
Glutathione Deficiency during Early Postnatal Development Causes Schizophrenia-Like Symptoms and a Reduction in BDNF Levels in the Cortex and Hippocampus of Adult Sprague–Dawley Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Impact of Chronic Administration of BSO and GBR 12909 during the Early Postnatal Life on the Development of Social Deficits in Adolescence and Adulthood
2.2. The Impact of Chronic Administration of BSO and GBR 12909 during the Early Postnatal Life on the Development of Cognitive Deficits in Adolescence and Adulthood
2.3. The Impact of Chronic Administration of BSO and GBR 12909 during the Early Postnatal Life on the Manifestation of Positive Lescence and Adulthood
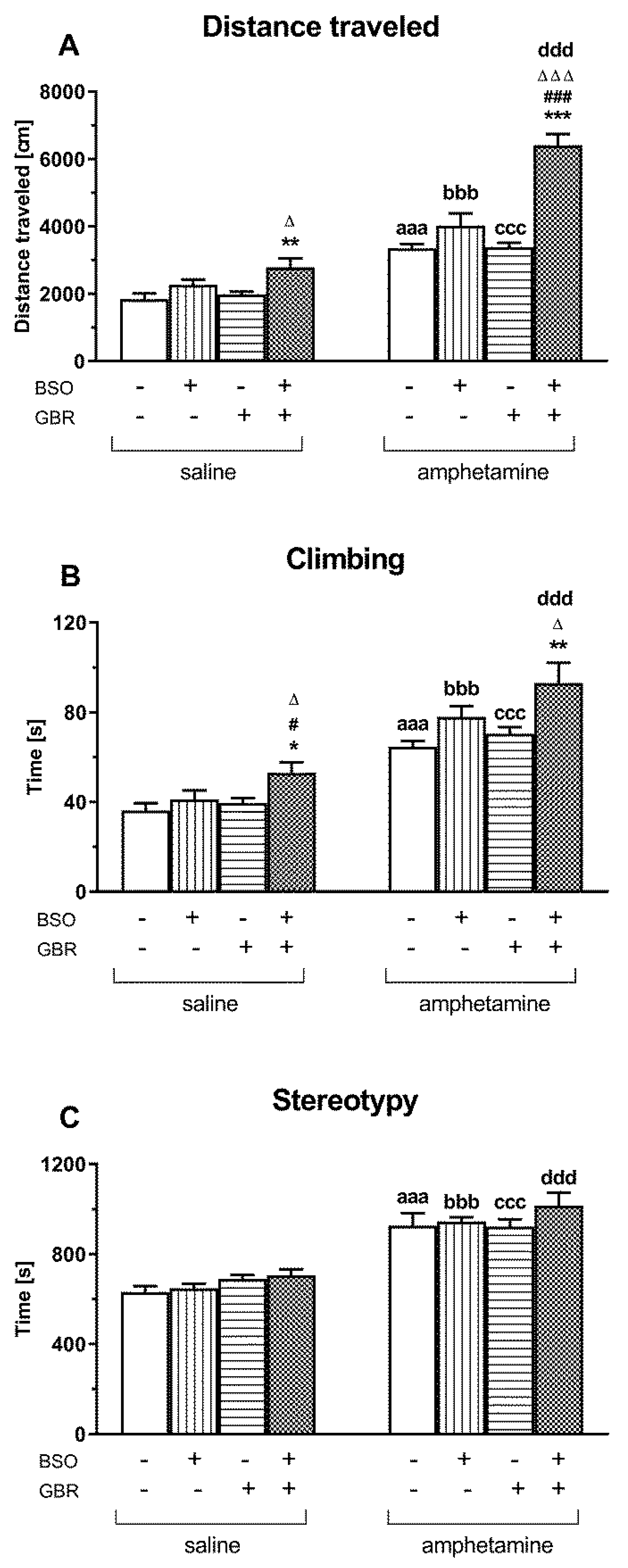
2.4. The Impact of Chronic Administration of BSO and GBR 12909 during the Early Postnatal Life on the Spontaneous and AMF-Induced Locomotor Activity and Stereotypy in Adult Rats Determined Using Actometers
2.5. The Impact of Chronic Administration of BSO and GBR 12909 during the Early Postnatal Life on BDNF mRNA and Protein Levels in the Prefrontal Cortex and Hippocampus of Adult Rats
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Treatment
5.2. Social Interaction Test
5.3. Novel Object Recognition Test
5.4. Open Field Test
5.5. Locomotor Activity Assessed in Actometers
5.6. BDNF Expression Analysis
5.7. ELISA Assay
5.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGrath, J.; Saha, S.; Chant, D.; Welham, J. Schizophrenia: A concise overview of incidence, prevalence, and mortality. Epidemiol. Rev. 2008, 30, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamminga, C.A.; Holcomb, H.H. Phenotype of schizophrenia: A review and formulation. Mol. Psychiatry 2005, 10, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Nuechterlein, K.H.; Ventura, J.; Subotnik, K.L.; Bartzokis, G. The early longitudinal course of cognitive deficits in schizophrenia. J. Clin. Psychiatry 2014, 75 (Suppl. S2), 25–29. [Google Scholar] [CrossRef] [Green Version]
- Keefe, R.S. The longitudinal course of cognitive impairment in schizophrenia: An examination of data from premorbid through posttreatment phases of illness. J. Clin. Psychiatry 2014, 75 (Suppl. S2), 8–13. [Google Scholar] [CrossRef]
- Kahn, R.S.; Keefe, R.S. Schizophrenia is a cognitive illness: Time for a change in focus. JAMA Psychiatry 2013, 70, 1107–1112. [Google Scholar] [CrossRef]
- Lepage, M.; Bodnar, M.; Bowie, C.R. Neurocognition: Clinical and functional outcomes in schizophrenia. Can. J. Psychiatry 2015, 59, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Strassnig, M.T.; Raykov, T.; O’Gorman, C.; Bowie, C.R.; Sabbag, S.; Durand, D.; Patterson, T.L.; Pinkham, A.; Penn, D.L.; Harvey, P.D. Determinants of different aspects of everyday outcome in schizophrenia: The roles of negative symptoms, cognition, and functional capacity. Schizophr. Res. 2015, 165, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassnig, M.; Bowie, C.; Pinkham, A.E.; Penn, D.; Twamley, E.W.; Patterson, T.L.; Harvey, P.D. Which levels of cognitive impairments and negative symptoms are related to functional deficits in schizophrenia? J. Psychiatr. Res. 2018, 104, 124–129. [Google Scholar] [CrossRef]
- van Os, J.; Rutten, B.P.; Poulton, R. Gene-environment interactions in schizophrenia: Review of epidemiological findings and future directions. Schizophr. Bull. 2008, 34, 1066–1082. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Levitt, P. Schizophrenia as a disorder of neurodevelopment. Annu. Rev. Neurosci. 2002, 25, 409–432. [Google Scholar] [CrossRef]
- Rapoport, J.L.; Gogtay, N. Childhood onset schizophrenia: Support for a progressive neurodevelopmental disorder. Int. J. Dev. Neurosci. 2011, 29, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzsimmons, J.; Kubicki, M.; Shenton, M.E. Review of functional and anatomical brain connectivity findings in schizophrenia. Curr. Opin. Psychiatry 2013, 26, 172–187. [Google Scholar] [CrossRef]
- Uhlhaas, P.J. Dysconnectivity, large-scale networks and neuronal dynamics in schizophrenia. Curr. Opin. Neurobiol. 2013, 23, 283–290. [Google Scholar] [CrossRef]
- Do, K.Q.; Cabungcal, J.H.; Frank, A.; Steullet, P.; Cuenod, M. Redox dysregulation, neurodevelopment, and schizophrenia. Curr. Opin. Neurobiol. 2019, 19, 220–230. [Google Scholar] [CrossRef]
- Steullet, P.; Cabungcal, J.H.; Kulak, A.; Kraftsik, R.; Chen, Y.; Dalton, T.P.; Cuenod, M.; Do, K.Q. Redox dysregulation affects the ventral but not dorsal hippocampus: Impairment of parvalbumin neurons, gamma oscillations, and related behaviors. J. Neurosci. 2010, 30, 2547–2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steullet, P.; Cabungcal, J.H.; Coyle, J.; Didriksen, M.; Gill, K.; Grace, A.A.; Hensch, T.K.; LaMantia, A.S.; Lindemann, L.; Maynard, T.M.; et al. Oxidative stress-driven parvalbumin interneuron impairment as a common mechanism in models of schizophrenia. Mol. Psychiatry 2017, 22, 936–943. [Google Scholar] [CrossRef]
- Bitanihirwe, B.K.; Woo, T.U. Oxidative stress in schizophrenia: An integrated approach. Neurosci. Biobehav. Rev. 2011, 35, 878–893. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.K.; Keshavan, M.S. Antioxidants, redox signaling, and pathophysiology in schizophrenia: An integrative view. Antioxid. Redox. Signal. 2011, 15, 2011–2035. [Google Scholar] [CrossRef] [PubMed]
- Pries, L.K.; Gülöksüz, S.; Kenis, G. DNA methylation in schizophrenia. Adv. Exp. Med. Biol. 2017, 978, 211–236. [Google Scholar]
- Ovenden, E.S.; McGregor, N.W.; Emsley, R.A.; Warnich, L. DNA methylation and antipsychotic treatment mechanisms in schizophrenia: Progress and future directions. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 81, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N. Role of methionine on epigenetic modification of DNA methylation and gene expression in animals. Anim. Nutr. 2018, 4, 11–16. [Google Scholar] [CrossRef]
- Do, K.Q.; Trabesinger, A.H.; Kirsten-Krüger, M.; Lauer, C.J.; Dydak, U.; Hell, D.; Holsboer, F.; Boesiger, P.; Cuénod, M. Schizophrenia: Glutathione deficit in cerebrospinal fluid and prefrontal cortex in vivo. Eur. J. Neurosci. 2000, 12, 3721–3728. [Google Scholar] [CrossRef]
- Yao, J.K.; Leonard, S.; Reddy, R. Altered glutathione redox state in schizophrenia. Dis. Markers 2006, 22, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Gawryluk, J.W.; Wang, J.F.; Andreazza, A.C.; Shao, L.; Young, L.T. Decreased levels of glutathione, the major brain antioxidant, in post-mortem prefrontal cortex from patients with psychiatric disorders. Int. J. Neuropsychopharmacol. 2011, 14, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altuntas, I.; Aksoy, H.; Coskun, I.; Cayköylü, A.; Akçay, F. Erythrocyte superoxide dismutase and glutathione peroxidase activities, and malondialdehyde and reduced glutathione levels in schizophrenic patients. Clin. Chem Lab. Med. 2000, 38, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Raffa, M.; Mechri, A.; Othman, L.B.; Fendri, C.; Gaha, L.; Kerkeni, A. Decreased glutathione levels and antioxidant enzyme activities in untreated and treated schizophrenic patients. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1178–1183. [Google Scholar] [CrossRef]
- Raffa, M.; Atig, F.; Mhalla, A.; Kerkeni, A.; Mechri, A. Decreased glutathione levels and impaired antioxidant enzyme activities in drug-naive first-episode schizophrenic patients. BMC Psychiatry 2011, 11, 124. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Tan, Y.L.; Zhou, D.F.; Cao, L.Y.; Wu, G.Y.; Haile, C.N.; Kosten, T.A.; Kosten, T.R. Disrupted antioxidant enzyme activity and elevated lipid peroxidation products in schizophrenic patients with tardive dyskinesia. J. Clin. Psychiatry 2017, 68, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Dietrich-Muszalska, A.; Olas, B.; Głowacki, R.; Bald, E. Oxidative/nitrative modifications of plasma proteins and thiols from patients with schizophrenia. Neuropsychobiology 2009, 59, 1–7. [Google Scholar] [CrossRef]
- Tosic, M.; Ott, J.; Barral, S.; Bovet, P.; Deppen, P.; Gheorghita, F.; Matthey, M.L.; Parnas, J.; Preisig, M.; Saraga, M.; et al. Schizophrenia and oxidative stress: Glutamate cysteine ligase modifier as a susceptibility gene. Am. J. Hum. Genet. 2006, 79, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Gysin, R.; Kraftsik, R.; Sandell, J.; Bovet, P.; Chappuis, C.; Conus, P.; Deppen, P.; Preisig, M.; Ruiz, V.; Steullet, P.; et al. Impaired glutathione synthesis in schizophrenia: Convergent genetic and functional evidence. Proc. Natl. Acad. Sci. USA 2007, 104, 16621–16626. [Google Scholar] [CrossRef] [Green Version]
- Gysin, R.; Kraftsik, R.; Boulat, O.; Bovet, P.; Conus, P.; Comte-Krieger, E.; Polari, A.; Steullet, P.; Preisig, M.; Teichmann, T.; et al. Genetic dysregulation of glutathione synthesis predicts alteration of plasma thiol redox status in schizophrenia. Antioxid. Redox. Signal. 2011, 15, 2003–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzawa, D.; Obata, T.; Shirayama, Y.; Nonaka, H.; Kanazawa, Y.; Yoshitome, E.; Takanashi, J.; Matsuda, T.; Shimizu, E.; Ikehira, H.; et al. Negative correlation between brain glutathione level and negative symptoms in schizophrenia: A 3T 1H-MRS study. PLoS ONE 2008, 3, e1944. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Erat, S.A.; Joseph, J.A. Spatial learning and memory deficits induced by dopamine administration with decreased glutathione. Free Radic. Biol. Med. 1998, 24, 1149–1158. [Google Scholar] [CrossRef]
- Almaguer-Melian, W.; Cruz-Aguado, R.; Bergado, J.A. Synaptic plasticity is impaired in rats with a low glutathione content. Synapse 2000, 38, 369–374. [Google Scholar] [CrossRef]
- Cruz-Aguado, R.; Almaguer-Melian, W.; Díaz, C.M.; Lorigados, L.; Bergado, J. Behavioral and biochemical effects of glutathione depletion in the rat brain. Brain Res. Bull. 2001, 55, 327–333. [Google Scholar] [CrossRef]
- Castagné, V.; Cuenod, M.; Do, K.Q. An animal model with relevance to schizophrenia: Sex-dependent cognitive deficits in Osteogenic Disorder-Shionogi rats induced by glutathione synthesis and dopamine uptake inhibition during development. Neuroscience 2004, 123, 821–834. [Google Scholar] [CrossRef]
- Castagné, V.; Rougemont, M.; Cuenod, M.; Do, K.Q. Low brain glutathione and ascorbic acid associated with dopamine uptake inhibition during rat’s development induce long-term cognitive deficit: Relevance to schizophrenia. Neurobiol. Dis. 2004, 15, 93–105. [Google Scholar] [CrossRef]
- Cabungcal, J.H.; Preissmann, D.; Delseth, C.; Cuénod, M.; Do, K.Q.; Schenk, F. Transitory glutathione deficit during brain development induces cognitive impairement in juvenile and adult rats: Relevance to schizophrenia. Neurobiol. Dis. 2007, 26, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Broquist, H.P. Buthionine sulfoximine, an experimental tool to induce glutathione deficiency: Elucidation of glutathione and ascorbate in their role as antioxidants. Nutr. Rev. 1992, 50, 110–111. [Google Scholar] [CrossRef]
- Masukawa, T.; Saim, M.; Tochino, Y. Methods for depleting brain glutathione. Life Sci. 1989, 44, 417–424. [Google Scholar] [CrossRef]
- Górny, M.; Wnuk, A.; Kamińska, A.; Kamińska, K.; Chwatko, G.; Bilska-Wilkosz, A.; Iciek, M.; Kajta, M.; Rogóż, Z.; Lorenc-Koci, E. Glutathione deficiency and alterations in the sulfur amino acid homeostasis during early postnatal development as potential triggering factors for schizophrenia-like behavior in adult rats. Molecules 2019, 24, 4253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laruelle, M.; Abi-Dargham, A.; van Dyck, C.H.; Gil, R.; D’Souza, C.D.; Erdos, J.; McCance, E.; Rosenblatt, W.; Fingado, C.; Zoghbi, S.S.; et al. Single photon emission computerized tomography imaging of amphetamine-induced dopamine release in drug-free schizophrenic subjects. Proc. Natl. Acad. Sci. USA 1996, 93, 9235–9240. [Google Scholar] [CrossRef] [Green Version]
- Breier, A.; Su, T.P.; Saunders, R.; Carson, R.E.; Kolachana, B.S.; de Bartolomeis, A.; Weinberger, D.R.; Weisenfeld, N.; Malhotra, A.K.; Eckelman, W.C.; et al. Schizophrenia is associated with elevated amphetamine-induced synaptic dopamine concentrations: Evidence from a novel positron emission tomography method. Proc. Natl. Acad. Sci. USA 1997, 94, 2569–2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abi-Dargham, A.; Gil, R.; Krystal, J.; Baldwin, R.; Seibyl, J.; Bowers, M.; van Dyck, C.H.; Charney, D.S.; Innis, R.B.; Laruelle, M. Increased striatal dopamine transmission in schizophrenia: Confirmation in a second cohort. Am. J. Psychiatry 1998, 155, 761–767. [Google Scholar] [PubMed]
- Levine, E.S.; Dreyfus, C.F.; Black, I.B.; Plummer, M.R. Brain-derived neurotrophic factor rapidly enhances synaptic transmission in hippocampal neurons via postsynaptic tyrosine kinase receptors. Proc. Natl. Acad. Sci. USA 1995, 92, 8074–8077. [Google Scholar] [CrossRef] [Green Version]
- Levine, E.S.; Crozier, R.A.; Black, I.B.; Plummer, M.R. Brain-derived neurotrophic factor modulates hippocampal synaptic transmission by increasing N-methyl-D-aspartic acid receptor activity. Proc. Natl. Acad. Sci. USA 1998, 95, 10235–10239. [Google Scholar] [CrossRef] [Green Version]
- Poo, M.M. Neurotrophins as synaptic modulators. Nat. Rev. Neurosci. 2001, 2, 24–32. [Google Scholar] [CrossRef]
- Lu, Y.; Christian, K.; Lu, B. BDNF: A key regulator for protein synthesis-dependent LTP and long-term memory? Neurobiol. Learn. Mem. 2008, 89, 312–323. [Google Scholar] [CrossRef] [Green Version]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [Green Version]
- Tyzio, R.; Holmes, G.L.; Ben-Ari, Y.; Roustem Khazipov, R. Timing of the developmental switch in GABA(A) mediated signaling from excitation to inhibition in CA3 rat hippocampus using gramicidin perforated patch and extracellular recordings. Epilepsia 2007, 48 (Suppl. S5), 96–105. [Google Scholar] [CrossRef]
- Ben-Ari, Y.; Gaiarsa, J.L.; Tyzio, R.; Khazipov, R. GABA: A pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol. Rev. 2007, 87, 1215–1284. [Google Scholar] [CrossRef] [PubMed]
- Dzhala, V.I.; Staley, K.J. Excitatory actions of endogenously released GABA contribute to initiation of ictal epileptiform activity in the developing hippocampus. J. Neurosci. 2003, 23, 1840–1846. [Google Scholar] [CrossRef] [Green Version]
- Khazipov, R.; Khalilov, I.; Tyzio, R.; Morozova, E.; Ben Ari, Y.; Holmes, G.L. Developmental changes in GABAergic actions and seizure susceptibility in the rat hippocampus. Eur. J. Neurosci. 2004, 19, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Khazipov, R.; Leinekugel, X.; Caillard, O.; Gaiarsa, J.L. GABA-A, NMDA and AMPA receptors: A developmentally regulated ‘menage a trois’. Trends Neurosci. 1997, 20, 523–529. [Google Scholar] [CrossRef]
- Leinekugel, X.; Khalilov, I.; McLean, H.; Caillard, O.; Gaiarsa, J.L.; Ben-Ari, Y.; Khazipov, R. GABA is the principal fast-acting excitatory transmitter in the neonatal brain. Adv. Neurol. 1999, 79, 189–201. [Google Scholar] [PubMed]
- Ben Ari, Y. Developing networks play a similar melody. Trends Neurosci. 2001, 24, 353–360. [Google Scholar] [CrossRef]
- Ben Ari, Y. Excitatory actions of GABA during development: The nature of the nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Zafra, F.; Hengerer, B.; Leibrock, J.; Thoenen, H.; Lindholm, D. Activity dependent regulation of BDNF and NGF mRNAs in the rat hippocampus is mediated by non-NMDA glutamate receptors. EMBO J. 1990, 9, 3545–3550. [Google Scholar] [CrossRef]
- Lindholm, D.; Castrén, E.; Berzaghi, M.; Blöchl, A.; Thoenen, H. Activity-dependent and hormonal regulation of neurotrophin mRNA levels in the brain--implications for neuronal plasticity. J. Neurobiol. 1994, 25, 1362–1372. [Google Scholar] [CrossRef]
- Marty, S.; Berzaghi, M.P.; Berninger, B. Neurotrophins and activity-dependent plasticity of cortical interneurons. Trends Neurosci. 1997, 20, 198–202. [Google Scholar] [CrossRef]
- Perkins, D.O.; Jeffries, C.D.; Do, K.Q. Potential roles of redox dysregulation in the development of schizophrenia. Biol. Psychiatry 2020, 88, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Nanda, D.; Telputt, J.; Collard, K.J. Changes in brain glutathione levels during postnatal development in the rat. Dev. Brain Res. 1996, 94, 238–241. [Google Scholar] [CrossRef]
- Danion, J.M.; Rizzo, L.; Bruant, A. Functional mechanisms underlying impaired recognition memory and conscious awareness in patients with schizophrenia. Arch. Gen. Psychiatry 1999, 56, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Doniger, G.M.; Silipo, G.; Rabinowicz, E.F.; Snotgrass, J.G.; Javitt, D.C. Impaired sensory processing as a basis for object-recognition deficits in schizophrenia. Am. J. Psychiatry 2001, 158, 1818–1826. [Google Scholar] [CrossRef]
- Heckers, S.; Curran, T.; Goff, D.; Rauch, S.L.; Fischman, A.J.; Alpert, N.M.; Schacter, D.L. Abnormalities in the thalamus and prefrontal cortex during episodic object recognition in schizophrenia. Biol. Psychiatry 2000, 48, 651–657. [Google Scholar] [CrossRef]
- Shoval, G.; Weizman, A. The possible role of neurotrophins in the pathogenesis and therapy of schizophrenia. Eur. Neuropsychopharmacol. 2005, 15, 319–329. [Google Scholar] [CrossRef]
- Buckley, P.F.; Mahadik, S.; Pillai, A.; Terry, A., Jr. Neurotrophins and schizophrenia. Schizophr. Res. 2007, 94, 1–11. [Google Scholar] [CrossRef]
- Steullet, P.; Cabungcal, J.H.; Monin, A.; Dwir, D.; O’Donnell, P.; Cuenod, M.; Do, K.Q. Redox dysregulation, neuroinflammation, and NMDA receptor hypofunction: A “central hub” in schizophrenia pathophysiology? Schizophr. Res. 2016, 176, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Lorenc-Koci, E. Dysregulation of glutathione synthesis in psychiatric disorders. In Studies on Psychiatric Disorders. Oxidative Stress in Applied Basic Research and Clinical Practice; Dietrich-Muszalska, A., Chauhan, V., Grignon, S., Eds.; Springer: New York, NY, USA; Berlin/Heidelberg, Germany; Dordrecht, The Netherlands; London, UK, 2015; pp. 269–299. [Google Scholar]
- Lipska, B.K.; Weinberger, D.R. To model a psychiatric disorder in animals. Schizophrenia as a reality test. Neuropsychopharmacology 2000, 23, 223–239. [Google Scholar] [CrossRef] [Green Version]
- Ashe, P.C.; Chlan-Fourney, J.; Juorio, A.V.; Li, X.M. Brain-derived neurotrophic factor (BDNF) mRNA in rats with neonatal ibotenic acid lesions of the ventral hippocampus. Brain Res. 2002, 956, 126–135. [Google Scholar] [CrossRef]
- Roceri, M.; Cirulli, F.; Pessina, C.; Peretto, P.; Racagni, G.; Riva, M.A. Postnatal repeated maternal deprivation produces age-dependent changes of brain-derived neurotrophic factor expression in selected rat brain regions. Biol. Psychiatry 2004, 55, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, F.; Bedogni, F.; Perez, J.; Racagni, G.; Riva, M.A. Corticostriatal brain-derived neurotrophic factor dysregulation in adult rats following prenatal stress. Eur. J. Neurosci. 2005, 20, 1348–1354. [Google Scholar] [CrossRef]
- Davis, K.L.; Kahn, R.S.; Ko, G.; Davidson, M. Dopamine in schizophrenia: A review and reconceptualization. Am. J. Psychiatry 1991, 148, 1474–1486. [Google Scholar] [PubMed]
- Abi-Dargham, A. Do we still believe in the dopamine hypothesis? New data bring new evidence. Int. J. Neuropsychopharmacol. 2004, 7 (Suppl. S1), S1–S5. [Google Scholar] [CrossRef]
- Howes, O.D.; Kapur, S. The dopamine hypothesis of schizophrenia: Version III--the final common pathway. Schizophr. Bull. 2009, 35, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Moghaddam, B. Cognitive dysfunction in schizophrenia: Convergence of gamma-aminobutyric acid and glutamate alterations. Arch. Neurol. 2006, 63, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A. The neurochemical circuitry of schizophrenia. Pharmacopsychiatry 2006, 39 (Suppl. S1), S10–S14. [Google Scholar] [CrossRef]
- Rogóż, Z.; Hereta, M.; Lorenc-Koci, E. Co-treatment with aripiprazole and mirtazapine reversed the schizophrenia-like behavior in adult Sprague-Dawley rats induced by glutathione deficit during early postnatal brain development. In Proceedings of the International Conference on Pharmaceutics and Drug Delivery Systems, Dubai, United Arab Emirates, 19–20 March 2021. [Google Scholar]
- Chowdhury, R.; Guitart-Masip, M.; Bunzeck, N.; Dolan, R.J.; Düzel, E. Dopamine modulates episodic memory persistence in old age. J. Neurosci. 2012, 32, 14193–14204. [Google Scholar] [CrossRef] [Green Version]
- Rocchetti, J.; Isingrini, E.; Dal Bo, G.; Sagheby, S.; Menegaux, A.; Tronche, F.; Levesque, D.; Moquin, L.; Gratton, A.; Wong, T.P.; et al. Presynaptic D2 dopamine receptors control long-term depression expression and memory processes in the temporal hippocampus. Biol. Psychiatry 2015, 77, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Kamińska, K.; Rogóż, Z. The effect of combined treatment with risperidone and antidepressants on the MK-801-induced deficits in the social interaction test in rats. Pharmacol. Rep. 2015, 67, 1183–1187. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lech, M.A.; Leśkiewicz, M.; Kamińska, K.; Rogóż, Z.; Lorenc-Koci, E. Glutathione Deficiency during Early Postnatal Development Causes Schizophrenia-Like Symptoms and a Reduction in BDNF Levels in the Cortex and Hippocampus of Adult Sprague–Dawley Rats. Int. J. Mol. Sci. 2021, 22, 6171. https://doi.org/10.3390/ijms22126171
Lech MA, Leśkiewicz M, Kamińska K, Rogóż Z, Lorenc-Koci E. Glutathione Deficiency during Early Postnatal Development Causes Schizophrenia-Like Symptoms and a Reduction in BDNF Levels in the Cortex and Hippocampus of Adult Sprague–Dawley Rats. International Journal of Molecular Sciences. 2021; 22(12):6171. https://doi.org/10.3390/ijms22126171
Chicago/Turabian StyleLech, Marta Anna, Monika Leśkiewicz, Kinga Kamińska, Zofia Rogóż, and Elżbieta Lorenc-Koci. 2021. "Glutathione Deficiency during Early Postnatal Development Causes Schizophrenia-Like Symptoms and a Reduction in BDNF Levels in the Cortex and Hippocampus of Adult Sprague–Dawley Rats" International Journal of Molecular Sciences 22, no. 12: 6171. https://doi.org/10.3390/ijms22126171