
Structural and Biochemical Characterization of a Dye-Decolorizing Peroxidase from Dictyostelium discoideum

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
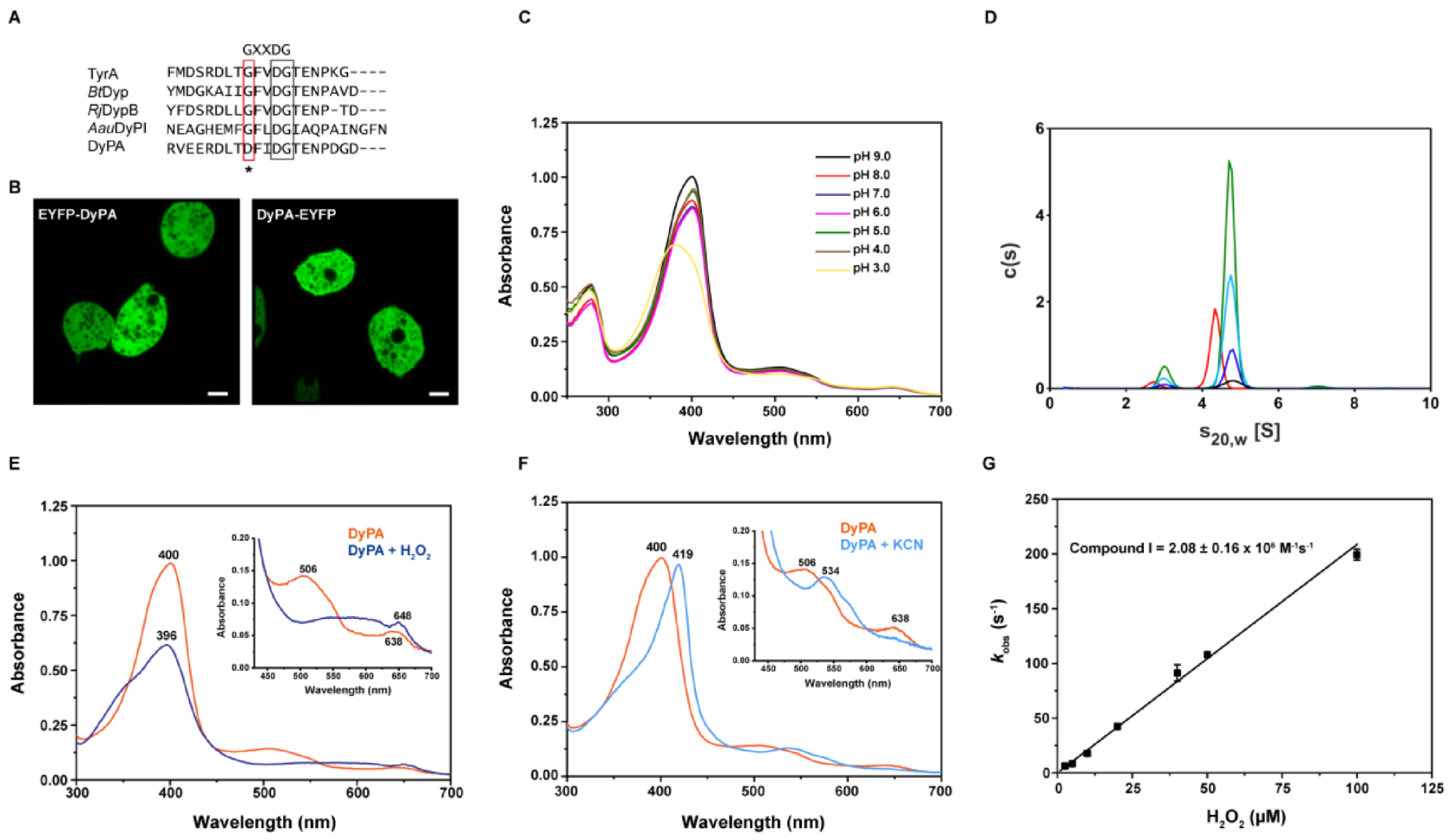
2.1. Localization of Dictyostelium DyPA
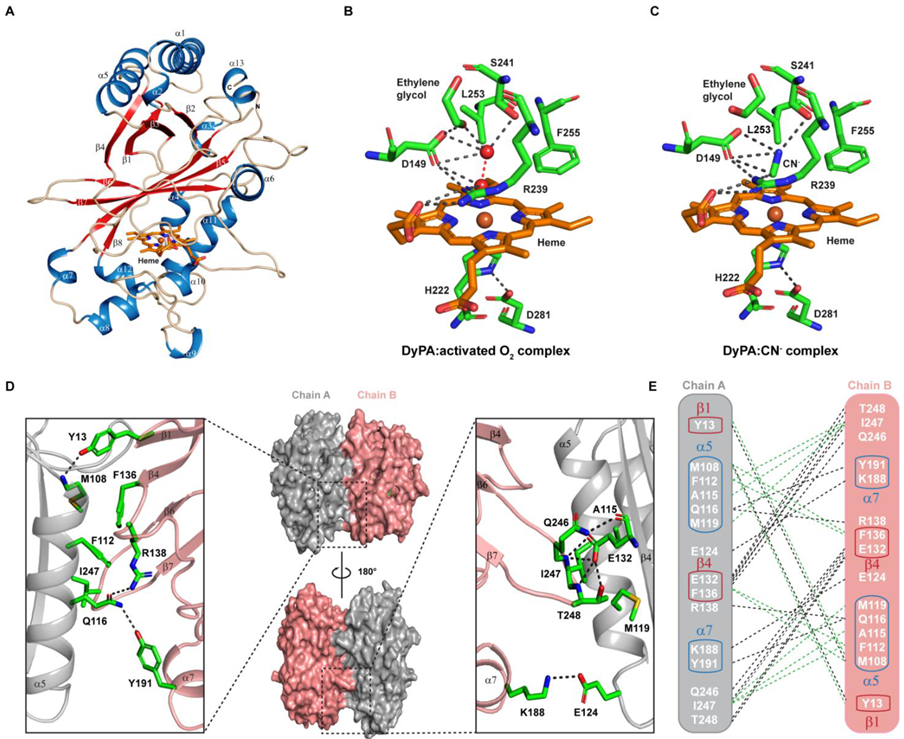
2.2. Analysis of the Oligomerization State of Dictyostelium DyPA by Analytical Ultracentrifugation
2.3. Absorption Spectra of Dictyostelium DyPA in the Presence of Peroxide or Cyanide and the Formation of Compound I
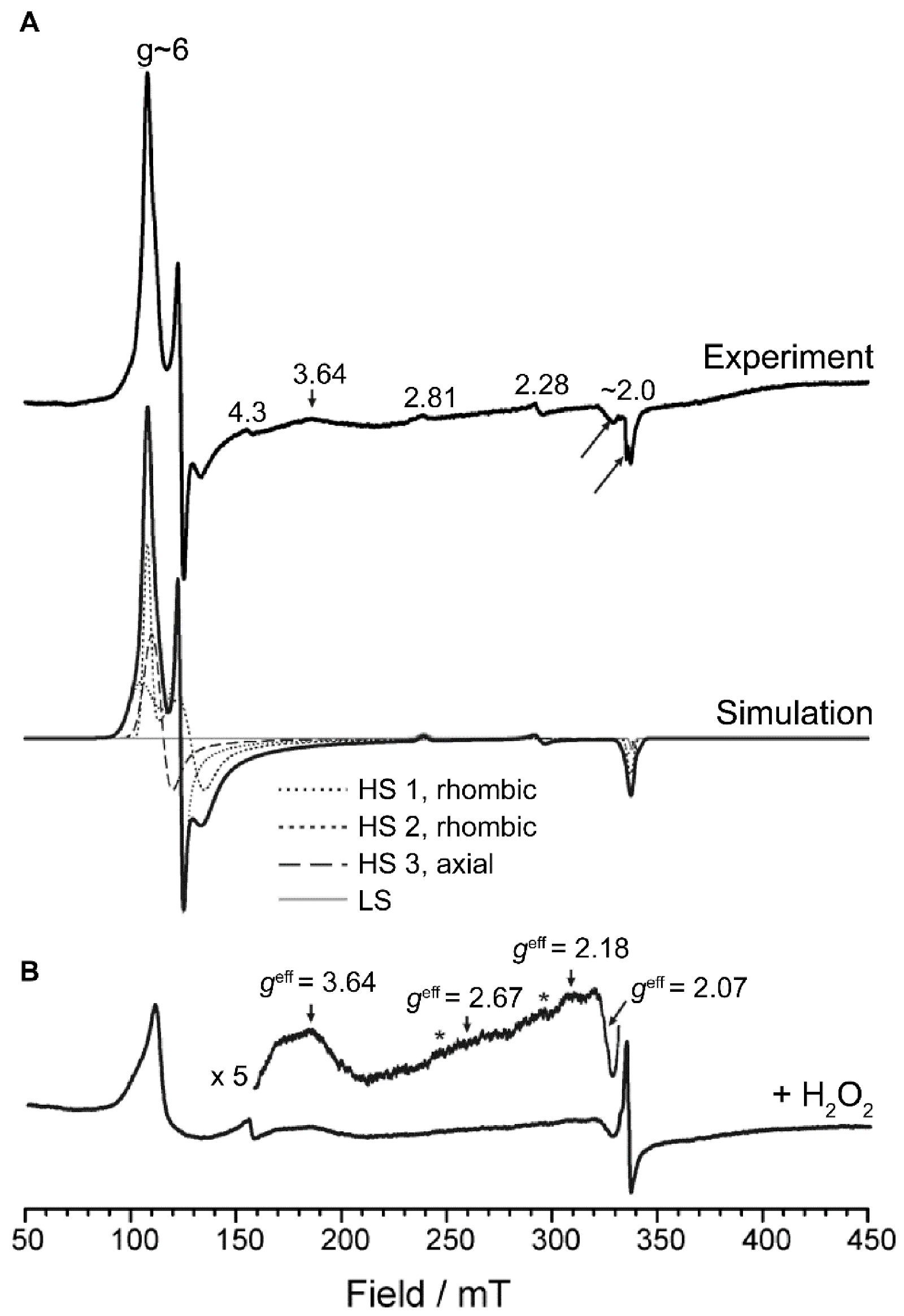
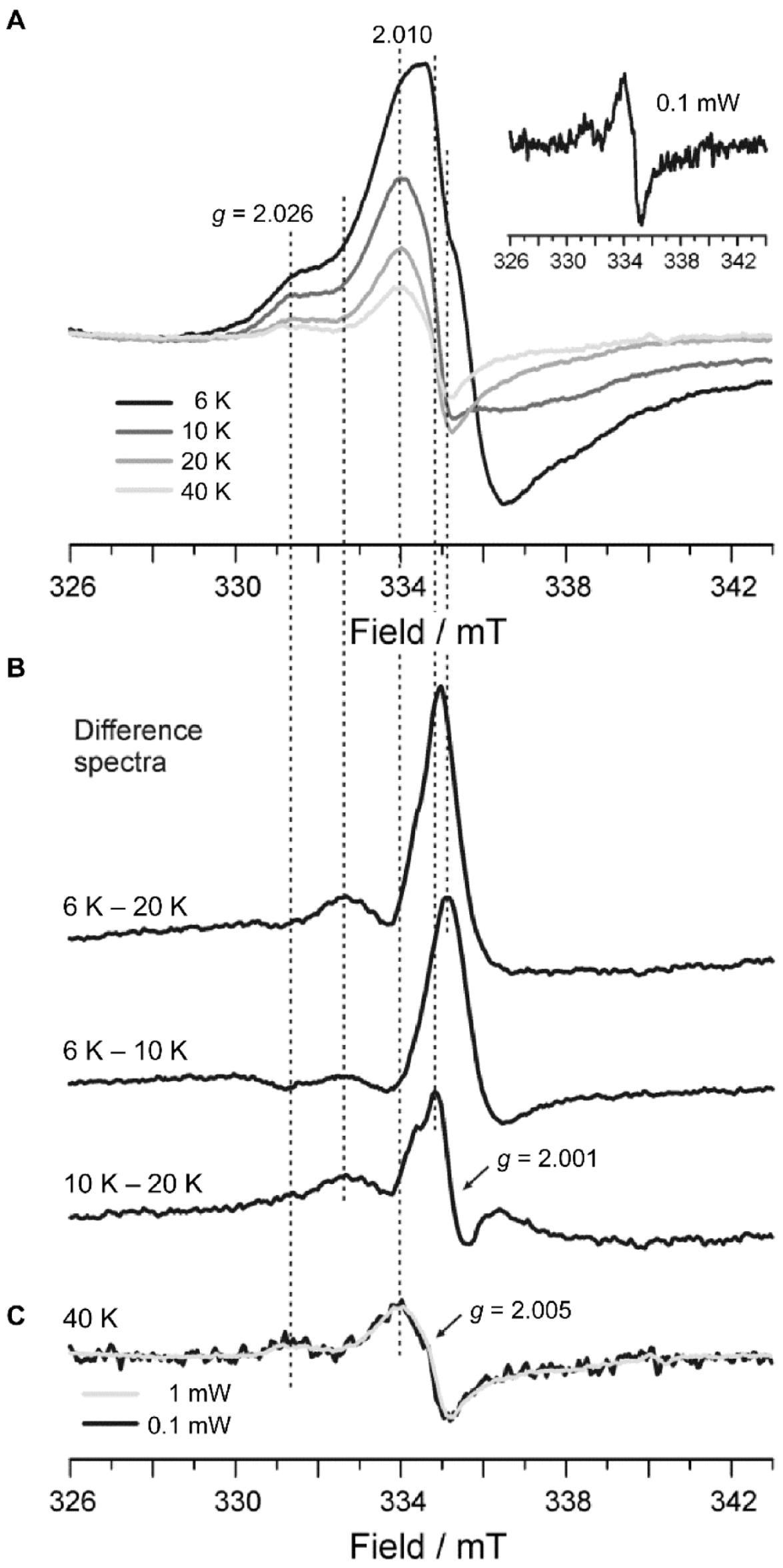
2.4. Electron Paramagnetic Resonance Spectroscopy
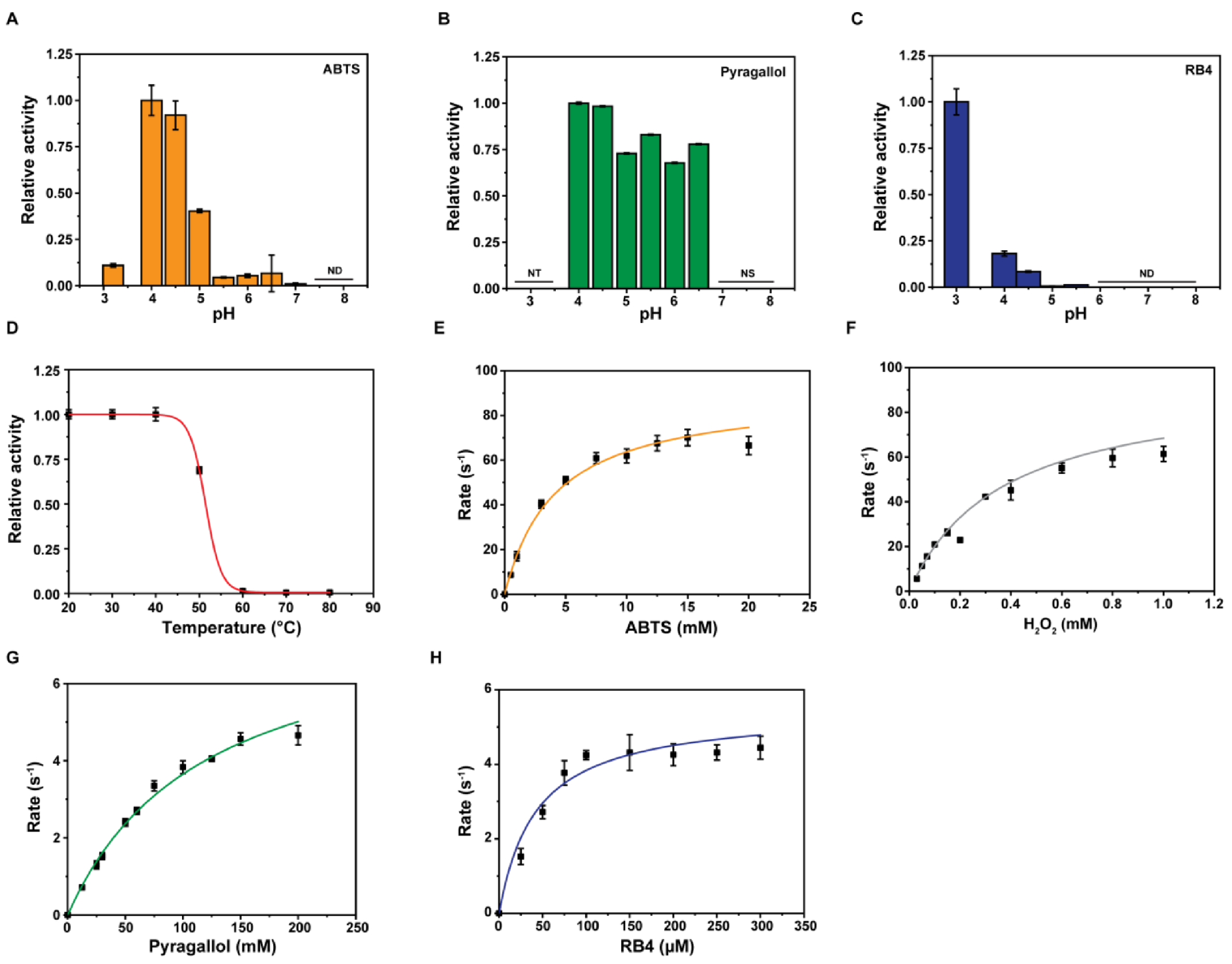
2.5. Substrate Specificities of Dictyostelium DyPA
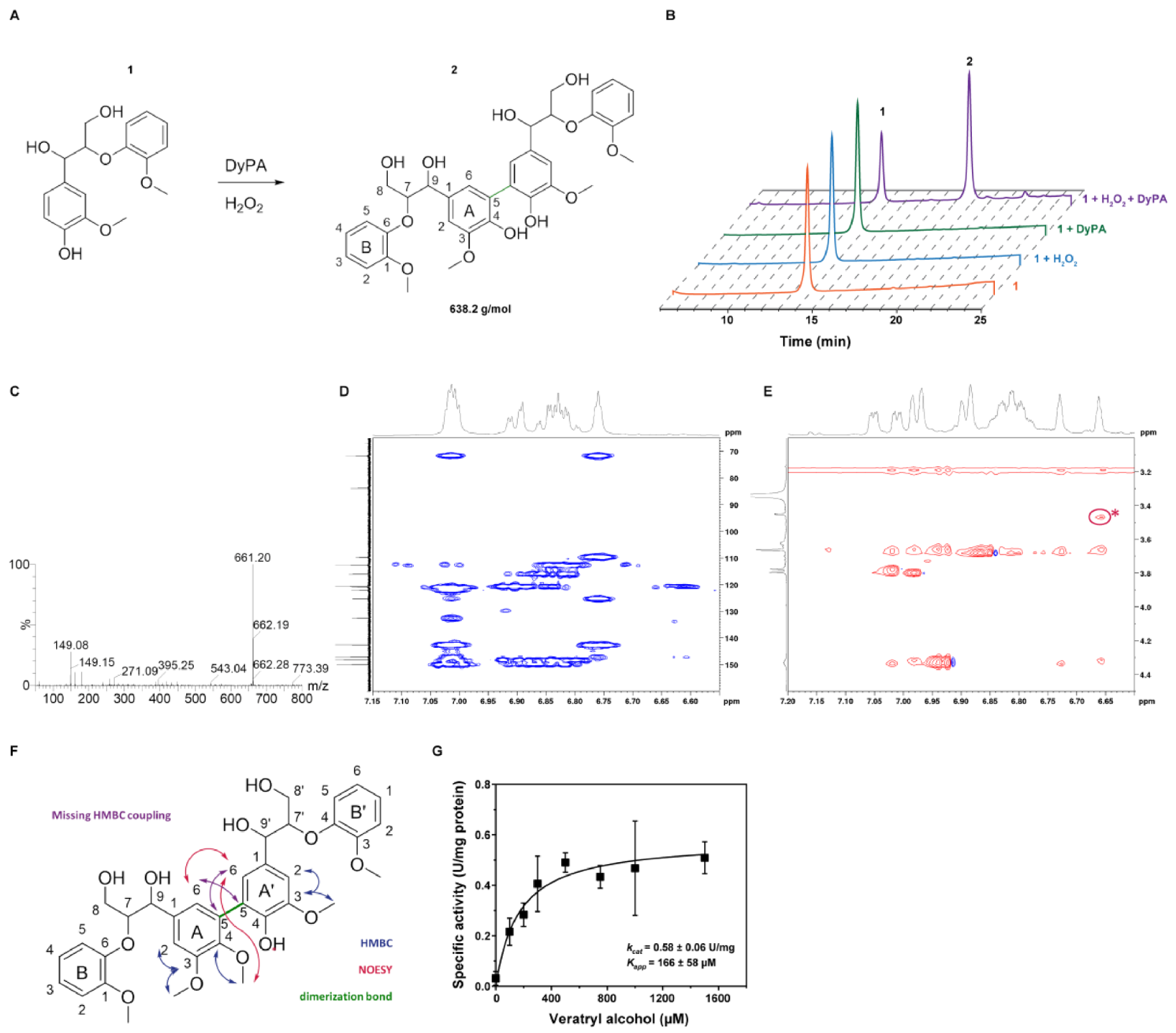
2.6. Oxidation of ß-Aryl Ether Lignin Model Substrate and Veratryl Alcohol
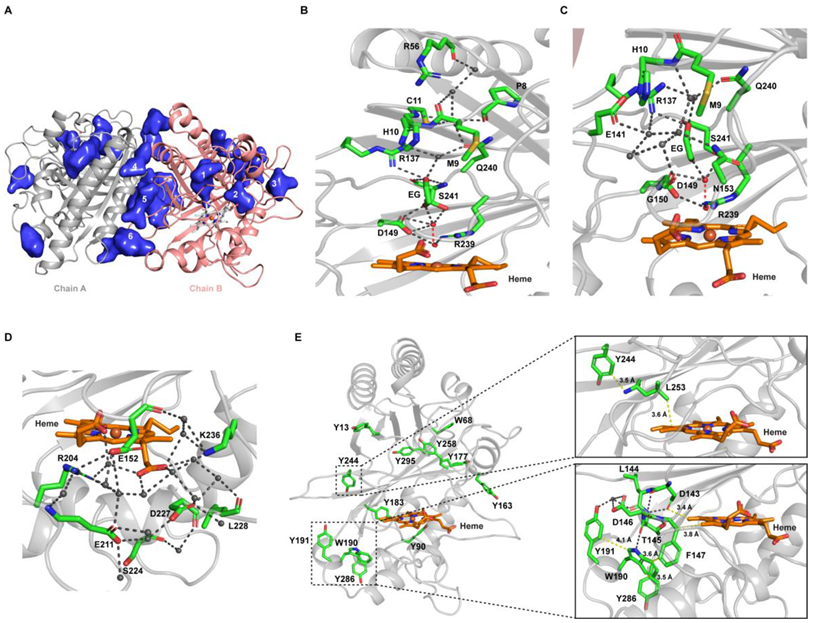
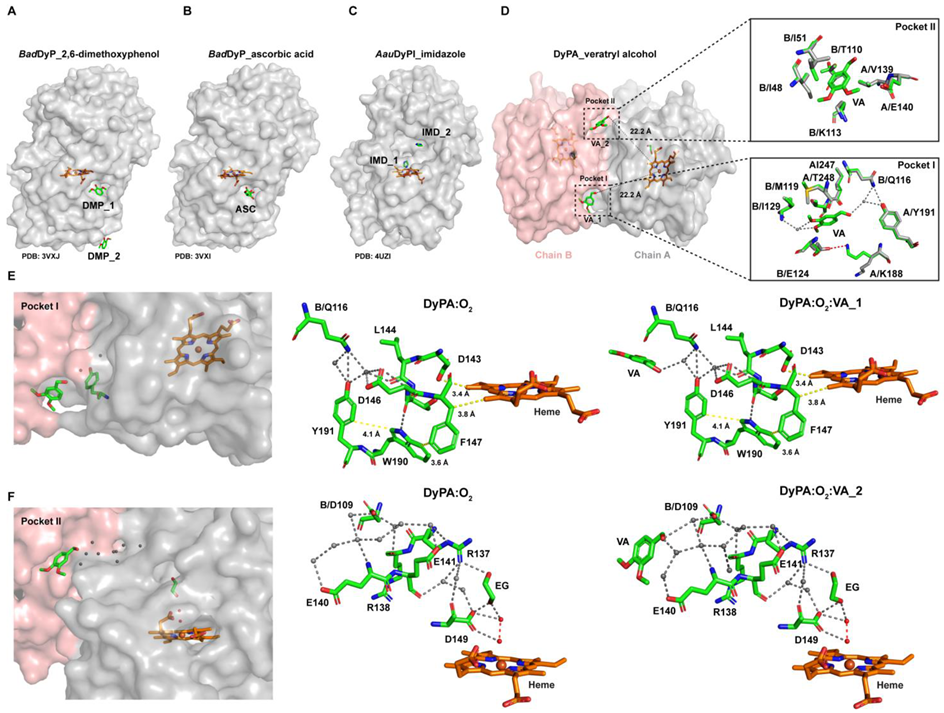
2.7. Structural Features of Dictyostelium DyPA:O2, Dictyostelium DyPA:CN− Complex and Dictyostelium DyPA:O2:VA Complexes
3. Materials and Methods
3.1. Chemicals
3.2. Protein Purification
3.3. Plasmid Construction, Cell Culture, and Fluorescence Microscopy
3.4. UV-Visible Spectroscopy
3.5. Analytical Ultracentrifugation
3.6. Stopped-Flow Kinetics
3.7. Electron Paramagnetic Resonance Spectroscopy
3.8. EPR Spectra Simulation
3.9. Steady-State Kinetic Measurements
3.10. Thermal Stability of Dictyostelium DyPA
3.11. Oxidation of β-aryl Ether Lignin Model Substrate
3.12. NMR Spectroscopy
3.13. LC-MS Analysis
3.14. Crystallization, Data Collection, and Structure Determination
3.15. Bioinformatics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DyP | Dye-decolorizing peroxidase |
| EYFP | Enhanced yellow fluorescent protein |
| GGBGE | Guaiacylglycerol-β-guaiacyl ether |
| HMBC | Heteronuclear multiple bond correlation |
| HRP | Horseradish peroxidase |
| KCN | Potassium cyanide |
| NOESY | Nuclear Overhauser enhancement spectroscopy |
| PDB | Protein data bank |
| RMSD | Root mean square deviation |
| RB5 | Reactive black5 |
| QM/MM | Quantum mechanics and molecular mechanics |
| TAT | Twin-arginine transport |
| mT | milliTesla |
| VA | Veratryl alcohol |
References
- Kelly, B.; Carrizo, G.E.; Edwards-Hicks, J.; Sanin, D.E.; Stanczak, M.A.; Priesnitz, C.; Flachsmann, L.J.; Curtis, J.D.; Mittler, G.; Musa, Y.; et al. Sulfur sequestration promotes multicellularity during nutrient limitation. Nature 2021, 591, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Shaulsky, G.; Kessin, R.H. The cold war of the social amoebae. Curr. Biol. 2007, 17, R684–R692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pears, C.J.; Gross, J.D. Microbe Profile: Dictyostelium discoideum: Model system for development, chemotaxis and biomedical research. Microbiology (Reading) 2021, 167. [Google Scholar] [CrossRef]
- Savelli, B.; Li, Q.; Webber, M.; Jemmat, A.M.; Robitaille, A.; Zamocky, M.; Mathé, C.; Dunand, C. RedoxiBase: A database for ROS homeostasis regulated proteins. Redox Biol. 2019, 26, 101247. [Google Scholar] [CrossRef]
- Linde, D.; Ayuso-Fernandez, I.; Laloux, M.; Aguiar-Cervera, J.E.; de Lacey, A.L.; Ruiz-Duenas, F.J.; Martínez, A.T. Comparing Ligninolytic Capabilities of Bacterial and Fungal Dye-Decolorizing Peroxidases and Class-II Peroxidase-Catalases. Int. J. Mol. Sci. 2021, 22, 2629. [Google Scholar] [CrossRef]
- Ogola, H.J.O.; Kamiike, T.; Hashimoto, N.; Ashida, H.; Ishikawa, T.; Shibata, H.; Sawa, Y. Molecular characterization of a novel peroxidase from the cyanobacterium Anabaena sp. strain PCC 7120. Appl. Environ. Microbiol. 2009, 75, 7509–7518. [Google Scholar] [CrossRef] [Green Version]
- Van Bloois, E.; Pazmiño, D.E.T.; Winter, R.T.; Fraaije, M.W. A robust and extracellular heme-containing peroxidase from Thermobifida fusca as prototype of a bacterial peroxidase superfamily. Appl. Microbiol. Biotechnol. 2010, 86, 1419–1430. [Google Scholar] [CrossRef] [Green Version]
- Büttner, E.; Ullrich, R.; Strittmatter, E.; Piontek, K.; Plattner, D.A.; Hofrichter, M.; Liers, C. Oxidation and nitration of mononitrophenols by a DyP-type peroxidase. Arch. Biochem. Biophys. 2015, 574, 86–92. [Google Scholar] [CrossRef]
- Kim, S.J.; Shoda, M. Decolorization of molasses and a dye by a newly isolated strain of the fungus Geotrichum candidum Dec 1. Biotechnol. Bioeng. 1999, 62, 114–119. [Google Scholar] [CrossRef]
- Roberts, J.N.; Singh, R.; Grigg, J.C.; Murphy, M.E.; Bugg, T.D.; Eltis, L.D. Characterization of dye-decolorizing peroxidases from Rhodococcus jostii RHA1. Biochemistry 2011, 50, 5108–5119. [Google Scholar] [CrossRef]
- Fernández-Fueyo, E.; Linde, D.; Almendral, D.; López-Lucendo, M.F.; Ruiz-Dueñas, F.J.; Martínez, A.T. Description of the first fungal dye-decolorizing peroxidase oxidizing manganese(II). Appl. Microbiol. Biotechnol. 2015, 99, 8927–8942. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.E.; Barros, T.; Chang, M.C.Y. Identification and Characterization of a Multifunctional Dye Peroxidase from a Lignin-Reactive Bacterium. ACS Chem. Biol. 2012, 7, 2074–2081. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Bugg, T.D. Characterisation of Dyp-type peroxidases from Pseudomonas fluorescens Pf-5: Oxidation of Mn(II) and polymeric lignin by Dyp1B. Arch. Biochem. Biophys. 2015, 574, 93–98. [Google Scholar] [CrossRef]
- Singh, R.; Eltis, L.D. The multihued palette of dye-decolorizing peroxidases. Arch. Biochem. Biophys. 2015, 574, 56–65. [Google Scholar] [CrossRef]
- Sugano, Y. DyP-type peroxidases comprise a novel heme peroxidase family. Cell. Mol. Life Sci. 2009, 66, 1387–1403. [Google Scholar] [CrossRef]
- Yoshida, T.; Sugano, Y. A structural and functional perspective of DyP-type peroxidase family. Arch. Biochem. Biophys. 2015, 574, 49–55. [Google Scholar] [CrossRef]
- Sugano, Y.; Muramatsu, R.; Ichiyanagi, A.; Sato, T.; Shoda, M. DyP, a unique dye-decolorizing peroxidase, represents a novel heme peroxidase family: ASP171 replaces the distal histidine of classical peroxidases. J. Biol. Chem. 2007, 282, 36652–36658. [Google Scholar] [CrossRef] [Green Version]
- Eichinger, L.; Pachebat, J.A.; Glöckner, G.; Rajandream, M.-A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Roberts, J.N.; Hardiman, E.M.; Singh, R.; Eltis, L.D.; Bugg, T.D. Identification of DypB from Rhodococcus jostii RHA1 as a lignin peroxidase. Biochemistry 2011, 50, 5096–5107. [Google Scholar] [CrossRef]
- Rai, A.; Fedorov, R.; Manstein, D.J. Expression, purification and crystallization of a dye-decolourizing peroxidase from Dictyostelium discoideum. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 252–255. [Google Scholar] [CrossRef] [Green Version]
- Schuck, P. Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and lamm equation modeling. Biophys. J. 2000, 78, 1606–1619. [Google Scholar] [CrossRef] [Green Version]
- Lebowitz, J.; Lewis, M.S.; Schuck, P. Modern analytical ultracentrifugation in protein science: A tutorial review. Protein Sci. 2002, 11, 2067–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfanzagl, V.; Nys, K.; Bellei, M.; Michlits, H.; Mlynek, G.; Battistuzzi, G.; Djinovic-Carugo, K.; Van Doorslaer, S.; Furtmüller, P.G.; Hofbauer, S.; et al. Roles of distal aspartate and arginine of B-class dye-decolorizing peroxidase in heterolytic hydrogen peroxide cleavage. J. Biol. Chem. 2018, 293, 14823–14838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lučić, M.; Svistunenko, D.A.; Wilson, M.T.; Chaplin, A.K.; Davy, B.; Ebrahim, A.; Axford, D.; Tosha, T.; Sugimoto, H.; Owada, S.; et al. Serial Femtosecond Zero Dose Crystallography Captures a Water-Free Distal Heme Site in a Dye-Decolorising Peroxidase to Reveal a Catalytic Role for an Arginine in Fe(IV) =O Formation. Angew. Chem. Int. Ed. Engl. 2020, 59, 21656–21662. [Google Scholar] [CrossRef]
- Edwards, S.L.; Poulos, T.L. Ligand binding and structural perturbations in cytochrome c peroxidase. A crystallographic study. J. Biol. Chem. 1990, 265, 2588–2595. [Google Scholar] [CrossRef]
- Yoshida, T.; Tsuge, H.; Konno, H.; Hisabori, T.; Sugano, Y. The catalytic mechanism of dye-decolorizing peroxidase DyP may require the swinging movement of an aspartic acid residue. FEBS J. 2011, 278, 2387–2394. [Google Scholar] [CrossRef]
- Fukuyama, K.; Kunishima, N.; Amada, F.; Kubota, T.; Matsubara, H. Crystal structures of cyanide- and triiodide-bound forms of Arthromyces ramosus peroxidase at different pH values. Perturbations of active site residues and their implication in enzyme catalysis. J. Biol. Chem. 1995, 270, 21884–21892. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Lopez, J.N.; Smith, A.T.; Thorneley, R.N.F. Role of arginine 38 in horseradish peroxidase. A critical residue for substrate binding and catalysis. J. Biol. Chem. 1996, 271, 4023–4030. [Google Scholar] [CrossRef] [Green Version]
- Wengenack, N.L.; Todorovic, S.; Yu, L.; Rusnak, F. Evidence for differential binding of isoniazid by Mycobacterium tuberculosis KatG and the isoniazid-resistant mutant KatG(S315T). Biochemistry 1998, 37, 15825–15834. [Google Scholar] [CrossRef]
- Ivancich, A.; Jakopitsch, C.; Auer, M.; Un, S.; Obinger, C. Protein-based radicals in the catalase-peroxidase of synechocystis PCC6803: A multifrequency EPR investigation of wild-type and variants on the environment of the heme active site. J. Am. Chem. Soc. 2003, 125, 14093–14102. [Google Scholar] [CrossRef]
- Slappendel, S.; Veldink, G.A.; Vliegenthart, J.F.; Aasa, R.; Malmstrom, B.G. EPR spectroscopy of soybean lipoxygenase-1. Description and quantification of the high-spin fe(III) signals. Biochim. Biophys. Acta Protein Struct. 1981, 667, 77–86. [Google Scholar] [CrossRef]
- Hirst, J.; Wilcox, S.K.; Ai, J.; Moenne-Loccoz, P.; Loehr, T.M.; Goodin, D.B. Replacement of the axial histidine ligand with imidazole in cytochrome c peroxidase. 2. Effects on heme coordination and function. Biochemistry 2001, 40, 1274–1283. [Google Scholar] [CrossRef]
- Yonetani, T.; Anni, H. Yeast cytochrome c peroxidase. Coordination and spin states of heme prosthetic group. J. Biol. Chem. 1987, 262, 9547–9554. [Google Scholar] [CrossRef]
- Schulz, C.E.; Devaney, P.W.; Winkler, H.; Debrunner, P.G.; Doan, N.; Chiang, R.; Rutter, R.; Hager, L.P. Horseradish peroxidase compound I: Evidence for spin coupling between the heme iron and a ‘free’ radical. FEBS Lett. 1979, 103, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Switala, J.; Loewen, P.C.; Ivancich, A. Two [Fe(IV)=O Trp*] intermediates in M. tuberculosis catalase-peroxidase discriminated by multifrequency (9-285 GHz) EPR spectroscopy: Reactivity toward isoniazid. J. Am. Chem Soc. 2007, 129, 15954–15963. [Google Scholar] [CrossRef]
- Ivancich, A.; Mazza, G.; Desbois, A. Comparative electron paramagnetic resonance study of radical intermediates in turnip peroxidase isozymes. Biochemistry 2001, 40, 6860–6866. [Google Scholar] [CrossRef]
- Fielding, A.J.; Singh, R.; Boscolo, B.; Loewen, P.C.; Ghibaudi, E.M.; Ivancich, A. Intramolecular electron transfer versus substrate oxidation in lactoperoxidase: Investigation of radical intermediates by stopped-flow absorption spectrophotometry and (9-285 GHz) electron paramagnetic resonance spectroscopy. Biochemistry 2008, 47, 9781–9792. [Google Scholar] [CrossRef]
- Yu, M.A.; Egawa, T.; Shinzawa-Itoh, K.; Yoshikawa, S.; Guallar, V.; Yeh, S.-R.; Rousseau, D.L.; Gerfen, G.J. Two tyrosyl radicals stabilize high oxidation states in cytochrome C oxidase for efficient energy conservation and proton translocation. J. Am. Chem. Soc. 2012, 134, 4753–4761. [Google Scholar] [CrossRef] [Green Version]
- Miki, Y.; Pogni, R.; Acebes, S.; Lucas, F.; Fernández-Fueyo, E.; Baratto, M.C.; Fernández, M.I.; de los Ríos, V.; Ruiz-Dueñas, F.J.; Sinicropi, A.; et al. Formation of a tyrosine adduct involved in lignin degradation by Trametopsis cervina lignin peroxidase: A novel peroxidase activation mechanism. Biochem. J. 2013, 452, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Chaplin, A.K.; Chicano, T.M.; Hampshire, B.V.; Wilson, M.T.; Hough, M.A.; Svistunenko, D.A.; Worrall, J.A.R. An Aromatic Dyad Motif in Dye Decolourising Peroxidases Has Implications for Free Radical Formation and Catalysis. Chemistry 2019, 25, 6141–6153. [Google Scholar] [CrossRef] [Green Version]
- Nys, K.; Furtmüller, P.G.; Obinger, C.; Van Doorslaer, S.; Pfanzagl, V. On the Track of Long-Range Electron Transfer in B-Type Dye-Decolorizing Peroxidases: Identification of a Tyrosyl Radical by Computational Prediction and Electron Paramagnetic Resonance Spectroscopy. Biochemistry 2021, 60, 1226–1241. [Google Scholar] [CrossRef] [PubMed]
- Peisach, J.; Blumberg, W.E.; Ogawa, S.; Rachmilewitz, E.A.; Oltzik, R. The effects of protein conformation on the heme symmetry in high spin ferric heme proteins as studied by electron paramagnetic resonance. J. Biol. Chem. 1971, 246, 3342–3355. [Google Scholar] [CrossRef]
- Chen, C.; Shrestha, R.; Jia, K.; Gao, P.F.; Geisbrecht, B.V.; Bossmann, S.H.; Shi, J.; Li, P. Characterization of Dye-decolorizing Peroxidase (DyP) from Thermomonospora curvata Reveals Unique Catalytic Properties of A-type DyPs. J. Biol. Chem. 2015, 290, 23447–23463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmanpour, R.; Rea, D.; Jamshidi, S.; Fülöp, V.; Bugg, T.D. Structure of Thermobifida fusca DyP-type peroxidase and activity towards Kraft lignin and lignin model compounds. Arch. Biochem. Biophys. 2016, 594, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Min, K.; Gong, G.; Woo, H.M.; Kim, Y.; Um, Y. A dye-decolorizing peroxidase from Bacillus subtilis exhibiting substrate-dependent optimum temperature for dyes and beta-ether lignin dimer. Sci. Rep. 2015, 5, 8245. [Google Scholar] [CrossRef]
- Zubieta, C.; Joseph, R.; Krishna, S.S.; McMullan, D.; Kapoor, M.; Axelrod, H.L.; Miller, M.D.; Abdubek, P.; Acosta, C.; Astakhova, T.; et al. Identification and structural characterization of heme binding in a novel dye-decolorizing peroxidase, TyrA. Proteins Struct. Funct. Bioinform. 2007, 69, 234–243. [Google Scholar] [CrossRef]
- Zubieta, C.; Krishna, S.S.; Kapoor, M.; Kozbial, P.; McMullan, D.; Axelrod, H.L.; Miller, M.D.; Abdubek, P.; Ambing, E.; Astakhova, T.; et al. Crystal structures of two novel dye-decolorizing peroxidases reveal a beta-barrel fold with a conserved heme-binding motif. Proteins Struct. Funct. Bioinform. 2007, 69, 223–233. [Google Scholar] [CrossRef]
- Strittmatter, E.; Liers, C.; Ullrich, R.; Wachter, S.; Hofrichter, M.; Plattner, D.A.; Piontek, K. First crystal structure of a fungal high-redox potential dye-decolorizing peroxidase: Substrate interaction sites and long-range electron transfer. J. Biol. Chem. 2013, 288, 4095–4102. [Google Scholar] [CrossRef] [Green Version]
- De Geus, D.C.; Thomassen, E.A.J.; Hagedoorn, P.-L.; Pannu, N.S.; van Duijn, E.; Abrahams, J.P. Crystal structure of chlorite dismutase, a detoxifying enzyme producing molecular oxygen. J. Mol. Biol. 2009, 387, 192–206. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, Z.; Wang, J.; Cui, Y.; Liu, S.; Ma, Y.; Gu, L.; Xu, S. Crystal structure and biochemical features of dye-decolorizing peroxidase YfeX from Escherichia coli O157 Asp143 and Arg232 play divergent roles toward different substrates. Biochem. Biophys. Res. Commun. 2017, 484, 40–44. [Google Scholar] [CrossRef]
- Uchida, T.; Sasaki, M.; Tanaka, Y.; Ishimori, K. A Dye-Decolorizing Peroxidase from Vibrio cholerae. Biochemistry 2015, 54, 6610–6621. [Google Scholar] [CrossRef]
- Liu, X.; Du, Q.; Wang, Z.; Zhu, D.; Huang, Y.; Li, N.; Wei, T.; Xu, S.; Gu, L. Crystal structure and biochemical features of EfeB/YcdB from Escherichia coli O157: ASP235 plays divergent roles in different enzyme-catalyzed processes. J. Biol. Chem. 2011, 286, 14922–14931. [Google Scholar] [CrossRef] [Green Version]
- Bersuker, I.B. Electronic Structure and Properties of Transition Metal Compounds; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Sundaramoorthy, M.; Terner, J.; Poulos, T.L. The crystal structure of chloroperoxidase: A heme peroxidase--cytochrome P450 functional hybrid. Structure 1995, 3, 1367–1377. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Grigg, J.C.; Armstrong, Z.; Murphy, M.E.; Eltis, L.D. Distal heme pocket residues of B-type dye-decolorizing peroxidase: Arginine but not aspartate is essential for peroxidase activity. J. Biol. Chem. 2012, 287, 10623–10630. [Google Scholar] [CrossRef] [Green Version]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Yu, J.; Zhou, Y.; Tanaka, I.; Yao, M. Roll: A new algorithm for the detection of protein pockets and cavities with a rolling probe sphere. Bioinformatics 2010, 26, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Doyle, W.A.; Blodig, W.; Veitch, N.C.; Piontek, K.; Smith, A.T. Two substrate interaction sites in lignin peroxidase revealed by site-directed mutagenesis. Biochemistry 1998, 37, 15097–15105. [Google Scholar] [CrossRef]
- Choinowski, T.; Blodig, W.; Winterhalter, K.H.; Piontek, K. The crystal structure of lignin peroxidase at 1.70 A resolution reveals a hydroxy group on the cbeta of tryptophan 171: A novel radical site formed during the redox cycle. J. Mol. Biol. 1999, 286, 809–827. [Google Scholar] [CrossRef]
- Smith, A.T.; Doyle, W.A.; Dorlet, P.; Ivancich, A. Spectroscopic evidence for an engineered, catalytically active Trp radical that creates the unique reactivity of lignin peroxidase. Proc. Natl. Acad. Sci. USA 2009, 106, 16084–16089. [Google Scholar] [CrossRef] [Green Version]
- Linde, D.; Ruiz-Dueñas, F.J.; Fernández-Fueyo, E.; Guallar, V.; Hammel, K.E.; Pogni, R.; Martínez, A.T. Basidiomycete DyPs: Genomic diversity, structural-functional aspects, reaction mechanism and environmental significance. Arch. Biochem. Biophys. 2015, 574, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Strittmatter, E.; Wachter, S.; Liers, C.; Ullrich, R.; Hofrichter, M.; Plattner, D.A.; Piontek, K. Radical formation on a conserved tyrosine residue is crucial for DyP activity. Arch. Biochem. Biophys. 2013, 537, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Tsuge, H.; Hisabori, T.; Sugano, Y. Crystal structures of dye-decolorizing peroxidase with ascorbic acid and 2, 6-dimethoxyphenol. FEBS Lett. 2012, 586, 4351–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strittmatter, E.; Serrer, K.; Liers, C.; Ullrich, R.; Hofrichter, M.; Piontek, K.; Schleicher, E.; Plattner, D.A. The toolbox of Auricularia auricula-judae dye-decolorizing peroxidase—Identification of three new potential substrate-interaction sites. Arch. Biochem. Biophys. 2015, 574, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Dueñas, F.J.; Morales, M.; García, E.; Miki, Y.; Martinez, M.J.; Martínez, A.T. Substrate oxidation sites in versatile peroxidase and other basidiomycete peroxidases. J. Exp. Bot. 2009, 60, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Acebes, S.; Ruiz-Dueñas, F.J.; Toubes, M.; Sáez-Jiménez, V.; Pérez-Boada, M.; Lucas, M.F.; Martínez, A.T.; Guallar, V. Mapping the Long-Range Electron Transfer Route in Ligninolytic Peroxidases. J. Phys. Chem. B 2017, 121, 3946–3954. [Google Scholar] [CrossRef]
- Romero, J.O.; Fernández-Fueyo, E.; Avila-Salas, F.; Recabarren, R.; Alzate-Morales, J.; Martínez, A.T. Binding and Catalytic Mechanisms of Veratryl Alcohol Oxidation by Lignin Peroxidase: A Theoretical and Experimental Study. Comput. Struct. Biotechnol. J. 2019, 17, 1066–1074. [Google Scholar] [CrossRef]
- Efimov, I.; Badyal, S.K.; Metcalfe, C.L.; Macdonald, I.; Gumiero, A.; Raven, E.L.; Moody, P.C.E. Proton delivery to ferryl heme in a heme peroxidase: Enzymatic use of the Grotthuss mechanism. J. Am. Chem. Soc. 2011, 133, 15376–15383. [Google Scholar] [CrossRef]
- Knetsch, M.L.; Tsiavaliaris, G.; Zimmermann, S.; Rühl, U.; Manstein, D.J. Expression vectors for studying cytoskeletal proteins in Dictyostelium discoideum. J. Muscle Res. Cell Motil. 2002, 23, 605–611. [Google Scholar] [CrossRef]
- Rai, A.; Tzvetkov, N.; Manstein, D.J. Functional dissection of the dictyostelium discoideum dynamin B mitochondrial targeting sequence. PLoS ONE 2013, 8, e56975. [Google Scholar] [CrossRef]
- Rai, A.; Nöthe, H.; Tzvetkov, N.; Korenbaum, E.; Manstein, D.J. Dictyostelium dynamin B modulates cytoskeletal structures and membranous organelles. Cell. Mol. Life Sci. 2011, 68, 2751–2767. [Google Scholar] [CrossRef] [Green Version]
- Laue, M.T.; Shah, B.D.; Rigdeway, T.M.; Pelletier, S.L. Computer-Aided Interpretation of Analytical Sedimentation Data for Proteins; Royal Society of Chemistry: Cambridge, UK, 1992; pp. 90–125. [Google Scholar]
- Stoll, S.; Schweiger, A. EasySpin, a comprehensive software package for spectral simulation and analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. Sect. D Biol. Crystallogr. 1997, 53, 240–255. [Google Scholar] [CrossRef]
- Project, C.C. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 1994, 50, 760–763. [Google Scholar]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; López, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Holm, L. Benchmarking fold detection by DaliLite v.5. Bioinformatics 2019, 35, 5326–5327. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| gx | gy | gz | HStrainx1 | HStrainy1 | HStrainz1 | E/D2 | R3 | % | |
|---|---|---|---|---|---|---|---|---|---|
| HS 1, rh | 6.44 | 5.44 | 2.00 | 1087 | 183 | 146 | 0.0208 | 6.25% | 33 |
| HS 2, rh | 6.21 | 5.20 | 1.99 | 343 | 840 | 89 | 0.0210 | 6.31% | 44 |
| HS 3, ax | 5.97 | 1.97 | 841 | 111 | - | - | 19 | ||
| LS | 2.81 | 2.28 | 1.99 | 190 | 150 | 60 | - | - | 4 |
| Substrate | Km (mM) | kcat (s−1) | kcat/Km (M−1 s−1) |
|---|---|---|---|
| ABTS | 4.1 ± 0.4 | 89.9 ± 4.3 | 2.19 × 104 |
| H2O2 * | 0.37 ± 0.063 | 93.65 ± 9.1 | 2.53 × 105 |
| Pyrogallol | 119.0 ± 11.0 | 7.9 ± 0.4 | 66.38 |
| Reactive blue 4 | 0.04 ± 0.01 | 5.2 ± 0.38 | 1.3 × 105 |
| Veratryl alcohol | 0.166 ± 0.058 | 3.38 × 10−4 ± 0.35 × 10−4 | 2.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rai, A.; Klare, J.P.; Reinke, P.Y.A.; Englmaier, F.; Fohrer, J.; Fedorov, R.; Taft, M.H.; Chizhov, I.; Curth, U.; Plettenburg, O.; et al. Structural and Biochemical Characterization of a Dye-Decolorizing Peroxidase from Dictyostelium discoideum. Int. J. Mol. Sci. 2021, 22, 6265. https://doi.org/10.3390/ijms22126265
Rai A, Klare JP, Reinke PYA, Englmaier F, Fohrer J, Fedorov R, Taft MH, Chizhov I, Curth U, Plettenburg O, et al. Structural and Biochemical Characterization of a Dye-Decolorizing Peroxidase from Dictyostelium discoideum. International Journal of Molecular Sciences. 2021; 22(12):6265. https://doi.org/10.3390/ijms22126265
Chicago/Turabian StyleRai, Amrita, Johann P. Klare, Patrick Y. A. Reinke, Felix Englmaier, Jörg Fohrer, Roman Fedorov, Manuel H. Taft, Igor Chizhov, Ute Curth, Oliver Plettenburg, and et al. 2021. "Structural and Biochemical Characterization of a Dye-Decolorizing Peroxidase from Dictyostelium discoideum" International Journal of Molecular Sciences 22, no. 12: 6265. https://doi.org/10.3390/ijms22126265