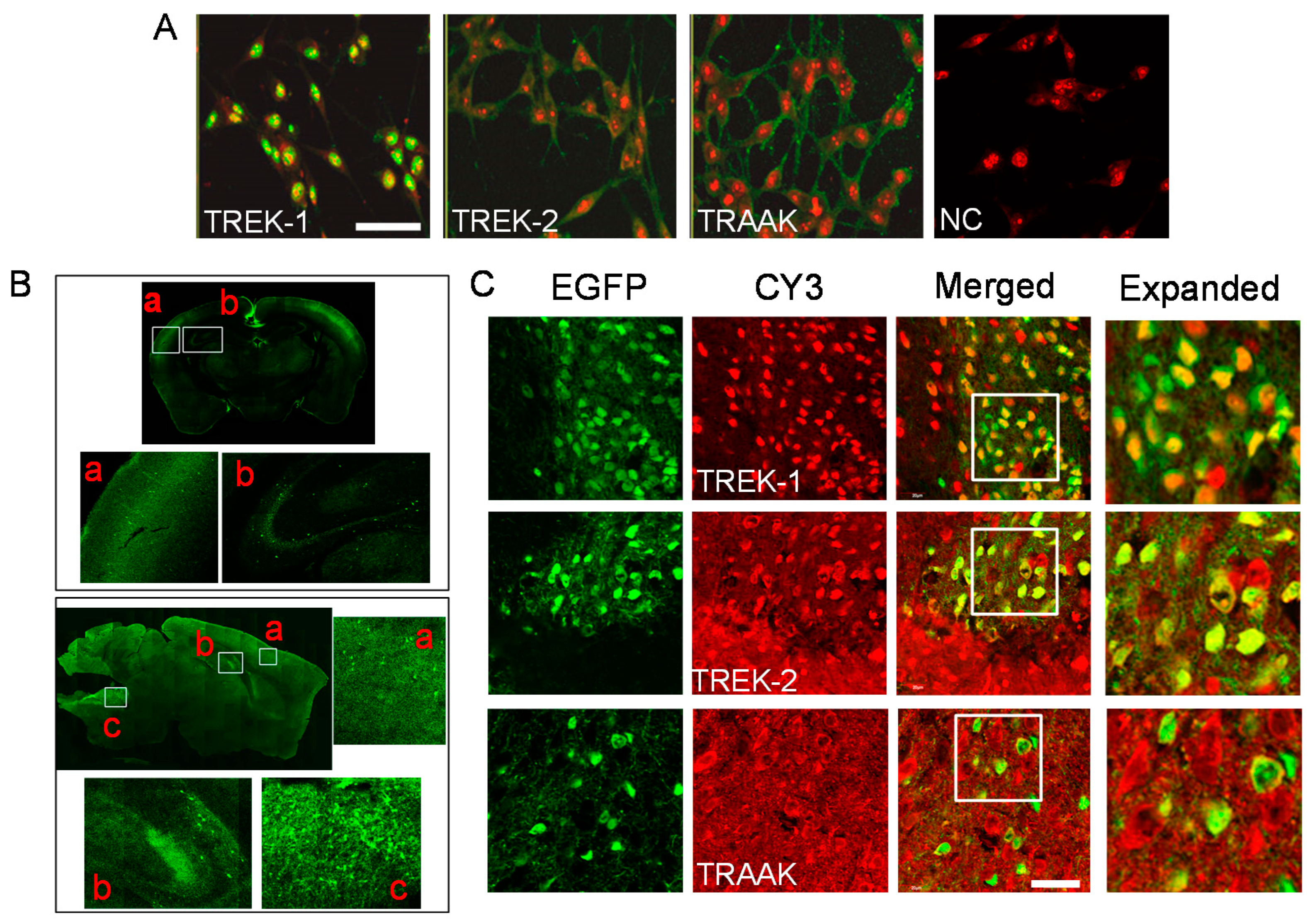
2.2. Localization of TREK/TRAAK Proteins in B35 Cells and GABAergic Neurons
2.3. TREK-2 Expression Levels Increased by GABA Receptor Agonists in B35 Cells
2.4. Activation of TREK-2 by Muscimol
3. Discussion
3.1. Up-Regulation of TREK-2 Expression in GABAergic Neurons by Muscimol
3.2. Activation of TREK-2 by GABAA Receptor Agonists
3.3. The Physiological Role of TREK-2 Activation in GABAergic Neurons
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Glutamate Decarboxylase 67 (GAD67)-GFP Knock-In Mice
4.4. RT-PCR and Real-Time PCR
4.5. Immunostaining
4.6. Western Blot Analysis
4.7. Constructs
4.8. Electrophysiological Studies
4.9. TREK-2 Channel Binding to a Fluorescent Muscimol
4.10. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Meng, D.; Li, H.Q.; Deisseroth, K.; Leutgeb, S.; Spitzer, N.C. Neuronal activity regulates neurotransmitter switching in the adult brain following light-induced stress. Proc. Natl. Acad. Sci. USA 2018, 115, 5064–5071. [Google Scholar] [CrossRef] [Green Version]
- Marder, E.; Prinz, A.A. Modeling stability in neuron and network function: The role of activity in homeostasis. Bioessays 2002, 24, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Kim, D. Physiology and pharmacology of two-pore domain potassium channels. Curr. Pharm. Des. 2005, 11, 2717–2736. [Google Scholar] [CrossRef]
- Kang, D.; Han, J.; Kim, D. Mechanism of Inhibition of TREK-2 (K2P10.1) by the Gq-coupled M3 muscarinic receptor. Am. J. Physiol. Cell Physiol. 2006, 291, C649–C656. [Google Scholar] [CrossRef]
- Talley, E.M.; Solorzano, G.; Lei, Q.; Kim, D.; Bayliss, D.A. Cns distribution of members of the two-pore-domain (KCNK) potassium channel family. J. Neurosci. 2001, 21, 7491–7505. [Google Scholar] [CrossRef] [PubMed]
- Hervieu, G.J.; Cluderay, J.E.; Gray, C.W.; Green, P.J.; Ranson, J.L.; Randall, A.D.; Meadows, H.J. Distribution and expression of TREK-1, a two-pore-domain potassium channel, in the adult rat CNS. Neuroscience 2001, 103, 899–919. [Google Scholar] [CrossRef]
- Deng, P.Y.; Xiao, Z.; Yang, C.; Rojanathammanee, L.; Grisanti, L.; Watt, J.; Geiger, J.D.; Liu, R.; Porter, J.E.; Lei, S. GABA(B) receptor activation inhibits neuronal excitability and spatial learning in the entorhinal cortex by activating TREK-2 K+ channels. Neuron 2009, 63, 230–243. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W.; DeLorey, T.M. GABA Receptor Physiology and Pharmacology; Siegel, G.J., Agranoff, B.W., Albers, R.W., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1999. [Google Scholar]
- Patel, A.J.; Honore, E.; Lesage, F.; Fink, M.; Romey, G.; Lazdunski, M. Inhalational anesthetics activate two-pore-domain background K+ channels. Nat. Neurosci. 1999, 2, 422–426. [Google Scholar] [CrossRef]
- Perouansky, M.; Pearce, R.A. Is anesthesia caused by potentiation of synaptic or intrinsic inhibition? Recent insights into the mechanisms of volatile anesthetics. J. Basic Clin. Physiol. Pharm. 2000, 11, 83–107. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.S.; Kolesky, S.E.; Jenkins, A. General anesthetic actions on GABA(A) receptors. Curr. Neuropharmacol. 2010, 8, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Schubert, D.; Heinemann, S.; Carlisle, W.; Tarikas, H.; Kimes, B.; Patrick, J.; Steinbach, J.H.; Culp, W.; Brandt, B.L. Clonal cell lines from the rat central nervous system. Nature 1974, 249, 224–227. [Google Scholar] [CrossRef]
- Tyndale, R.F.; Hales, T.G.; Olsen, R.W.; Tobin, A.J. Distinctive patterns of GABAA receptor subunit mRNAs in 13 cell lines. J. Neurosci. 1994, 14, 5417–5428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proks, P.; Schewe, M.; Conrad, L.J.; Rao, S.; Rathje, K.; Rodstrom, K.E.J.; Carpenter, E.P.; Baukrowitz, T.; Tucker, S.J. Norfluoxetine inhibits TREK-2 K2P channels by multiple mechanisms including state-independent effects on the selectivity filter gate. J. Gen. Physiol. 2021, 153, e202012812. [Google Scholar] [CrossRef] [PubMed]
- Aller, M.I.; Wisden, W. Changes in expression of some two-pore domain potassium channel genes (KCNK) in selected brain regions of developing mice. Neuroscience 2008, 151, 1154–1172. [Google Scholar] [CrossRef]
- Shi, S.Q.; Shi, Z.; Jiang, Z.P.; Qi, L.W.; Sun, X.M.; Li, C.X.; Liu, J.F.; Xiao, W.F.; Zhang, S.G. Effects of exogenous GABA on gene expression of Caragana intermedia roots under NaCl stress: Regulatory roles for H2O2 and ethylene production. Plant. Cell Environ. 2010, 33, 149–162. [Google Scholar] [CrossRef]
- Kanasaki, H.; Tumurbaatar, T.; Oride, A.; Hara, T.; Okada, H.; Kyo, S. Gamma-aminobutyric acidA receptor agonist, muscimol, increases KiSS-1 gene expression in hypothalamic cell models. Reprod. Med. Biol. 2017, 16, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Nuss, P. Anxiety disorders and GABA neurotransmission: A disturbance of modulation. Neuropsychiatr. Dis. Treat. 2015, 11, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Jie, W.; Huang, L.; Wei, P.; Li, S.; Luo, Z.; Friedman, A.K.; Meredith, A.L.; Han, M.H.; Zhu, X.H.; et al. Nuclear BK channels regulate gene expression via the control of nuclear calcium signaling. Nat. Neurosci. 2014, 17, 1055–1063. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.H.; Byun, J.K.; Jeon, W.I.; Choi, S.Y.; Park, J.; Lee, B.H.; Yang, J.E.; Park, J.B.; O’Grady, S.M.; Kim, D.Y.; et al. Nuclear localization and functional characteristics of voltage-gated potassium channel Kv1.3. J. Biol. Chem. 2015, 290, 12547–12557. [Google Scholar] [CrossRef] [Green Version]
- Labrakakis, C.; Muller, T.; Schmidt, K.; Kettenmann, H. GABA(A) receptor activation triggers a Cl- conductance increase and a K+ channel blockade in cerebellar granule cells. Neuroscience 1997, 79, 177–189. [Google Scholar] [CrossRef]
- von Blankenfeld, G.; Kettenmann, H. Glutamate and GABA receptors in vertebrate glial cells. Mol. Neurobiol. 1991, 5, 31–43. [Google Scholar] [CrossRef]
- Stelzer, A. Ion. Channels; Narahashi, T., Ed.; Springer Science+Business Media: New York, NY, USA, 1992; Volume 3, p. 54. [Google Scholar]
- Akk, G.; Germann, A.L.; Sugasawa, Y.; Pierce, S.R.; Evers, A.S.; Steinbach, J.H. Enhancement of Muscimol Binding and Gating by Allosteric Modulators of the GABAA Receptor: Relating Occupancy to State Functions. Mol. Pharmacol. 2020, 98, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Benkherouf, A.Y.; Taina, K.R.; Meera, P.; Aalto, A.J.; Li, X.G.; Soini, S.L.; Wallner, M.; Uusi-Oukari, M. Extrasynaptic delta-GABAA receptors are high-affinity muscimol receptors. J. Neurochem. 2019, 149, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vashchinkina, E.; Manner, A.K.; Vekovischeva, O.; den Hollander, B.; Uusi-Oukari, M.; Aitta-Aho, T.; Korpi, E.R. Neurosteroid Agonist at GABAA receptor induces persistent neuroplasticity in VTA dopamine neurons. Neuropsychopharmacology 2014, 39, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Franks, N.P.; Lieb, W.R. Molecular and cellular mechanisms of general anaesthesia. Nature 1994, 367, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Sirois, J.E.; Pancrazio, J.J.; Lynch, C., 3rd; Bayliss, D.A. Multiple ionic mechanisms mediate inhibition of rat motoneurones by inhalation anaesthetics. J. Physiol. 1998, 512, 851–862. [Google Scholar] [CrossRef]
- Talley, E.M.; Sirois, J.E.; Lei, Q.; Bayliss, D.A. Two-pore-Domain (KCNK) potassium channels: Dynamic roles in neuronal function. Neuroscientist 2003, 9, 46–56. [Google Scholar] [CrossRef]
- Hui, K.M.; Huen, M.S.; Wang, H.Y.; Zheng, H.; Sigel, E.; Baur, R.; Ren, H.; Li, Z.W.; Wong, J.T.; Xue, H. Anxiolytic effect of wogonin, a benzodiazepine receptor ligand isolated from Scutellaria baicalensis Georgi. Biochem. Pharmacol. 2002, 64, 1415–1424. [Google Scholar] [CrossRef]
- Kim, E.J.; Kang, D.; Han, J. Baicalein and wogonin are activators of rat TREK-2 two-pore domain K+ channel. Acta Physiol. 2011, 202, 185–192. [Google Scholar] [CrossRef]
- Yang, J.H.; Siregar, A.S.; Kim, E.J.; Nyiramana, M.M.; Shin, E.J.; Han, J.; Sohn, J.T.; Kim, J.W.; Kang, D. Involvement of TREK-1 Channel in Cell Viability of H9c2 Rat Cardiomyoblasts Affected by Bupivacaine and Lipid Emulsion. Cells 2019, 8, 454. [Google Scholar] [CrossRef] [Green Version]
- Ango, F.; di Cristo, G.; Higashiyama, H.; Bennett, V.; Wu, P.; Huang, Z.J. Ankyrin-based subcellular gradient of neurofascin, an immunoglobulin family protein, directs GABAergic innervation at purkinje axon initial segment. Cell 2004, 119, 257–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyaya, B.; Di Cristo, G.; Higashiyama, H.; Knott, G.W.; Kuhlman, S.J.; Welker, E.; Huang, Z.J. Experience and activity-dependent maturation of perisomatic GABAergic innervation in primary visual cortex during a postnatal critical period. J. Neurosci. 2004, 24, 9598–9611. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Curtis, C.; Tavare, S.; Tower, J. A screen of apoptosis and senescence regulatory genes for life span effects when over-expressed in Drosophila. Aging 2009, 1, 191–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, H.; Kim, Y.; Kim, D. TREK-2, a new member of the mechanosensitive tandem-pore K+ channel family. J. Biol. Chem. 2000, 275, 17412–17419. [Google Scholar] [CrossRef] [Green Version]
- Schnorbusch, K.; Lembrechts, R.; Pintelon, I.; Timmermans, J.P.; Brouns, I.; Adriaensen, D. GABAergic signaling in the pulmonary neuroepithelial body microenvironment: Functional imaging in GAD67-GFP mice. Histochem. Cell Biol. 2013, 140, 549–566. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Acc. No. | Primer Sequences (5′–3′) | Expected Size (bp) | Application |
|---|---|---|---|---|
| GAPDH | AF106860 | F: CTAAAGGGCATCCTGGGC R: TTACTCCTTGGAGGCCATG F: CATGGCCTTCCGTGTTC R: CTGCTTCACCACCTTCTT | 201 103 | RT-PCR Real-time PCR |
| Gabra1 (GABAA Receptor) | L08490 | F: ATCTTTGGGCCTGGACCCTC R: CGGGCTGGCTCCCTTGTCCA | 580 | RT-PCR |
| Gabbr1 (GABAB Receptor) | NM_031028 | F: GTCTGGAGGAGGTGGTCGTT R: ACAAACGGGAACTGGCTTCT | 534 | RT-PCR |
| Kcnk3 (TASK-1) | NM_033376 | F: TGTTCTGCATGTTCTACGCG R: TGGAGTACTGCAGCTTCTCG F: GAGCTGCCTAAGCGGTA R: AAAGTGCAGCATTTCAGCATA | 702 142 | RT-PCR Real-time PCR |
| Kcnk9 (TASK-3) | AF192366 | F: TGACTACTATAGGGTTCGGCG R: AAGTAGGTGTTCCTCAGCACG F: GAAGTTTCTATGGAGAACATGGTGA R: CGTGGAAGAAGCTCCAAT | 517 105 | RT-PCR Real-time PCR |
| Kcnk2 (TREK-1) | AF325671 | F: TGCCAAAGTGGAGGACACAT R: CTCTCCCACCTCTTCCTTCG F: TCTGAATGAATCAGAATGCTTTGCTA R: TCTGAATGAATCAGAATGCTTTGCTA | 361 106 | RT-PCR Real-time PCR |
| Kcnk10 (TREK-2) | AF196965 | F: CAGCCCAAGAGTGCCACTAA R: GGATCCCAAAGATGGCGTAT F: TCAGTATGATTGGAGACTGGC R: ACTCAGCAGTGACATTAGC | 493 107 | RT-PCR Real-time PCR |
| Kcnk4 (TRAAK) | AF302842 | F: CACCACTGTAGGCTTTGGCGATTATG R: ACTCTGCGTGTCTGAGGACTCGTCG F: CTGCCTGCTCTTTGTCC R: TACAGTGGTGAGTGTCACTATAAC | 445 103 | RT-PCR Real-time PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-J.; Kwon, O.-S.; Hur, C.-G.; Nyiramana, M.M.; Lee, D.-K.; Hong, S.-G.; Han, J.; Kang, D. Muscimol Directly Activates the TREK-2 Channel Expressed in GABAergic Neurons through Its N-Terminus. Int. J. Mol. Sci. 2021, 22, 9320. https://doi.org/10.3390/ijms22179320
Kim E-J, Kwon O-S, Hur C-G, Nyiramana MM, Lee D-K, Hong S-G, Han J, Kang D. Muscimol Directly Activates the TREK-2 Channel Expressed in GABAergic Neurons through Its N-Terminus. International Journal of Molecular Sciences. 2021; 22(17):9320. https://doi.org/10.3390/ijms22179320
Chicago/Turabian StyleKim, Eun-Jin, Oh-Sang Kwon, Chang-Gi Hur, Marie Merci Nyiramana, Dong-Kun Lee, Seong-Geun Hong, Jaehee Han, and Dawon Kang. 2021. "Muscimol Directly Activates the TREK-2 Channel Expressed in GABAergic Neurons through Its N-Terminus" International Journal of Molecular Sciences 22, no. 17: 9320. https://doi.org/10.3390/ijms22179320