
Recent Progress in Enhancing Fungal Disease Resistance in Ornamental Plants

 , ,
, ,
Abstract
:1. Introduction
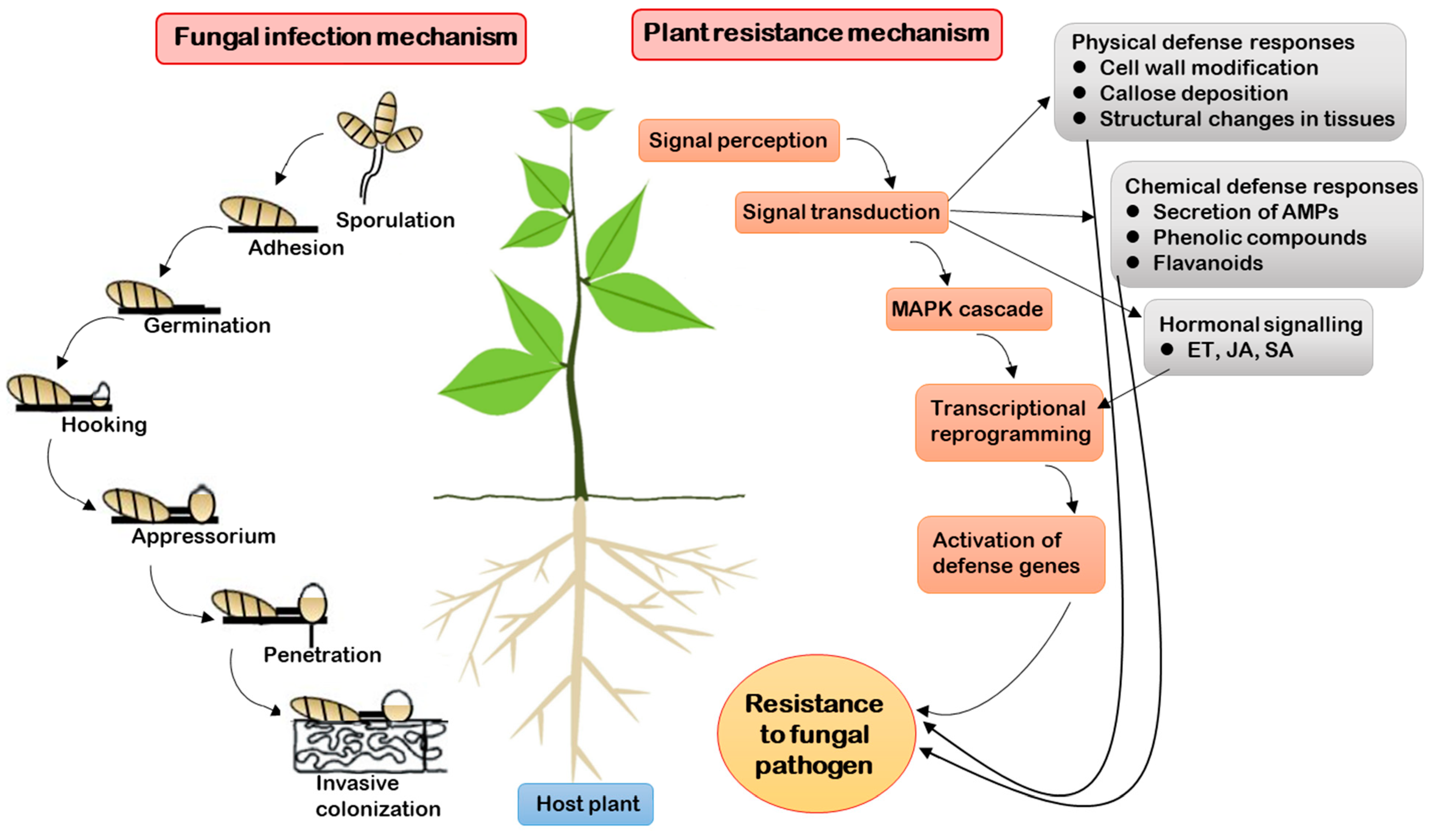
2. General Mechanisms of Fungal Pathogen Infection and Plant Resistance
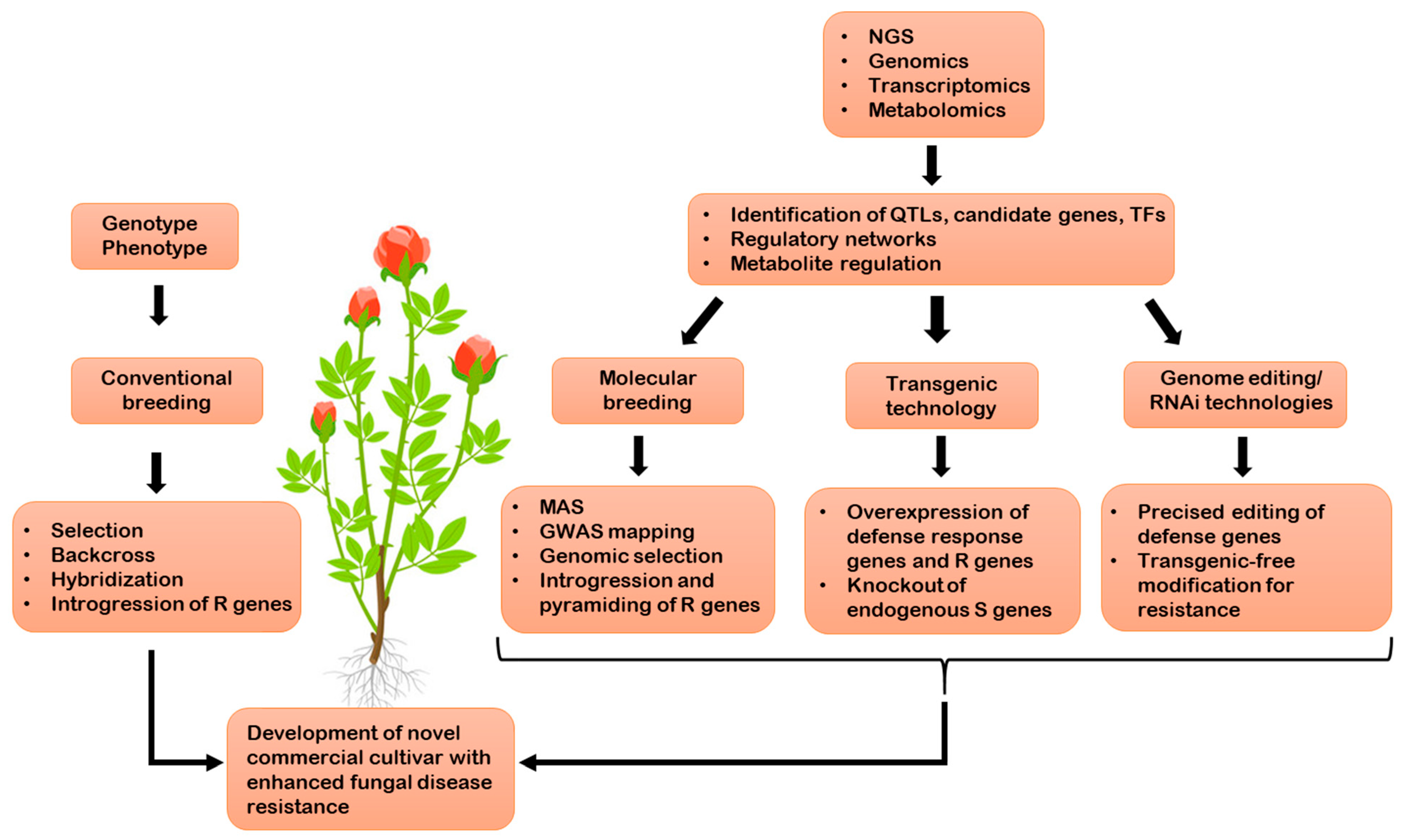
3. Molecular Breeding for Fungal Disease Resistance
4. Genetic Engineering for Improved Fungal Disease Resistance in Ornamental Plants
4.1. Rose
4.2. Chrysanthemum
4.3. Petunia
4.4. Lily
4.5. Other Ornamentals
5. Host-Induced Gene Silencing (HIGS) and Spray-Induced Gene Silencing (SIGS) Used to Control Fungal Pathogens
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lecomte, C.; Alabouvette, C.; Edel-Hermann, V.; Robert, F.; Steinberg, C. Biological control of ornamental plant diseases caused by Fusarium oxysporum: A review. Biol. Cont. 2016, 101, 17–30. [Google Scholar] [CrossRef]
- Chandler, S.F.; Sanchez, C. Genetic modification; the development of transgenic ornamental plant varieties. Plant Biotechnol. J. 2012, 10, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Royal FloraHolland in Facts and Figures. 2019 Annual Report. Available online: https://www.royalfloraholland.com/en (accessed on 15 March 2021).
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Parisi, C.; Tillie, P.; Rodríguez-Cerezo, E. The global pipeline of GM crops out to 2020. Nat. Biotechnol. 2016, 34, 31–36. [Google Scholar] [CrossRef]
- Punja, Z.K. Recent developments toward achieving fungal disease resistance in transgenic plants. Canad. J. Plant Pathol. 2006, 28, S298–S308. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Introductory Mycology; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant pathogenic fungi. In The Fungal Kingdom; John Wiley and Sons: Washington, DC, USA, 2017; pp. 701–726. [Google Scholar]
- Gavrilescu, M.; Chisti, Y. Biotechnology—A sustainable alternative for chemical industry. Biotechnol. Adv. 2005, 23, 471–499. [Google Scholar] [CrossRef]
- Hammond, J.; Hsu, H.-T.; Huang, Q.; Jordan, R.; Kamo, K.; Pooler, M. Transgenic approaches to disease resistance in ornamental crops. J. Crop Imp. 2006, 17, 155–210. [Google Scholar] [CrossRef]
- Jeseničnik, T.; Štajner, N.; Radišek, S.; Jakše, J. RNA interference core components identified and characterised in Verticillium nonalfalfae, a vascular wilt pathogenic plant fungi of hops. Sci. Rep. 2019, 9, 1–12. [Google Scholar]
- Hammond-Kosack, K.E.; Parker, J.E. Deciphering plant–pathogen communication: Fresh perspectives for molecular resistance breeding. Curr. Opin. Biotechnol. 2003, 14, 177–193. [Google Scholar] [CrossRef]
- Michelmore, R.W. The impact zone: Genomics and breeding for durable disease resistance. Curr. Opin. Plant Biol. 2003, 6, 397–404. [Google Scholar] [CrossRef]
- Debener, T. Current strategies and future prospects of resistance breeding in ornamentals. Acta Hortic. 2009, 836, 125–130. [Google Scholar] [CrossRef]
- Arens, P.; Bijman, P.; Tang, N.; Shahin, A.; Van Tuyl, J. Mapping of disease resistance in ornamentals: A long haul. Acta Hortic. 2012, 953, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Parmar, N.; Singh, K.H.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3 Biotech 2017, 7, 1–35. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Mushtaq, M.; Sakina, A.; Wani, S.H.; Shikari, A.B.; Tripathi, P.; Zaid, A.; Galla, A.; Abdelrahman, M.; Sharma, M.; Singh, A.K. Harnessing genome editing techniques to engineer disease resistance in plants. Front. Plant Sci. 2019, 10, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, W. Molecular mechanisms of fungal pathogenicity to plants. Ann. Rev. Phytopathol. 1994, 32, 461–477. [Google Scholar] [CrossRef]
- Nicholson, R.L.; Epstein, L. Adhesion of fungi to the plant surface. In The Fungal Spore and Disease Initiation in Plants and Animals; Plenum Press: New York, NY, USA, 1991; pp. 3–23. [Google Scholar]
- Hamer, J.E.; Howard, R.J.; Chumley, F.G.; Valent, B. A mechanism for surface attachment in spores of a plant pathogenic fungus. Science 1988, 239, 288–290. [Google Scholar] [CrossRef]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Uma, B.; Rani, T.S.; Podile, A.R. Warriors at the gate that never sleep: Non-host resistance in plants. J. Plant Physiol. 2011, 168, 2141–2152. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, O.X.; Ronald, P.C. Genetic engineering for disease resistance in plants: Recent progress and future perspectives. Plant Physiol. 2019, 180, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Thomma, B.P.; Cammue, B.P.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Kachroo, A.; Vincelli, P.; Kachroo, P. Signaling mechanisms underlying resistance responses: What have we learned, and how is it being applied? Phytopathology 2017, 107, 1452–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutrot, F.; Zipfel, C. Function, discovery, and exploitation of plant pattern recognition receptors for broad-spectrum disease resistance. Ann. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Kumar, R.; Kumar, V.; Won, S.Y.; Shukla, P. Engineering disease resistant plants through CRISPR-Cas9 technology. GM Crops Food 2021, 12, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Hamel, L.-P.; Nicole, M.-C.; Duplessis, S.; Ellis, B.E. Mitogen-activated protein kinase signaling in plant-interacting fungi: Distinct messages from conserved messengers. Plant Cell 2012, 24, 1327–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.S.; Arraes, F.B.M.; de Araújo Campos, M.; Grossi-de-Sa, M.; Fernandez, D.; de Souza Cândido, E.; Cardoso, M.H.; Franco, O.L.; Grossi-de-Sa, M.F. Potential biotechnological assets related to plant immunity modulation applicable in engineering disease-resistant crops. Plant Sci. 2018, 270, 72–84. [Google Scholar] [CrossRef]
- González, G.; Fuentes, L.; Moya-León, M.A.; Sandoval, C.; Herrera, R. Characterization of two PR genes from Fragaria chiloensis in response to Botrytis cinerea infection: A comparison with Fragaria x ananassa. Physiol. Mol. Plant Pathol. 2013, 82, 73–80. [Google Scholar] [CrossRef]
- Silvia Sebastiani, M.; Bagnaresi, P.; Sestili, S.; Biselli, C.; Zechini, A.; Orrù, L.; Cattivelli, L.; Ficcadenti, N. Transcriptome analysis of the melon-Fusarium oxysporum f. sp. melonis race 1.2 pathosystem in susceptible and resistant plants. Front. Plant Sci. 2017, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Derksen, H.; Rampitsch, C.; Daayf, F. Signaling cross-talk in plant disease resistance. Plant Sci. 2013, 207, 79–87. [Google Scholar] [CrossRef]
- Won, S.Y.; Kim, J.S.; Kang, S.-H.; Sohn, S.-H. Current status and prospects of chrysanthemum genomics. J. Plant Biotechnol. 2016, 43, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Debener, T. Molecular tools for modern ornamental plant breeding and selection. Acta Hortic. 2001, 552, 121–128. [Google Scholar] [CrossRef]
- Debener, T.; Byrne, D.H. Disease resistance breeding in rose: Current status and potential of biotechnological tools. Plant Sci. 2014, 228, 107–117. [Google Scholar] [CrossRef]
- Byrne, D.H. Rose structural genomics. In Genetics and Genomics of Rosaceae; Springer: New York, NY, USA, 2009; pp. 353–379. [Google Scholar]
- Gar, O.; Sargent, D.J.; Tsai, C.-J.; Pleban, T.; Shalev, G.; Byrne, D.H.; Zamir, D. An autotetraploid linkage map of rose (Rosa hybrida) validated using the strawberry (Fragaria vesca) genome sequence. PLoS ONE 2011, 6, e20463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiller, M.; Linde, M.; Hibrand-Saint Oyant, L.; Tsai, C.-J.; Byrne, D.H.; Smulders, M.J.; Foucher, F.; Debener, T. Towards a unified genetic map for diploid roses. Theor. App. Genet. 2011, 122, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Malek, B.; Debener, T. Genetic analysis of resistance to blackspot (Diplocarpon rosae) in tetraploid roses. Theor. App. Genet. 1998, 96, 228–231. [Google Scholar] [CrossRef]
- Linde, M.; Mattiesch, L.; Debener, T. Rpp1, a dominant gene providing race-specific resistance to rose powdery mildew (Podosphaera pannosa): Molecular mapping, SCAR development and confirmation of disease resistance data. Theor. Appl. Genet. 2004, 109, 1261–1266. [Google Scholar] [CrossRef]
- Linde, M.; Hattendorf, A.; Kaufmann, H.; Debener, T. Powdery mildew resistance in roses: QTL mapping in different environments using selective genotyping. Theor. App. Genet. 2006, 113, 1081–1092. [Google Scholar] [CrossRef]
- Hosseini Moghaddam, H.; Leus, L.; Van Huylenbroeck, J.; Van Bockstaele, E.; De Riek, J. Pathotype dependent resistance mapping for powdery mildew in a diploid rose population. Acta Hortic. 2010, 870, 103–108. [Google Scholar] [CrossRef]
- Moghaddam, H.H.; Leus, L.; De Riek, J.; Van Huylenbroeck, J.; Van Bockstaele, E. Construction of a genetic linkage map with SSR, AFLP and morphological markers to locate QTLs controlling pathotype-specific powdery mildew resistance in diploid roses. Euphytica 2012, 184, 413–427. [Google Scholar] [CrossRef]
- Xu, Q.; Wen, X.; Deng, X. Isolation of TIR and nonTIR NBS–LRR resistance gene analogues and identification of molecular markers linked to a powdery mildew resistance locus in chestnut rose (Rosa roxburghii Tratt). Theor. App. Genet. 2005, 111, 819–830. [Google Scholar] [CrossRef]
- Xu, Q.; Wen, X.; Deng, X. Cloning of two classes of PR genes and the development of SNAP markers for powdery mildew resistance loci in chestnut rose (Rosa roxburghii Tratt). Mol. Breed. 2007, 19, 179–191. [Google Scholar] [CrossRef]
- Terefe-Ayana, D.; Yasmin, A.; Le, T.L.; Kaufmann, H.; Biber, A.; Kühr, A.; Linde, M.; Debener, T. Mining disease-resistance genes in roses: Functional and molecular characterization of the Rdr1 locus. Front. Plant Sci. 2011, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, H.; Qiu, X.; Wehmeyer, J.; Debener, T. Isolation, molecular characterization, and mapping of four rose MLO orthologs. Front. Plant Sci. 2012, 3, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Jiang, J.; Zhang, F.; Liu, Y.; Ding, L.; Chen, S.; Chen, F. Current achievements and future prospects in the genetic breeding of chrysanthemum: A review. Hortic. Res. 2019, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K.; Arens, P.; Lim, J.H.; Jung, J.A.; Shin, H.K. Inheritance Analysis and Marker Development of White Rust Resistance in Chrysanthemum. Kor. Hortic. Soc. Acad. Pres. 2017, 10, 47. (In Korean) [Google Scholar]
- Hirakawa, H.; Sumitomo, K.; Hisamatsu, T.; Nagano, S.; Shirasawa, K.; Higuchi, Y.; Kusaba, M.; Koshioka, M.; Nakano, Y.; Yagi, M. De novo whole-genome assembly in Chrysanthemum seticuspe, a model species of Chrysanthemums, and its application to genetic and gene discovery analysis. DNA Res. 2019, 26, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Heusden, A.W.; Jongerius, M.C.; Van Tuyl, J.M.; Straathof, T.P.; Mes, J. Molecular assisted breeding for disease resistance in lily. In XX International Eucarpia Symposium, Section Ornamentals, Strategies for New Ornamentals-Part II 572; ISHS: Leuven, Belgium, 2001; pp. 131–138. [Google Scholar]
- Shahin, A.; Arens, P.; Van Heusden, S.; Van Tuyl, J.M. Conversion of molecular markers linked to Fusarium and virus resistance in Asiatic lily hybrids. Acta Hortic. 2009, 836, 131–136. [Google Scholar] [CrossRef]
- Shahin, A.; Arens, P.; Van Heusden, A.W.; Van Der Linden, G.; Van Kaauwen, M.; Khan, N.; Schouten, H.J.; Van De Weg, W.E.; Visser, R.G.; Van Tuyl, J.M. Genetic mapping in Lilium: Mapping of major genes and quantitative trait loci for several ornamental traits and disease resistances. Plant Breed. 2011, 130, 372–382. [Google Scholar] [CrossRef]
- Moreno-Pachon, N.M.; Leeggangers, H.A.; Nijveen, H.; Severing, E.; Hilhorst, H.; Immink, R.G. Elucidating and mining the Tulipa and Lilium transcriptomes. Plant Mol. Biol. 2016, 92, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, N.; van der Lee, T.; Shahin, A.; Holdinga, M.; Bijman, P.; Caser, M.; Visser, R.G.; van Tuyl, J.M.; Arens, P. Genetic mapping of resistance to Fusarium oxysporum f. sp. tulipae in tulip. Mol. Breed. 2015, 35, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Esselink, G.D.; Visser, R.G.; van Tuyl, J.M.; Arens, P. Transcriptome analysis of Gerbera hybrida including in silico confirmation of defense genes found. Front. Plant Sci. 2016, 7, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Van Silfhout, A.; Shahin, A.; Egberts, R.; Beers, M.; Van der Velde, A.; Van Houten, A.; Van Tuyl, J.M.; Visser, R.G.; Arens, P. Genetic mapping and QTL analysis of Botrytis resistance in Gerbera hybrida. Mol. Breed. 2017, 37, 13. [Google Scholar] [CrossRef] [Green Version]
- Tanase, K.; Nishitani, C.; Hirakawa, H.; Isobe, S.; Tabata, S.; Ohmiya, A.; Onozaki, T. Transcriptome analysis of carnation (Dianthus caryophyllus L.) based on next-generation sequencing technology. BMC Genom. 2012, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Yamamoto, T.; Isobe, S.; Hirakawa, H.; Tabata, S.; Tanase, K.; Yamaguchi, H.; Onozaki, T. Construction of a reference genetic linkage map for carnation (Dianthus caryophyllus L.). BMC Genom. 2013, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Kosugi, S.; Hirakawa, H.; Ohmiya, A.; Tanase, K.; Harada, T.; Kishimoto, K.; Nakayama, M.; Ichimura, K.; Onozaki, T. Sequence analysis of the genome of carnation (Dianthus caryophyllus L.). DNA Res. 2014, 21, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Manczinger, L.; Antal, Z.; Kredics, L. Ecophysiology and breeding of mycoparasitic Trichoderma strains. Acta Microbiol. Immunol. Hung. 2002, 49, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H. Inducing fungus-resistance into plants through biotechnology. Notul. Sci. Biol. 2010, 2, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Cairns, T. Modern Roses XII; Academic Press: New York, NY, USA, 2007. [Google Scholar]
- Feng, L.-G.; Chen, C.; Sheng, L.-X.; Liu, P.; Tao, J.; Su, J.-L.; Zhao, L.-Y. Comparative analysis of headspace volatiles of Chinese Rosa rugosa. Molecules 2010, 15, 8390–8399. [Google Scholar] [CrossRef]
- Dohm, A.; Ludwig, C.; Schilling, D.; Debener, T. Transformation of roses with genes for antifungal proteins to reduce their susceptibility to fungal diseases. Acta Hortic. 2002, 572, 105–111. [Google Scholar] [CrossRef]
- Firoozabady, E.; Moy, Y.; Courtney-Gutterson, N.; Robinson, K. Regeneration of transgenic rose (Rosa hybrida) plants from embryogenic tissue. Bio/technology 1994, 12, 609–613. [Google Scholar] [CrossRef]
- Linde, M.; Shishkoff, N. DISEASE|Powdery Mildew. In Encyclopedia of Rose Science; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA; Oxford, UK, 2003; pp. 158–165. Available online: https://www.elsevier.com/books/encyclopedia-of-rose-science/roberts/978-0-08-091797-9 (accessed on 11 March 2021).
- Terras, F.; Schoofs, H.; De Bolle, M.; Van Leuven, F.; Rees, S.B.; Vanderleyden, J.; Cammue, B.; Broekaert, W.F. Analysis of two novel classes of plant antifungal proteins from radish (Raphanus sativus L.) seeds. J. Biol. Chem. 1992, 267, 15301–15309. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Terras, F.; Cammue, B.; Osborn, R.W. Plant defensins: Novel antimicrobial peptides as components of the host defense system. Plant Physiol. 1995, 108, 1353. [Google Scholar] [CrossRef] [PubMed]
- Morais, J.K.S.; Gomes, V.M.; Oliveira, J.T.A.; Santos, I.S.; Da Cunha, M.; Oliveira, H.D.; Oliveira, H.P.; Sousa, D.O.; Vasconcelos, I.M. Soybean toxin (SBTX), a protein from soybeans that inhibits the life cycle of plant and human pathogenic fungi. J. Agric. Food Chem. 2010, 58, 10356–10363. [Google Scholar] [CrossRef]
- Thevissen, K.; Ghazi, A.; De Samblanx, G.W.; Brownlee, C.; Osborn, R.W.; Broekaert, W.F. Fungal membrane responses induced by plant defensins and thionins. J. Biol. Chem. 1996, 271, 15018–15025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Gasic, K.; Cammue, B.; Broekaert, W.; Korban, S.S. Transgenic rose lines harboring an antimicrobial protein gene, Ace-AMP1, demonstrate enhanced resistance to powdery mildew (Sphaerotheca pannosa). Planta 2003, 218, 226–232. [Google Scholar] [CrossRef]
- Pourhosseini, L.; Kermani, M.J.; Habashi, A.A.; Khalighi, A. Efficiency of direct and indirect shoot organogenesis in different genotypes of Rosa hybrida. Plant Cell Tissue Organ Cult. 2013, 112, 101–108. [Google Scholar] [CrossRef]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Gen. 2006, 38, 716–720. [Google Scholar] [CrossRef]
- Humphry, M.; Reinstaedler, A.; Ivanov, S.; Bisseling, T.; Panstruga, R. Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsMLO1. Mol. Plant Pathol. 2011, 12, 866–878. [Google Scholar] [CrossRef]
- Bhat, R.A.; Miklis, M.; Schmelzer, E.; Schulze-Lefert, P.; Panstruga, R. Recruitment and interaction dynamics of plant penetration resistance components in a plasma membrane microdomain. Proc. Nat. Acad. Sci. USA 2005, 102, 3135–3140. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Wang, Q.; Zhang, H.; Jian, H.; Zhou, N.; Ji, C.; Yan, H.; Bao, M.; Tang, K. Antisense RhMLO1 gene transformation enhances resistance to the powdery mildew pathogen in Rosa multiflora. Plant Mol. Biol. Rep. 2015, 33, 1659–1665. [Google Scholar] [CrossRef]
- Xiang, G.; Zhang, H.; Jian, H.; Yan, H.; Wang, Q.; Zhou, N.; Li, S.; Tang, K.; Qiu, X. De Novo assembly and characterization of the transcriptome of susceptible and resistant rose species in response to powdery mildew. Sci. Hortic. 2019, 257, 108653. [Google Scholar] [CrossRef]
- Drewes-Alvarez, R. Disease/Black Spot. In Encyclopedia of Rose Science; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA; Oxford, UK, 2003; pp. 148–153. Available online: https://www.elsevier.com/books/encyclopedia-of-rose-science/roberts/978-0-08-091797-9 (accessed on 11 March 2021).
- Marchant, R.; Davey, M.R.; Lucas, J.A.; Lamb, C.J.; Dixon, R.A.; Power, J.B. Expression of a chitinase transgene in rose (Rosa hybrida L.) reduces development of blackspot disease (Diplocarpon rosae Wolf). Mol. Breed. 1998, 4, 187–194. [Google Scholar] [CrossRef]
- Neu, E.; Domes, H.S.; Menz, I.; Kaufmann, H.; Linde, M.; Debener, T. Interaction of roses with a biotrophic and a hemibiotrophic leaf pathogen leads to differences in defense transcriptome activation. Plant Mol. Biol. 2019, 99, 299–316. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Cao, X.; Ma, C.; Zhang, Z.; Zhao, N.; Ali, A.; Hou, T.; Xiang, Z.; Zhuang, J.; Wu, S. Potential applications and antifungal activities of engineered nanomaterials against gray mold disease agent Botrytis cinerea on rose petals. Front. Plant Sci. 2017, 8, 1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, B.; Duncan, G.H.; Harrison, J.G.; Harding, L.A.; Elad, Y.; Zimand, G. Effect of humidity on infection of rose petals by dry-inoculated conidia of Botrytis cinerea. Mycol. Res. 1995, 99, 1303–1310. [Google Scholar] [CrossRef]
- Cao, X.; Yan, H.; Liu, X.; Li, D.; Sui, M.; Wu, J.; Yu, H.; Zhang, Z. A detached petal disc assay and virus-induced gene silencing facilitate the study of Botrytis cinerea resistance in rose flowers. Hortic. Res. 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Liu, X.; Shu, L.; Zhang, H.; Zhang, S.; Song, Y.; Zhang, Z. Global analysis of the AP2/ERF gene family in rose (Rosa chinensis) genome unveils the role of RcERF099 in Botrytis resistance. BMC Plant Biol. 2020, 20, 1–15. [Google Scholar] [CrossRef]
- Mekapogu, M.; Kwon, O.K.; Lee, K.J.; Ahn, M.S.; Park, J.T.; Jung, J.A. Identification of standard type cultivars in Chrysanthemum (Dendranthema grandiflorum) using SSR markers. Hortic. Environ. Biotechnol. 2020, 61, 153–161. [Google Scholar] [CrossRef]
- Mekapogu, M.; Vasamsetti, B.M.K.; Kwon, O.-K.; Ahn, M.-S.; Lim, S.-H.; Jung, J.-A. Anthocyanins in Floral Colors: Biosynthesis and Regulation in Chrysanthemum Flowers. Int. J. Mol. Sci. 2020, 21, 6537. [Google Scholar] [CrossRef] [PubMed]
- Dowrick, G.; El-Bayoumi, A. The origin of new forms of the garden Chrysanthemum. Euphytica 1966, 15, 32–38. [Google Scholar]
- Fukai, S.; de Jong, J.; Rademaker, W. Efficient genetic transformation of chrysanthemum (Dendranthema grandiflorum (Ramat.) Kitamura) using stem segments. Jpn. J. Breed. 1995, 45, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Liu, Z.; Chen, S.; Chen, F. Expression of P. mume PGIP gene in transgenic Dendranthema morifolium increased tolerance to disease resistance. Acta Bot. Boreal. Occident. Sin. 2010, 30, 1111–1116. [Google Scholar]
- Xu, G.; Chen, S.; Chen, F. Transgenic chrysanthemum plants expressing a harpin Xoo gene demonstrate induced resistance to Alternaria leaf spot and accelerated development. Russ. J. Plant Physiol. 2010, 57, 548–553. [Google Scholar] [CrossRef]
- Sen, S.; Kumar, S.; Ghani, M.; Thakur, M. Agrobacterium mediated genetic transformation of chrysanthemum (Dendranthema grandiflora Tzvelev) with rice chitinase gene for improved resistance against Septoria obesa. Plant Pathol. J. 2013, 12, 1–10. [Google Scholar] [CrossRef]
- Takatsu, Y.; Nishizawa, Y.; Hibi, T.; Akutsu, K. Transgenic chrysanthemum (Dendranthema grandiflorum (Ramat.) Kitamura) expressing a rice chitinase gene shows enhanced resistance to gray mold (Botrytis cinerea). Sci. Hortic. 1999, 82, 113–123. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Lim, S.; Yoda, H.; Choi, Y.-E.; Sano, H. Simultaneous activation of salicylate production and fungal resistance in transgenic chrysanthemum producing caffeine. Plant Signal. Behav. 2011, 6, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Choi, Y.-E.; Sano, H. Plant vaccination: Stimulation of defense system by caffeine production in planta. Plant Signal. Behav. 2010, 5, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Kim, C.H. Effect of temperature and pH on sporidia formation of Puccinia horiana on chrysanthemum and evaluation of varietal resistance. Plant Pathol. J. 1993, 9, 42–46. [Google Scholar]
- Dickens, J. Infection of chrysanthemum flowers by white rust (Puccinia horiana Henn.). Plant Pathol. 1970, 19, 122–124. [Google Scholar] [CrossRef]
- Ichikawa, H.; Kato, K.; Mochizuki, A.; Shinoyama, H.; Mitsuhara, I. Transgenic chrysanthemums (Chrysanthemum morifolium Ramat.) carrying both insect and disease resistance. Acta Hortic. 2015, 1087, 485–497. [Google Scholar]
- Bi, M.; Li, X.; Yan, X.; Liu, D.; Gao, G.; Zhu, P.; Mao, H. Chrysanthemum WRKY15-1 promotes resistance to Puccinia horiana Henn. via the salicylic acid signaling pathway. Hortic. Res. 2021, 8, 1–11. [Google Scholar] [CrossRef]
- Bombarely, A.; Moser, M.; Amrad, A.; Bao, M.; Bapaume, L.; Barry, C.S.; Bliek, M.; Boersma, M.R.; Borghi, L.; Bruggmann, R. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vandenbussche, M.; Chambrier, P.; Rodrigues Bento, S.; Morel, P. Petunia, your next supermodel? Front. Plant Sci. 2016, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L.; Jankovics, T.; Kovács, G.M.; Daughtrey, M.L. Oidium longipes, a new powdery mildew fungus on petunia in the USA: A potential threat to ornamental and vegetable solanaceous crops. Plant Dis. 2008, 92, 818–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, A.B.; Kobayashi, D.Y.; Bergen, M.S. Identification of bacteria for biological control of Botrytis cinerea on petunia using a petal disk assay. Plant Dis. 1996, 80, 1029–1033. [Google Scholar] [CrossRef]
- Esposito, S.; Colucci, M.; Frusciante, L.; Filippone, E.; Lorito, M.; Bressan, R. Antifungal transgenes expression in Petunia hybrida. Acta Hortic. 2000, 508, 157–162. [Google Scholar] [CrossRef]
- Khan, R.S.; Alam, S.S.; Munir, I.; Azadi, P.; Nakamura, I.; Mii, M. Botrytis cinerea-resistant marker-free Petunia hybrida produced using the MAT vector system. Plant Cell Tissue Organ Cult. 2011, 106, 11–20. [Google Scholar] [CrossRef]
- Khan, R.S.; Kameya, N.; Mii, M.; Nakamura, I. Transgenic Petunia hybrida expressing a synthetic fungal chitinase gene confers disease tolerance to Botrytis cinerea. Plant Biotechnol. 2012, 29, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Chen, Y.; Wilde, H.D. Reduction of MLO1 expression in petunia increases resistance to powdery mildew. Sci. Hortic. 2016, 201, 225–229. [Google Scholar] [CrossRef]
- Hou, P.-F.; Chen, C.-Y. Early stages of infection of lily leaves by Botrytis elliptica and B. cinerea. Plant Pathol. Bull. 2003, 12, 103–108. [Google Scholar]
- De Cáceres González, F.F.N.; Davey, M.R.; Sanchez, E.C.; Wilson, Z.A. Conferred resistance to Botrytis cinerea in Lilium by overexpression of the RCH10 chitinase gene. Plant Cell Rep. 2015, 34, 1201–1209. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, Q.; Zhao, Y.Q.; Yang, J.; He, H.B.; Jia, G.X. The lre-miR159a-LrGAMYB pathway mediates resistance to grey mould infection in Lilium regale. Mol. Plant Pathol. 2020, 21, 749–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.H.; Pan, Y.C.; Ye, N.H.; Shih, Y.T.; Liu, F.W.; Chen, C.Y. LsGRP1, a class II glycine-rich protein of Lilium, confers plant resistance via mediating innate immune activation and inducing fungal programmed cell death. Mol. Plant Pathol. 2020, 21, 1149–1166. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, X.; Li, S.; Jiang, C.-Z.; Zhang, Y.; Niu, L. LrABCF1, a GCN-type ATP-binding cassette transporter from Lilium regale, is involved in defense responses against viral and fungal pathogens. Planta 2016, 244, 1185–1199. [Google Scholar] [CrossRef]
- Han, Q.; Chen, R.; Yang, Y.; Cui, X.; Ge, F.; Chen, C.; Liu, D. A glutathione S-transferase gene from Lilium regale Wilson confers transgenic tobacco resistance to Fusarium oxysporum. Sci. Hortic. 2016, 198, 370–378. [Google Scholar] [CrossRef]
- Li, H.; Liu, D.; He, H.; Zhang, N.; Ge, F.; Chen, C. Molecular cloning of a 14-3-3 protein gene from Lilium regale Wilson and overexpression of this gene in tobacco increased resistance to pathogenic fungi. Sci. Hortic. 2014, 168, 9–16. [Google Scholar] [CrossRef]
- Rao, J.; Liu, D.; Zhang, N.; He, H.; Ge, F.; Chen, C. Differential gene expression in incompatible interaction between Lilium regale Wilson and Fusarium oxysporum f. sp. lilii revealed by combined SSH and microarray analysis. Mol. Biol. 2014, 48, 802–812. [Google Scholar] [CrossRef]
- He, H.; Liu, D.; Zhang, N.; Zheng, W.; Han, Q.; Ji, B.; Ge, F.; Chen, C. The PR10 gene family is highly expressed in Lilium regale Wilson during Fusarium oxysporum f. sp. lilii infection. Gen. Genom. 2014, 36, 497–507. [Google Scholar] [CrossRef]
- Zhang, N.; Guan, R.; Yang, Y.; Bai, Z.; Ge, F.; Liu, D. Isolation and characterization of a Fusarium oxysporum-resistant gene LrGLP1 from Lilium regale Wilson. Vitr. Cell. Dev. Biol. Plant 2017, 53, 461–468. [Google Scholar] [CrossRef]
- Zuker, A.; Shklarman, E.; Scovel, G.; Ben-Meir, H.; Ovadis, M.; Neta-Sharir, I.; Ben-Yephet, Y.; Weiss, D.; Watad, A.; Vainstein, A. Genetic Engineering of Agronomic and Ornamental Traits in Carnation. Acta Hortic. 2001, 560, 91–94. [Google Scholar] [CrossRef]
- Brugliera, F.; Kalc-Wright, G.; Hyland, C.; Webb, L.; Herbert, S.; Sheehan, B.; Mason, J. Improvement of Fusarium wilt tolerance in carnations expressing chitinase. Int. Plant Mol. Biol. Rep. 2000, 18, 522–529. [Google Scholar]
- Ahn, B.; Shin, H.; Hwang, K.; Min, B.; Joung, H. Transformation of carnations with jasmonate methyl transferase gene for fusarium tolerance. In Vitro Cell. Dev. Biol. 2004, 40, 45A. [Google Scholar]
- Kamo, K.; Lakshman, D.; Bauchan, G.; Rajasekaran, K.; Cary, J.; Jaynes, J. Expression of a synthetic antimicrobial peptide, D4E1, in Gladiolus plants for resistance to Fusarium oxysporum f. sp. gladioli. Plant Cell, Tiss. Org. Cul. 2015, 121, 459–467. [Google Scholar] [CrossRef]
- Kamo, K.; Lakshman, D.; Pandey, R.; Guaragna, M.A.; Okubara, P.; Rajasekaran, K.; Cary, J.; Jordan, R. Resistance to Fusarium oxysporum f. sp. gladioli in transgenic Gladiolus plants expressing either a bacterial chloroperoxidase or fungal chitinase genes. Plant Cell Tissuse Organ Cult. 2016, 124, 541–553. [Google Scholar] [CrossRef]
- Bi, Y.-M.; Cammue, B.; Goodwin, P.; KrishnaRaj, S.; Saxena, P. Resistance to Botrytis cinerea in scented geranium transformed with a gene encoding the antimicrobial protein Ace-AMP1. Plant Cell Rep. 1999, 18, 835–840. [Google Scholar] [CrossRef]
- Narendra Ram, M.; Mohandas, S. Transformation of african violet (Saintpaulia ionantha) with glucanasechitinase genes using Agrobacterium tumefaciens. Acta Hortic. 2003, 624, 471–478. [Google Scholar] [CrossRef]
- Wang, Y.; Browning, M.; Ruemmele, B.A.; Chandlee, J.M.; Kausch, A.P.; Jackson, N. Glufosinate reduces fungal diseases in transgenic glufosinate-resistant bentgrasses (Agrostis spp). Weed Sci. 2003, 51, 130–137. [Google Scholar] [CrossRef]
- Bhattarai, K.; Conesa, A.; Xiao, S.; Peres, N.A.; Clark, D.G.; Parajuli, S.; Deng, Z. Sequencing and analysis of gerbera daisy leaf transcriptomes reveal disease resistance and susceptibility genes differentially expressed and associated with powdery mildew resistance. BMC Plant Biol. 2020, 20, 1–17. [Google Scholar] [CrossRef]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.-H. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase–encoding genes confers strong resistance to Fusarium species. Proc. Nat. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Jin, H. Spray-induced gene silencing: A powerful innovative strategy for crop protection. Trends Microbiol. 2017, 25, 4–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, H.; Kim, J.-I. Advanced strategies to control plant pathogenic fungi by host-induced gene silencing (HIGS) and spray-induced gene silencing (SIGS). Plant Biotechnol. Rep. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Andrade, C.; Tinoco, M.; Rieth, A.; Maia, F.; Aragão, F. Host-induced gene silencing in the necrotrophic fungal pathogen Sclerotinia sclerotiorum. Plant Pathol. 2016, 65, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Dou, T.; Shao, X.; Hu, C.; Liu, S.; Sheng, O.; Bi, F.; Deng, G.; Ding, L.; Li, C.; Dong, T. Host-induced gene silencing of Foc TR4 ERG6/11 genes exhibits superior resistance to Fusarium wilt of banana. Plant Biotechnol. J. 2020, 18, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Thomma, B.P. Host-induced gene silencing compromises Verticillium wilt in tomato and Arabidopsis. Mol. Plant Pathol. 2018, 19, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Jahan, S.N.; Åsman, A.K.; Corcoran, P.; Fogelqvist, J.; Vetukuri, R.R.; Dixelius, C. Plant-mediated gene silencing restricts growth of the potato late blight pathogen Phytophthora infestans. J. Exp. Bot. 2015, 66, 2785–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pliego, C.; Nowara, D.; Bonciani, G.; Gheorghe, D.M.; Xu, R.; Surana, P.; Whigham, E.; Nettleton, D.; Bogdanove, A.J.; Wise, R.P. Host-induced gene silencing in barley powdery mildew reveals a class of ribonuclease-like effectors. Mol. Plant-Microbe Int. 2013, 26, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Song, X.S.; Li, H.P.; Cao, L.H.; Sun, K.; Qiu, X.L.; Xu, Y.B.; Yang, P.; Huang, T.; Zhang, J.B. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to F usarium head blight and seedling blight in wheat. Plant Biotechnol. J. 2015, 13, 1335–1345. [Google Scholar] [CrossRef]
- Chen, W.; Kastner, C.; Nowara, D.; Oliveira-Garcia, E.; Rutten, T.; Zhao, Y.; Deising, H.B.; Kumlehn, J.; Schweizer, P. Host-induced silencing of Fusarium culmorum genes protects wheat from infection. J. Exp. Bot. 2016, 67, 4979–4991. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhu, J.; Liu, Z.; Wang, Z.; Zhou, C.; Wang, H. Host-induced gene silencing of rice blast fungus Magnaporthe oryzae pathogenicity genes mediated by the brome mosaic virus. Genes 2017, 8, 241. [Google Scholar] [CrossRef]
- Zhu, X.; Qi, T.; Yang, Q.; He, F.; Tan, C.; Ma, W.; Voegele, R.T.; Kang, Z.; Guo, J. Host-induced gene silencing of the MAPKK gene PsFUZ7 confers stable resistance to wheat stripe rust. Plant Physiol. 2017, 175, 1853–1863. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Zhu, X.; Tan, C.; Liu, P.; Guo, J.; Kang, Z.; Guo, J. Host-induced gene silencing of an important pathogenicity factor P s CPK 1 in Puccinia striiformis f. sp. tritici enhances resistance of wheat to stripe rust. Plant Biotechnol. J. 2018, 16, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Panwar, V.; Jordan, M.; McCallum, B.; Bakkeren, G. Host-induced silencing of essential genes in Puccinia triticina through transgenic expression of RNA i sequences reduces severity of leaf rust infection in wheat. Plant Biotechnol. J. 2018, 16, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.-M.; Thomma, B.P.; Huang, H.-D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef]
- McLoughlin, A.G.; Wytinck, N.; Walker, P.L.; Girard, I.J.; Rashid, K.Y.; de Kievit, T.; Fernando, W.D.; Whyard, S.; Belmonte, M.F. Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Lau, S.E.; Schwarzacher, T.; Othman, R.Y.; Harikrishna, J.A. dsRNA silencing of an R2R3-MYB transcription factor affects flower cell shape in a Dendrobium hybrid. BMC Plant Biol. 2015, 15, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davalos, A.; Henriques, R.; Latasa, M.J.; Laparra, M.; Coca, M. Literature review of baseline information on non-coding RNA (ncRNA) to support the risk assessment of ncRNA-based genetically modified plants for food and feed. EFSA Supp. Publ. 2019, 16, 220. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Reeves, P.T.; Hong, B.T.; Mitter, N. A perspective on RNAi-based biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Casacuberta, J.M.; Devos, Y.; du Jardin, P.; Ramon, M.; Vaucheret, H.; Nogue, F. Biotechnological uses of RNAi in plants: Risk assessment considerations. Trends Biotechnol. 2015, 33, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, N.; Devos, Y.; Alvarez-Alfageme, F.; Lanzoni, A.; Waigmann, E. Risk assessment considerations for genetically modified RNAi plants: EFSA’s activities and perspective. Front. Plant Sci. 2020, 11, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, Y.; Yamada, T.; Matsumiya, T.; Ui-Tei, K.; Saigo, K.; Morishita, S. dsCheck: Highly sensitive off-target search software for double-stranded RNA-mediated RNA interference. Nucleic Acids Res. 2005, 33, 589–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| S.No | Ornamental Plants Sold | Turnover (Million Euros) |
|---|---|---|
| 1 | Rose | 696 |
| 2 | Chrysanthemum | 328 |
| 3 | Phalaenopsis | 460 |
| 4 | Tulip | 285 |
| 5 | Gerbera | 148 |
| 6 | Lily | 144 |
| 7 | Kalanchoe | 65 |
| 8 | Anthurium | 60 |
| 9 | Potted rose | 57 |
| 10 | Lavender | 20 |
| Crop | Gene | Disease Resistance | Reference |
|---|---|---|---|
| Rose (Rosa hybrida) | Ace-AMP1 | powdery mildew (Podosphaera pannosa) | [82] |
| rice chitinase | powdery mildew (P. pannosa) | [83] | |
| RhMLO1, RgMLO6, RlMLO7 | powdery mildew (P. pannosa) | [87,88] | |
| rice chitinase | black spot (Diplocarpon rosae) | [90] | |
| chitinases, glucanases, and RIPs | black spot (D. rosae) | [75] | |
| Rdr1 | black spot (D. rosae) | [56] | |
| Chrysanthemum (Chrysanthemum morifolium) | PGIP | Alternaria leaf spot (Septoria chrysanthemi) | [100] |
| hairpinXoo | leaf spot (Alternaria tenuissima) | [101] | |
| chiII | leaf spot (Septoria obesa) | [102] | |
| RCC2 | gray mold (B. cinerea) | [103] | |
| CaXMT1, CaMXMT1, CaDXMT1 | gray mold (B. cinerea) | [104] | |
| Cry1Ab and sarcotoxin IA | white rust (P. horiana) | [108] | |
| CmWRKY15-1 | white rust | [109] | |
| Petunia (Petunia hybrida) | endochitinase and osmotin | gray mold (B. cinerea) | [114] |
| WD (Wasabi defensin) | gray mold (B. cinerea) | [115] | |
| NIC (Nakamura Ikuo Chitinase) | gray mold (B. cinerea) | [116] | |
| Lily (Lilium) | RCH10 chitinase | gray mold (B. cinerea) | [119] |
| Ire-miR159 | gray mold (B. elliptica) | [120] | |
| Carnation (Dianthus caryophyllus) | PR-1, osmotin, chitinase | Fusarium wilt (F. oxysporum) | [128] |
| bacterial chitinase | Fusarium wilt (F. oxysporum) | [129] | |
| jasmonate methyl transferase | Fusarium wilt (F. oxysporum) | [130] | |
| Gladioulus (Gladiolus communis) | D4E1 | Fusarium wilt (F.oxysporum) | [131] |
| Fungal exochitinase, endochitinase, bacterial chloroperoxidase | Fusarium wilt (F. oxysporum) | [132] | |
| Geranium (Pelargonium graveolens L. Herit.) | Ace-AMP1 | gray mold (B. cinerea) | [133] |
| African violets (Saintpaulia ionantha) | glucanase and chitinase | Fusarium and Pythium | [134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mekapogu, M.; Jung, J.-A.; Kwon, O.-K.; Ahn, M.-S.; Song, H.-Y.; Jang, S. Recent Progress in Enhancing Fungal Disease Resistance in Ornamental Plants. Int. J. Mol. Sci. 2021, 22, 7956. https://doi.org/10.3390/ijms22157956
Mekapogu M, Jung J-A, Kwon O-K, Ahn M-S, Song H-Y, Jang S. Recent Progress in Enhancing Fungal Disease Resistance in Ornamental Plants. International Journal of Molecular Sciences. 2021; 22(15):7956. https://doi.org/10.3390/ijms22157956
Chicago/Turabian StyleMekapogu, Manjulatha, Jae-A Jung, Oh-Keun Kwon, Myung-Suk Ahn, Hyun-Young Song, and Seonghoe Jang. 2021. "Recent Progress in Enhancing Fungal Disease Resistance in Ornamental Plants" International Journal of Molecular Sciences 22, no. 15: 7956. https://doi.org/10.3390/ijms22157956