
Hyperbilirubinemia Maintained by Chronic Supplementation of Unconjugated Bilirubin Improves the Clinical Course of Experimental Autoimmune Arthritis

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Bilirubin Levels
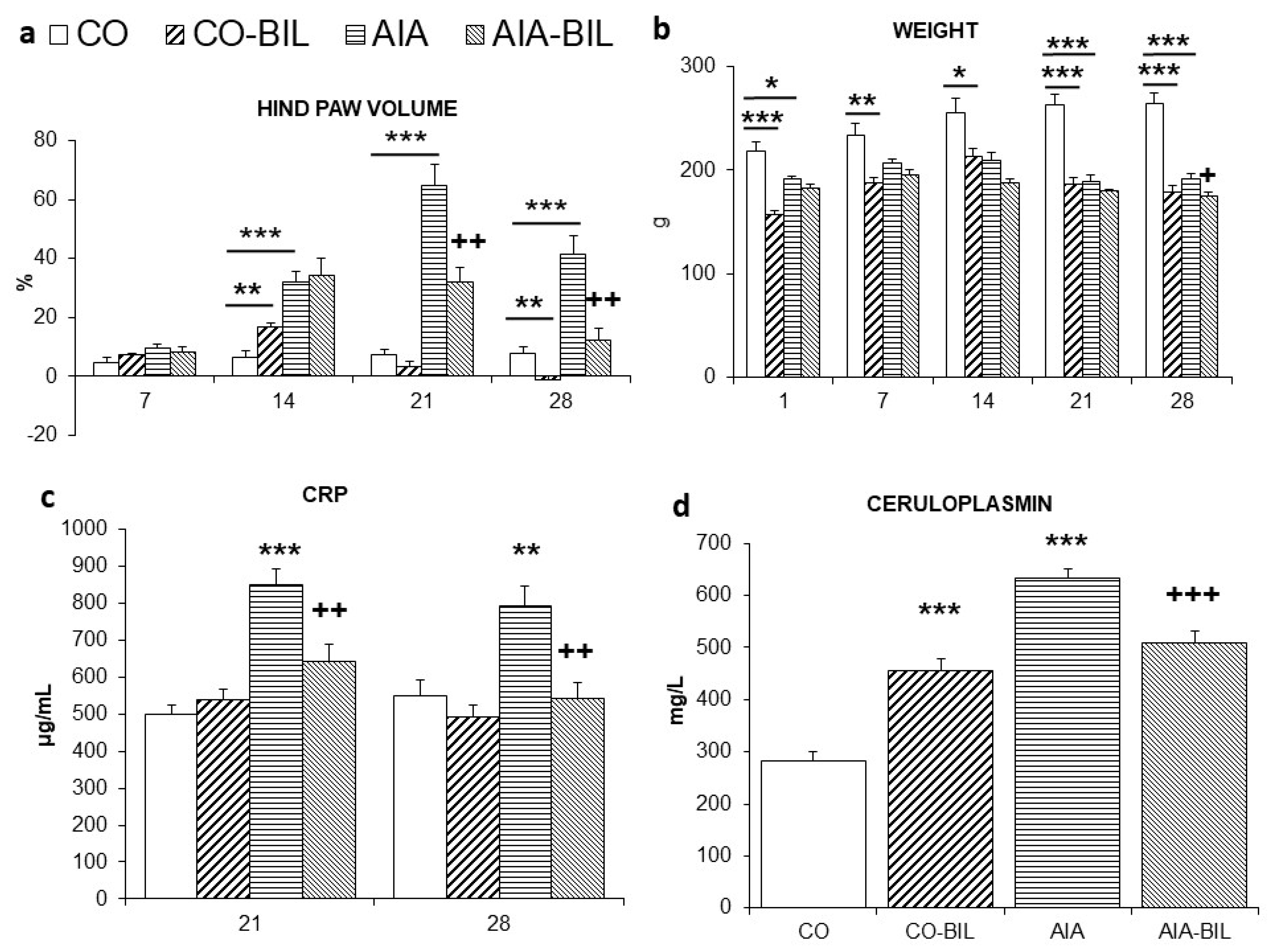
2.2. Changes in Hind Paw Volume and Body Weight
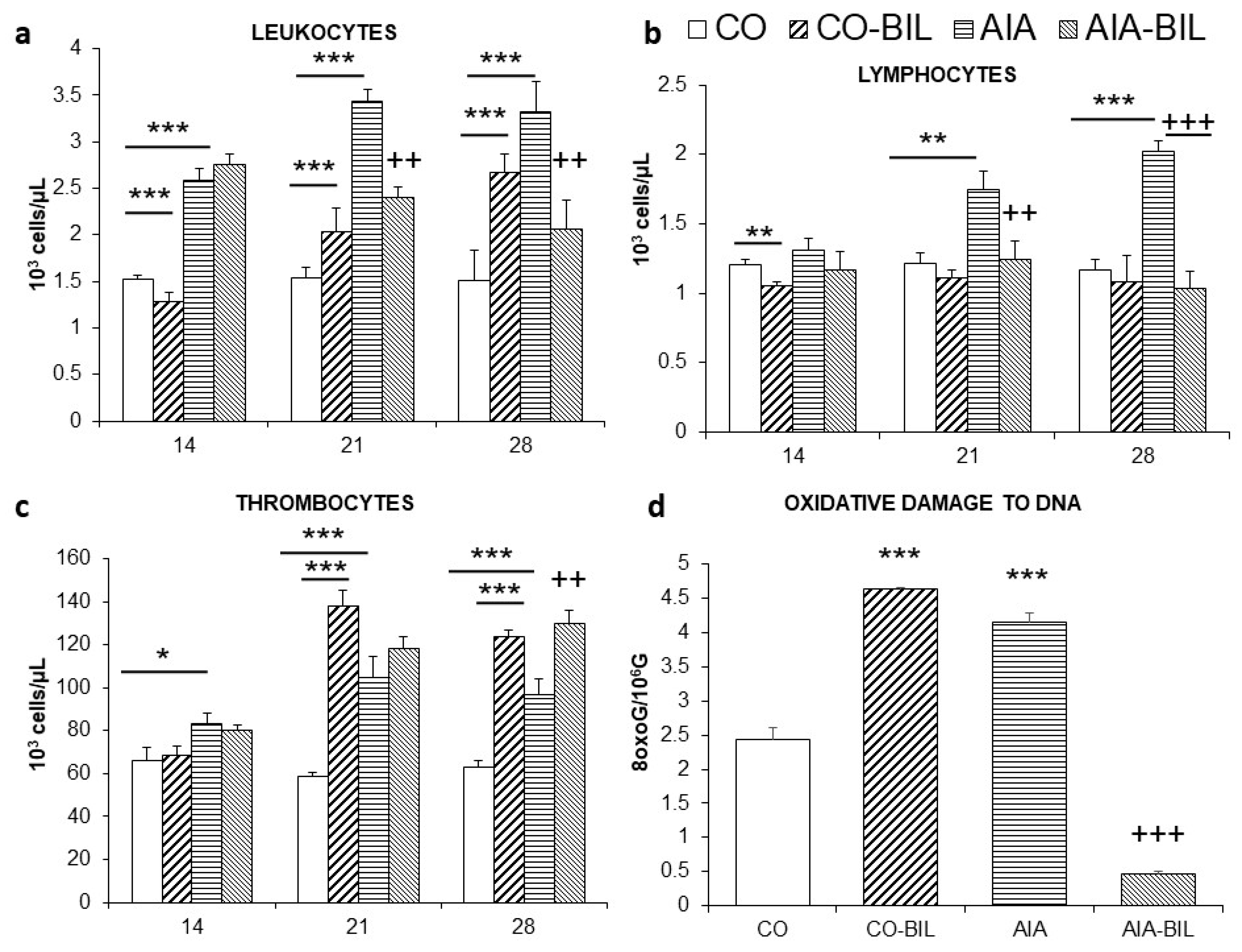
2.3. Changes in Blood Screen
2.4. Markers of Inflammation and Organ Weight
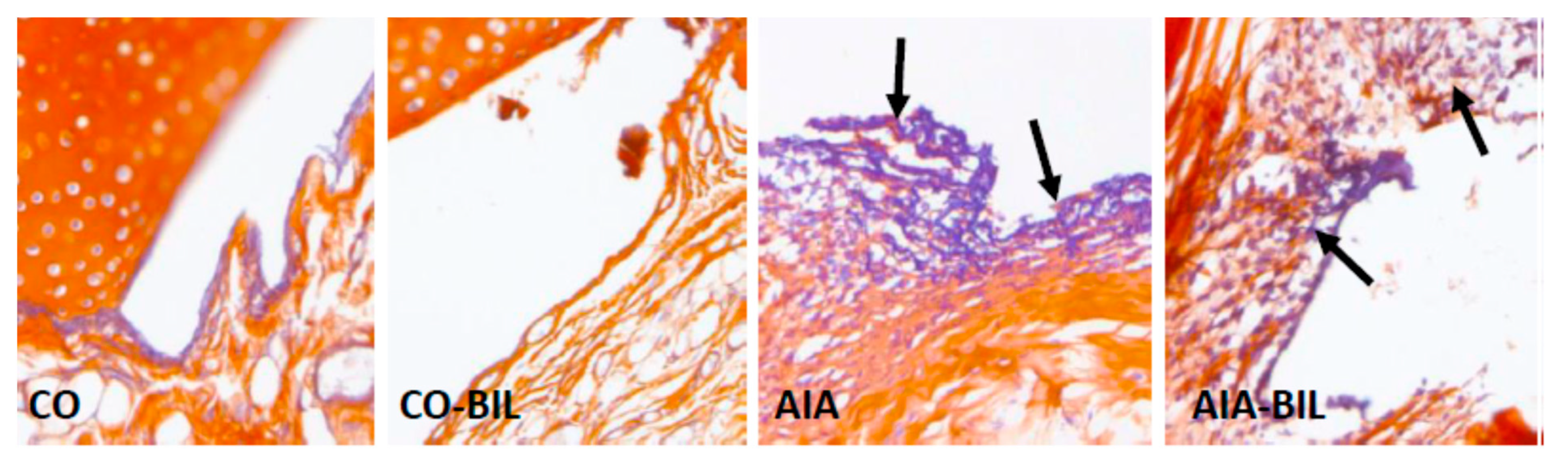
2.5. Oxidative Damage to DNA and Histological Examination
3. Discussion
4. Material and Methods
4.1. Animals and AIA Experimental Model
4.2. The Design of AIA Experiment
4.3. Blood Tests and Oxidative Damage Evaluation
4.4. Histology and Histochemistry
4.5. Morphometry and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Firestein, G.S. Immunologic mechanisms in the pathogenesis of rheumatoid arthritis. J. Clin. Rheumatol. 2005, 11, S39–S44. [Google Scholar] [CrossRef]
- Smolen, J.S.; Landewé, R.B.M.; Bijlsma, J.W.J.; Burmester, G.R.; Dougados, M.; Kerschbaumer, A.; McInnes, I.B.; Sepriano, A.; van Vollenhoven, R.F.; de Wit, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2019 update. Ann. Rheum. Dis. 2020, 79, S685–S699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnes, I.B.; Schett, G. The Pathogenesis of Rheumatoid Arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [Green Version]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Wang, Y.; Xu, D.; Nossent, J.; Pavlos, N.J.; Xu, J. Rheumatoid arthritis: Pathological mechanisms and modern pharmacologic therapies. Bone Res. 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Van Der Heijde, D.; Machold, K.; Aletaha, D.; Landewé, R. Proposal for a new nomenclature of disease-modifying antirheumatic drugs. Ann. Rheum. Dis. 2013, 73, 3–5. [Google Scholar] [CrossRef]
- Cutolo, M.; Sulli, A.; Pizzorni, C.; Seriolo, B.; Straub, R.H. Anti-inflammatory mechanisms of methotrexate in rheumatoid arthritis. Ann. Rheum. Dis. 2001, 60, 729–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazlewood, G.S.; Barnabe, C.; Tomlinson, G.; Marshall, D.; Devoe, D.; Bombardier, C. Methotrexate monotherapy and methotrexate combination therapy with traditional and biologic disease modifying antirheumatic drugs for rheumatoid arthritis: Abridged cochrane Systematic review and network meta-analysis. BMJ 2016, 353, i1777. [Google Scholar] [CrossRef] [Green Version]
- Taams, L.S. Interleukin-17 in rheumatoid arthritis: Trials and tribulations. J. Exp. Med. 2020, 217, 48. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, T.D.; Mikuls, T.R. Recent advances in the treatment of rheumatoid arthritis. Curr. Opin. Rheumatol. 2018, 30, 231–237. [Google Scholar] [CrossRef]
- Salehi, B.; Martorell, M.; Arbiser, J.; Sureda, A.; Martins, N.; Maurya, P.; Sharifi-Rad, M.; Kumar, P.; Sharifi-Rad, J. Antioxidants: Positive or negative actors? Biomolecules 2018, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jangi, S.; Otterbein, L.; Robson, S. The molecular basis for the immunomodulatory activities of unconjugated bilirubin. Int. J. Biochem. Cell Biol. 2013, 45, 2843–2851. [Google Scholar] [CrossRef]
- Sullivan, J.I.; Rockey, D.C. Diagnosis and evaluation of hyperbilirubinemia. Curr. Opin. Gastroenterol. 2017, 33, 164–170. [Google Scholar] [CrossRef]
- Bulmer, A.C.; Verkade, H.J.; Wagner, K.-H. Bilirubin and beyond: A review of lipid status in Gilbert’s syndrome and its relevance to cardiovascular disease protection. Prog. Lipid Res. 2013, 52, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Fischman, D.; Valluri, A.; Gorrepati, V.S.; Murphy, M.E.; Cheriyath, I.P.P. Bilirubin as a Protective Factor for Rheumatoid Arthritis: An NHANES Study of 2003–2006 Data. J. Clin. Med. Res. 2010, 2, 256–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedlak, T.W.; Snyder, S.H. Bilirubin benefits: Cellular protection by a biliverdin reductase antioxidant cycle. Pediatrics 2004, 113, 1776–1782. [Google Scholar] [CrossRef]
- Juping, D.; Yuan, Y.; Shiyong, C.; Jun, L.; Xiuxiu, Z.; Haijian, Y.; Jianfeng, S.; Bo, S. Serum bilirubin and the risk of rheumatoid arthritis. J. Clin. Lab. Anal. 2017, 31, e22118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fevery, J. Bilirubin in clinical practice: A review. Liver Int. 2008, 28, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Tapan, S.; Karadurmus, N.; Dogru, T.; Ercin, C.N.; Tasci, I.; Bilgi, C.; Kurt, I.; Erbil, M.K. Decreased small dense LDL levels in Gilbert’s syndrome. Clin. Biochem. 2011, 44, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Vítek, L.; Jirsa, M., Jr.; Brodanová, M.; Kaláb, M.; Mareček, Z.; Danzig, V.; Novotný, L.; Kotal, P. Gilbert syndrome and ischemic heart disease: A protective effect of elevated bilirubin levels. Atherosclerosis 2002, 160, 449–456. [Google Scholar] [CrossRef]
- Bauerova, K.; Drafi, F.; Kuncirova, V.; Ponist, S.; Mihalova, D.; Babal, P.; Sykora, T. Hyperbilirubinemia decreases physiological markers in adjuvant-induced arthritis. Physiol. Res. 2015, 64, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, P.; Lu, J.; Xiong, W.; Oger, J.; Tetzlaff, W.; Cynader, M. Bilirubin possesses powerful immunomodulatory activity and suppresses experimental autoimmune encephalomyelitis. J. Immunol. 2008, 181, 1887–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauerova, K.; Ponist, S.; Kuncirova, V.; Mihalova, D.; Paulovicova, E.; Volpi, N. Chondroitin sulfate effect on induced arthritis in rats. Osteoarthr. Cartil. 2011, 19, 1373–1379. [Google Scholar] [CrossRef] [Green Version]
- Feketeová, L.; Jančová, P.; Moravcová, P.; Janegová, A.; Bauerová, K.; Poništ, S.; Mihalová, D.; Janega, P.; Babál, P. Effect of methotrexate on inflammatory cells redistribution in experimental adjuvant arthritis. Rheumatol. Int. 2011, 32, 3517–3523. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.M.; Poduval, T.B. Immunomodulatory and immunotoxic effects of bilirubin: Molecular mechanisms. J. Leukoc. Biol. 2011, 90, 997–1015. [Google Scholar] [CrossRef]
- Stanescu, R.; Lider, O.; van Eden, W.; Holoshitz, J.; Cohen, I.R. Histopathology of arthritis induced in rats by active immunization to mycobacterial antigens or by systemic transfer of T lymphocyte lines. A light and electron microscopic study of the articular surface using cationized ferritin. Arthritis Rheum. 1987, 30, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Yonemitsu, Y.; Okano, S.; Nakagawa, K.; Nakashima, Y.; Irisa, T.; Iwamoto, Y.; Nagai, Y.; Hasegawa, M.; Sueishi, K. Arthritis in Rats Adjuvant-induced severity of joint disease in fibroblast growth factor-2 determines. J. Immunol. 2021, 168, 450–457. [Google Scholar] [CrossRef]
- Bonelli, M.; Savitskaya, A.; Steiner, C.W.; Rath, E.; Bilban, M.; Wagner, O.; Bach, F.H.; Smolen, J.S.; Scheinecker, C. Heme oxygenase-1 end-products carbon monoxide and biliverdin ameliorate murine collagen induced arthritis. Clin. Exp. Rheumatol. 2012, 30, 73–78. [Google Scholar]
- Hwang, H.-J.; Lee, S.-W.; Kim, S.-H. Relationship between bilirubin and C-reactive protein. Clin. Chem. Lab. Med. 2011, 49, 1823–1828. [Google Scholar] [CrossRef]
- Kadl, A.; Pontiller, J.; Exner, M.; Leitinger, N. Single bolus injection of bilirubin improves the clinical outcome in a mouse model of endotoxemia. Shock 2007, 28, 582–588. [Google Scholar] [CrossRef]
- Corchia, C.; Balata, A.; Soletta, G.; Mastroni, P.; Meloni, G.F. Increased bilirubin production, ceruloplasmin concentrations and hyperbilirubinaemia in full-term newborn infants. Early Hum. Dev. 1994, 38, 91–96. [Google Scholar] [CrossRef]
- Derelanko, M.J. Determination of erythrocyte life span in F-344, wistar, and sprague-dawley rats using a modification of the [H]diisopropylfluorophosphate ([H]DFP) method. Toxicol. Sci. 1987, 9, 271–276. [Google Scholar] [CrossRef]
- Brito, M.A.; Silva, R.F.M.; Brites, D. Bilirubin toxicity to human erythrocytes: A review. Clin. Chim. Acta 2006, 374, 46–56. [Google Scholar] [CrossRef] [PubMed]
- McDonagh, A.F. Bilirubin toxicity to human erythrocytes: A more sanguine view. Pediatrics 2007, 120, 175–178. [Google Scholar] [CrossRef]
- Jansen, T.; Daiber, A. Direct antioxidant properties of bilirubin and biliverdin. Is there a role for biliverdin reductase? Front. Pharmacol. 2012, 3, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonagh, A.F. The biliverdin–bilirubin antioxidant cycle of cellular protection: Missing a wheel? Free Radic. Biol. Med. 2010, 49, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Bauerova, K.; Acquaviva, A.; Ponist, S.; Gardi, C.; Vecchio, D.; Drafi, F.; Arezzini, B.; Bezakova, L.; Kuncirova, V.; Mihalova, D.; et al. Markers of inflammation and oxidative stress studied in adjuvant-induced arthritis in the rat on systemic and local level affected by pinosylvin and methotrexate and their combination. Autoimmunity 2015, 48, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Wallner, M.; Antl, N.; Rittmannsberger, B.; Schreidl, S.; Najafi, K.; Müllner, E.; Moelzer, C.; Ferk, F.; Knasmüller, S.; Marculescu, R.; et al. Anti-genotoxic potential of bilirubin in vivo: Damage to DNA in hyperbilirubinemic human and animal models. Cancer Prev. Res. 2013, 6, 1056–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.R.; Dobson, V.L.; Dusinská, M.; Kennedy, G.; Stĕtina, R. The comet assay: What can it really tell us. Mutat. Res. 1997, 375, 183–193. [Google Scholar] [CrossRef]
- Liu, J.; Dong, H.; Zhang, Y.; Cao, M.; Song, L.; Pan, Q.; Bulmer, A.C.; Adams, D.B.; Dong, X.; Wang, H. Bilirubin increases insulin sensitivity by regulating cholesterol metabolism, adipokines and PPARγ levels. Sci. Rep. 2015, 5, 9886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, C.; Weeke, P.; Fosbøl, E.; Brendorp, B.; Køber, L.; Coutinho, W.; Sharma, A.M.; Van Gaal, L.; Finer, N.; James, W.P.T.; et al. Acute effect of weight loss on levels of total bilirubin in obese, cardiovascular high-risk patients: An analysis from the lead-in period of the Sibutramine Cardiovascular Outcome trial. Metabolism 2009, 58, 1109–1115. [Google Scholar] [CrossRef]
- García-González, A.; Gaxiola-Robles, R.; Zenteno-Savín, T. Oxidative Stress in Patients with Rheumatoid Arthritis. Rev. Investig. Clin. 2015, 67, 46–53. [Google Scholar]
- Altindag, O. Increased DNA damage and oxidative stress in patients with rheumatoid arthritis. Clin. Biochem. 2007, 40, 167–171. [Google Scholar] [CrossRef]
- Veselinovic, M.; Barudzic, N.; Vuletic, M.; Zivkovic, V.; Tomic-Lucic, A.; Djuric, D.; Jakovljevic, V. Oxidative stress in rheumatoid arthritis patients: Relationship to diseases activity. Mol. Cell. Biochem. 2014, 391, 225–232. [Google Scholar] [CrossRef]
- Bordy, R.; Totoson, P.; Prati, C.; Marie, C.; Wendling, D.; Demougeot, C. Microvascular endothelial dysfunction in rheumatoid arthritis. Nat. Rev. Rheumatol. 2018, 14, 404–420. [Google Scholar] [CrossRef] [PubMed]
- England, B.R.; Thiele, G.M.; Anderson, D.R.; Mikuls, T.R. Increased cardiovascular risk in rheumatoid arthritis: Mechanisms and implications. BMJ 2018, 23, 361. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H.R. ROS as signalling molecules in T cells—Evidence for abnormal redox signalling in the autoimmune disease, rheumatoid arthritis. Redox. Rep. 2005, 10, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Souliotis, V.L.; Vlachogiannis, N.I.; Pappa, M.; Argyriou, A.; Ntouros, P.A.; Sfikakis, P.P. DNA Damage Response and Oxidative Stress in Systemic Autoimmunity. Int. J. Mol. Sci. 2020, 21, 55. [Google Scholar] [CrossRef] [Green Version]
- Bauerová, K.; Poništ, S.; Mihalová, D.; Dráfi, F.; Kuncírová, V. Utilization of adjuvant arthritis model for evaluation of new approaches in rheumatoid arthritis therapy focused on regulation of immune processes and oxidative stress. Interdiscip. Toxicol. 2011, 4, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Bendele, A.; McComb, J.; Gould, T.; McAbee, T.; Sennello, G.; Chlipala, E.; Guy, M. Animal models of arthritis: Relevance to human disease. Toxicol. Pathol. 1999, 27, 134–142. [Google Scholar] [CrossRef]
- Chaudhary, A.; Vinay, P. Rheumatoid Arthritis: Etiology, treatment and animal models. J. Drug Deliv. Ther. 2020, 10, 290–298. [Google Scholar] [CrossRef]
- Collins, A.R.; Dusinská, M.; Gedik, C.M.; Stĕtina, R. Oxidative damage to DNA: Do we have a reliable biomarker? Environ. Health Perspect. 1996, 104 (Suppl. S3), 465–469. [Google Scholar] [CrossRef] [PubMed]
- Pribyl, T. Serum polyphenol oxidase activity (ceruloplasmin) in conventional laboratory animals and man. Folia Biol. 1978, 24, 136–141. [Google Scholar]
- Gedik, C.M.; Collins, A. Establishing the background level of base oxidation in human lymphocyte DNA: Results of an interlaboratory validation study. FASEB J. 2005, 19, 82–84. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | ||||||||
|---|---|---|---|---|---|---|---|---|
| Group | 14 | 21 | 28 | |||||
| Erythrocytes (106/μL) | CO | 0.84 | ±0.04 | 0.96 | ±0.03 | 1.00 | ±0.03 | |
| CO-BIL | 0.94 | ±0.03 | 0.99 | ±0.13 | 0.96 | ±0.02 | ||
| AIA | 0.86 | ±0.02 | 0.96 | ±0.03 | 1.05 | ±0.02 | ||
| AIA-BIL | 0.84 | ±0.03 | 0.91 | ±0.02 | 0.92 ++ | ±0.03 | ||
| Hemoglobin (g/dL) | CO | 1.37 | ±0.05 | 1.63 | ±0.06 | 1.66 | ±0.04 | |
| CO-BIL | 1.69 ** | ±0.07 | 1.63 | ±0.03 | 1.56* | ±0.02 | ||
| AIA | 1.38 | ±0.03 | 1.51 | ±0.04 | 1.60 | ±0.04 | ||
| AIA-BIL | 1.31 | ±0.04 | 1.40 | ±0.04 | 1.42 + | ±0.06 | ||
| MCV (μL) | CO | 47.71 | ±0.84 | 46.86 | ±0.77 | 47.43 | ±0.72 | |
| CO-BIL | 46.63 | ±0.32 | 44.63 * | ±0.26 | 44.71 ** | ±0.29 | ||
| AIA | 45.00 ** | ±0.27 | 43.88 ** | ±0.35 | 42.86 *** | ±0.40 | ||
| AIA-BIL | 43.88 + | ±0.40 | 42.86 | ±0.40 | 42.67 | ±0.33 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sykora, T.; Babal, P.; Mikus-Kuracinova, K.; Drafi, F.; Ponist, S.; Dvorakova, M.; Janega, P.; Bauerova, K. Hyperbilirubinemia Maintained by Chronic Supplementation of Unconjugated Bilirubin Improves the Clinical Course of Experimental Autoimmune Arthritis. Int. J. Mol. Sci. 2021, 22, 8662. https://doi.org/10.3390/ijms22168662
Sykora T, Babal P, Mikus-Kuracinova K, Drafi F, Ponist S, Dvorakova M, Janega P, Bauerova K. Hyperbilirubinemia Maintained by Chronic Supplementation of Unconjugated Bilirubin Improves the Clinical Course of Experimental Autoimmune Arthritis. International Journal of Molecular Sciences. 2021; 22(16):8662. https://doi.org/10.3390/ijms22168662
Chicago/Turabian StyleSykora, Tomas, Pavel Babal, Kristina Mikus-Kuracinova, Frantisek Drafi, Silvester Ponist, Monika Dvorakova, Pavol Janega, and Katarina Bauerova. 2021. "Hyperbilirubinemia Maintained by Chronic Supplementation of Unconjugated Bilirubin Improves the Clinical Course of Experimental Autoimmune Arthritis" International Journal of Molecular Sciences 22, no. 16: 8662. https://doi.org/10.3390/ijms22168662