
Comparative TMT Proteomic Analysis Unveils Unique Insights into Helicoverpa armigera (Hübner) Resistance in Cajanus scarabaeoides (L.) Thouars

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
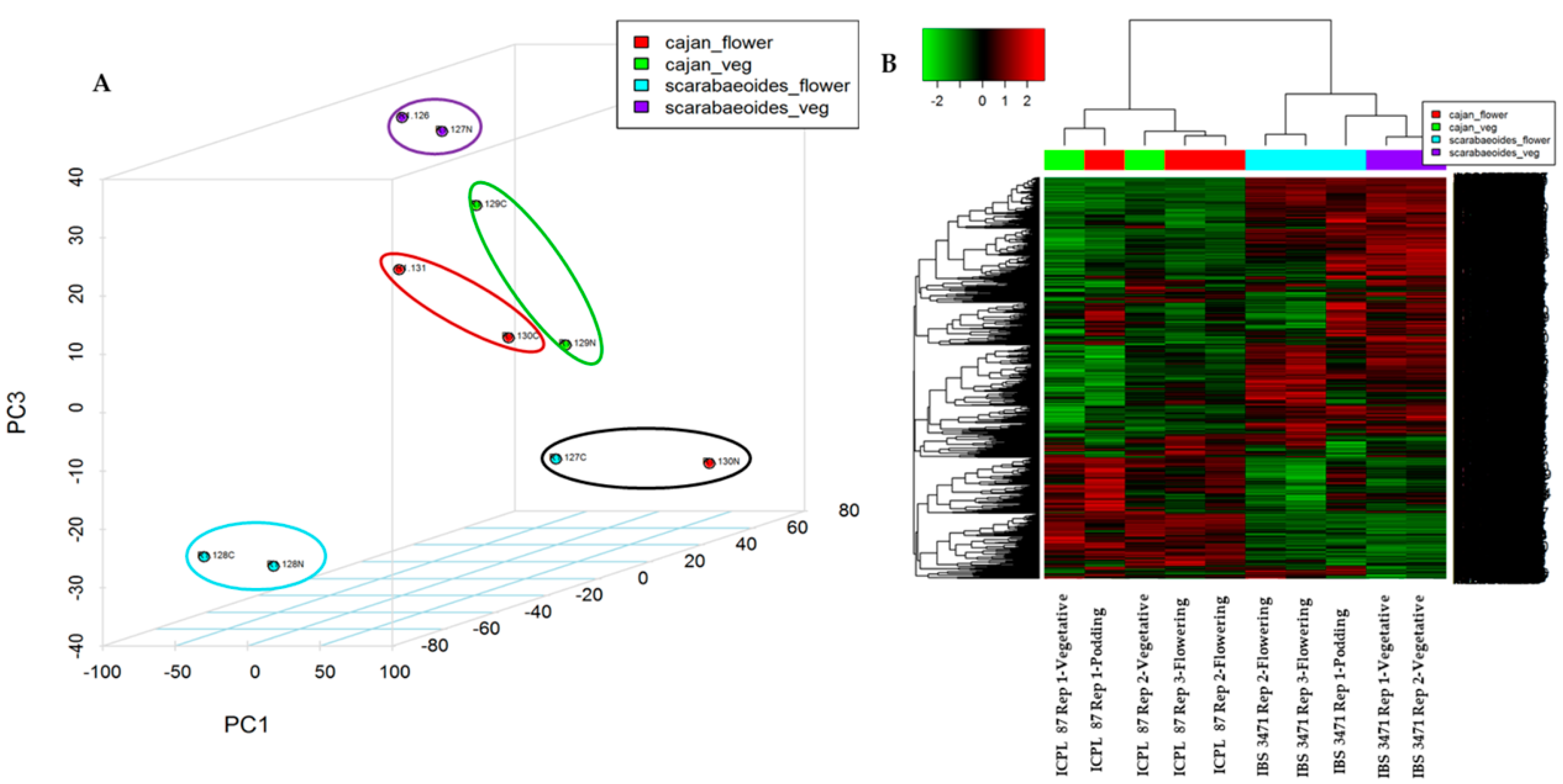
2.1. MS Analysis Revealed Differences in the Abundance of Hundreds of Proteins between H. armigera Tolerant and Susceptible Genotypes
2.2. PANTHER and KEGG GO Classification
2.3. Majority of the Defence-Related Proteins Were Involved in ROS and Signalling Pathways
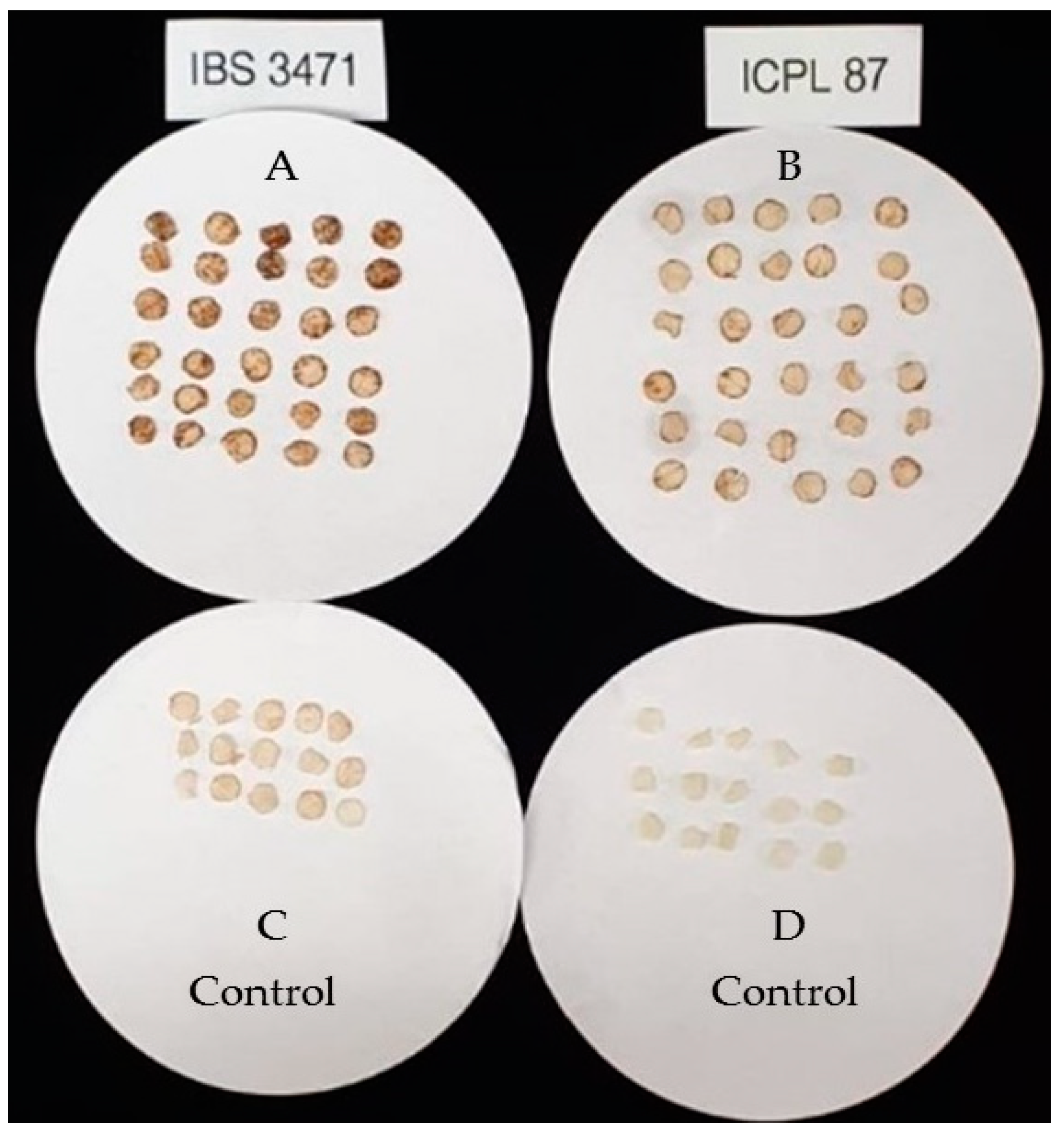
Oxidative Burst Examination of ROS Accumulation on Leaves
2.4. Quercetin-A Well-Known Insect Inhibitor Was More Abundant in IBS3471 Than in ICPL87 Leaves
Quercetin Antibiosis Effects on Helicoverpa Armigera Larval Growth and Development
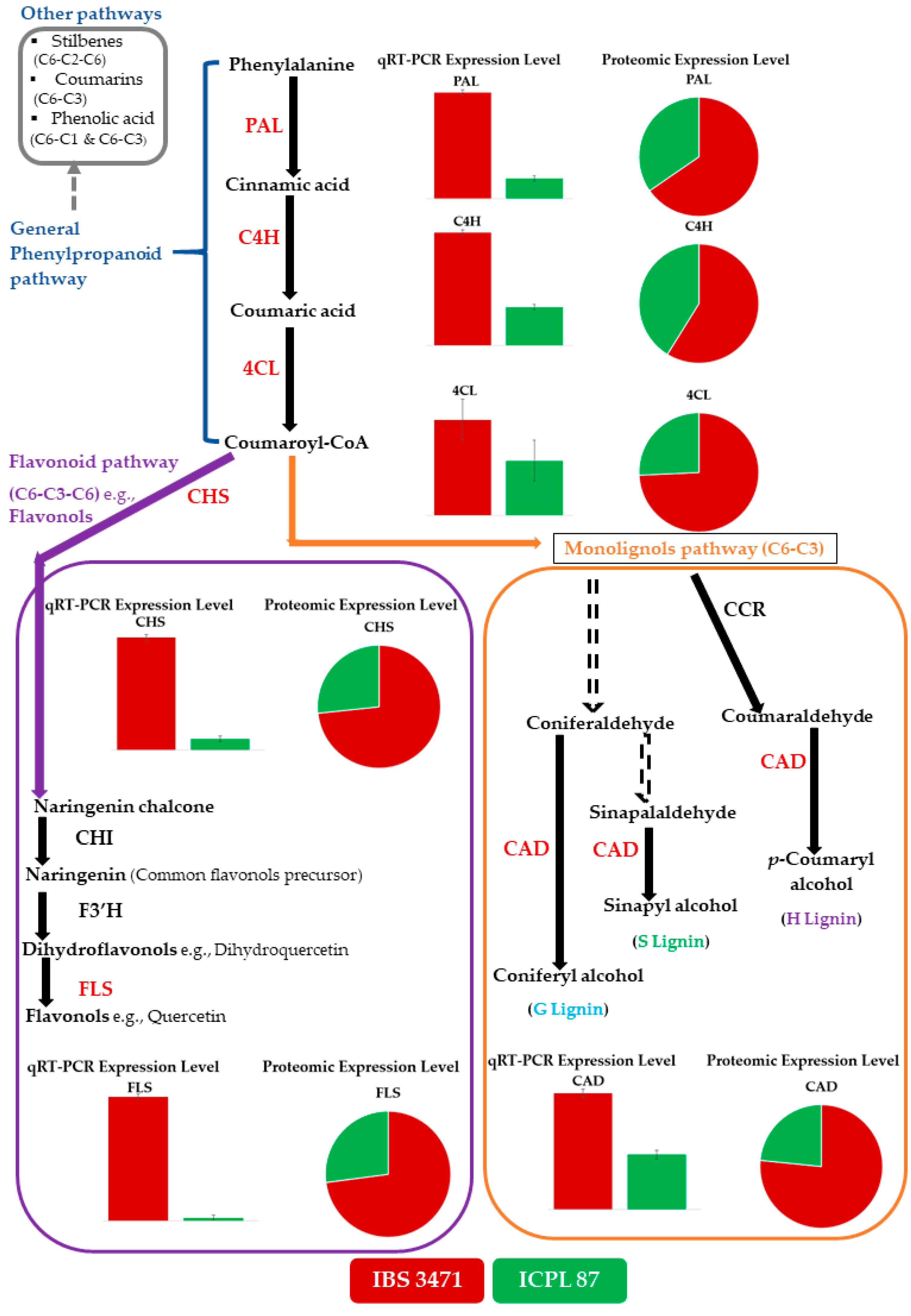
2.5. IBS 3471 Has More Quercetin and Monolignol Biosyntheses Enzymes Than ICPL 87
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. TMT-Based Protein Quantification
4.2.1. Protein Extraction, Reduction, Alkylation and Digestion
4.2.2. TMT Sample Labelling
4.2.3. Data Acquisition Using Nanoflow Liquid Chromatography-Electrospray Ionisation-Tandem Mass Spectrometry (NanoLC ESI MS/MS)
4.2.4. Protein Identification and Quantification
4.2.5. Gene Ontology (GO) Classification
4.3. DAB Staining Analysis
4.4. Identification of Flavonoids Compounds Using Liquid Chromatography-Mass Spectrometry (LCMS)
4.5. Biological Assays: Screening For H. Armigera Resistance (Antibiosis Mechanism) Using Artificial Diet Supplemented with Different Quercetin Concentrations Bioassay
4.6. Confirmatory and Validation Analysis of Protein Expression Using qRT-PCR Analysis
4.7. Data and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hillocks, R.J.; Minja, E.; Mwaga, A.; Nahdy, M.S.; Subrahmanyam, P. Diseases and pests of pigeonpea in eastern Africa: A review. Int. J. Pest Manag. 2000, 46, 7–18. [Google Scholar] [CrossRef]
- FAOSTAT. 2018. Available online: http://www.fao.org/faostat/en/ (accessed on 30 September 2020).
- Choudhary, A.K.; Raje, R.S.; Datta, S.; Sultana, R.; Ontagodi, T. Conventional and Molecular Approaches towards Genetic Improvement in Pigeonpea for Insects Resistance. Am. J. Plant Sci. 2013, 4, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Kriticos, D.J.; Ota, N.; Hutchison, W.D.; Beddow, J.; Walsh, T.; Tay, W.T.; Borchert, D.M.; Paula-Moraes, S.V.; Czepak, C.; Zalucki, M.P. Correction: The Potential Distribution of Invading Helicoverpa armigera in North America: Is It Just a Matter of Time? PLoS ONE 2015, 10, e0133224. [Google Scholar] [CrossRef]
- Tossou, E.; Tepa-Yotto, G.; Kpindou, O.K.D.; Sandeu, R.; Datinon, B.; Zeukeng, F.; Akoton, R.; Tchigossou, G.M.; Djègbè, I.; Vontas, J.; et al. Susceptibility Profiles of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) to Deltamethrin Reveal a Contrast between the Northern and the Southern Benin. Int. J. Environ. Res. Public Health 2019, 16, 1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohra, A.; Mallikarjuna, N.; Saxena, K.; Upadhyaya, H.; Vales, I.; Varshney, R. Harnessing the potential of crop wild relatives through genomics tools for pigeonpea improvement. J. Plant Biol. 2010, 37, 83–98. [Google Scholar]
- Saxena, K.B. Genetic Improvement of Pigeon Pea—A Review. Trop. Plant Biol. 2008, 1, 159–178. [Google Scholar] [CrossRef]
- Sharma, H. Crop protection compendium: Helicoverpa armigera. In Electron. Compend. Crop Prot; CAB International: Wallingford, UK, 2001. [Google Scholar]
- Howe, G.A.; Jander, G. Plant Immunity to Insect Herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [Green Version]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.; Villarroel, C.A.; Ataide, L.M.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant Defense against Insect Herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wink, M. Plant breeding: Importance of plant secondary metabolites for protection against pathogens and herbivores. Theor. Appl. Genet. 1988, 75, 225–233. [Google Scholar] [CrossRef]
- Chen, C.; Han, P.; Yan, W.; Wang, S.; Shi, X.; Zhou, X.; Desneux, N.; Gao, X. Uptake of quercetin reduces larval sensitivity to lambda-cyhalothrin in Helicoverpa armigera. J. Pest Sci. 2017, 91, 919–926. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Thompson, A.; Schäfer, J.; Kuhn, K.; Kienle, S.; Schwarz, J.; Schmidt, G.; Neumann, A.T.; Hamon, C. Tandem Mass Tags: A Novel Quantification Strategy for Comparative Analysis of Complex Protein Mixtures by MS/MS. Anal. Chem. 2003, 75, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Ngugi-Dawit, A.; Hoang, T.M.L.; Williams, B.; Higgins, T.J.; Mundree, S.G. A Wild Cajanus scarabaeoides (L.), Thouars, IBS 3471, for Improved Insect-Resistance in Cultivated Pigeonpea. Agronomy 2020, 10, 517. [Google Scholar] [CrossRef] [Green Version]
- Kuzina, V.; Ekstrøm, C.T.; Andersen, S.B.; Nielsen, J.K.; Olsen, C.E.; Bak, S. Identification of Defense Compounds in Barbarea vulgaris against the Herbivore Phyllotreta nemorum by an Ecometabolomic Approach. Plant Physiol. 2009, 151, 1977–1990. [Google Scholar] [CrossRef] [Green Version]
- Schuman, M.C.; Baldwin, I.T. The Layers of Plant Responses to Insect Herbivores. Annu. Rev. Entomol. 2016, 61, 373–394. [Google Scholar] [CrossRef]
- Njaci, I.; Ngugi-Dawit, A.; Oduor, R.O.; Kago, L.; Williams, B.; Hoang, L.T.M.; Mundree, S.G.; Ghimire, S.R. Comparative Analysis Delineates the Transcriptional Resistance Mechanisms for Pod Borer Resistance in the Pigeonpea Wild Relative Cajanus scarabaeoides (L.) Thouars. Int. J. Mol. Sci. 2020, 22, 309. [Google Scholar] [CrossRef]
- Rathinam, M.; Mishra, P.; Vasudevan, M.; Budhwar, R.; Mahato, A.; Prabha, A.L.; Singh, N.K.; Rao, U.; Sreevathsa, R. Comparative transcriptome analysis of pigeonpea, Cajanus cajan (L.) and one of its wild relatives Cajanus platycarpus (Benth.) Maesen. PLoS ONE 2019, 14, e0218731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathinam, M.; Mishra, P.; Mahato, A.K.; Singh, N.K.; Rao, U.; Sreevathsa, R. Comparative transcriptome analyses provide novel insights into the differential response of Pigeonpea (Cajanus cajan L.) and its wild relative (Cajanus platycarpus (Benth.) Maesen) to herbivory by Helicoverpa armigera (Hübner). Plant Mol. Biol. 2019, 101, 163–182. [Google Scholar] [CrossRef]
- Zhou, S.; Lou, Y.-R.; Tzin, V.; Jander, G. Alteration of plant primary metabolism in response to insect herbivory. Plant Physiol. 2015, 169, 1488–1498. [Google Scholar] [CrossRef] [Green Version]
- Rojas, C.M.; Esenthil-Kumar, M.; Etzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolwell, G.P.; Bindschedler, L.V.; Blee, K.A.; Butt, V.S.; Davies, D.R.; Gardner, S.L.; Gerrish, C.; Minibayeva, F. The apoplastic oxidative burst in response to biotic stress in plants: A three-component system. J. Exp. Bot. 2002, 53, 1367–1376. [Google Scholar] [CrossRef]
- Low, P.S.; Merida, J.R. The oxidative burst in plant defense: Function and signal transduction. Physiol. Plant. 1996, 96, 533–542. [Google Scholar] [CrossRef]
- Dangl, J.L.; Dietrich, R.A.; Thomas, H. Senescence and programmed cell death. In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists Press: Rockville, MD, USA, 2000; pp. 1044–1100. [Google Scholar]
- Jabs, T. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem. Pharmacol. 1999, 57, 231–245. [Google Scholar] [CrossRef]
- Collins, R.M.; Afzal, M.; Ward, D.A.; Prescott, M.C.; Sait, S.M.; Rees, H.H.; Tomsett, A.B. Differential Proteomic Analysis of Arabidopsis thaliana Genotypes Exhibiting Resistance or Susceptibility to the Insect Herbivore, Plutella xylostella. PLoS ONE 2010, 5, e10103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramputh, A.; Arnason, J.; Cass, L.; Simmonds, J. Reduced herbivory of the European corn borer (Ostrinia nubilalis) on corn transformed with germin, a wheat oxalate oxidase gene. Plant Sci. 2002, 162, 431–440. [Google Scholar] [CrossRef]
- Liu, Y.-H.; Offler, C.E.; Ruan, Y.-L. A simple, rapid, and reliable protocol to localize hydrogen peroxide in large plant organs by DAB-mediated tissue printing. Front. Plant Sci. 2014, 5, 745. [Google Scholar] [CrossRef] [PubMed]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Meitei, A.L.; Bhattacharjee, M.; Dhar, S.; Chowdhury, N.; Sharma, R.; Acharjee, S.; Sarmah, B.K. Activity of defense related enzymes and gene expression in pigeon pea (Cajanus cajan) due to feeding of Helicoverpa armigera larvae. J. Plant Interactions 2018, 13, 231–238. [Google Scholar] [CrossRef]
- Orozco-Cardenas, M.; Ryan, C.A. Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6553–6557. [Google Scholar] [CrossRef] [Green Version]
- Biała, W.; Jasiński, M. The Phenylpropanoid Case—It Is Transport That Matters. Front. Plant Sci. 2018, 9, 1610. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.-G.; Muchero, W. Regulation of Lignin Biosynthesis and Its Role in Growth-Defense Tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eynck, C.; Séguin-Swartz, G.; Clarke, W.E.; Parkin, I.A.P. Monolignol biosynthesis is associated with resistance to Sclerotinia sclerotiorum in Camelina sativa. Mol. Plant Pathol. 2012, 13, 887–899. [Google Scholar] [CrossRef] [PubMed]
- A Costa, M.; Collins, R.; Anterola, A.M.; Cochrane, F.C.; Davin, L.B.; Lewis, N.G. An in silico assessment of gene function and organization of the phenylpropanoid pathway metabolic networks in Arabidopsis thaliana and limitations thereof. Phytochemistry 2003, 64, 1097–1112. [Google Scholar] [CrossRef]
- Lauvergeat, V.; Lacomme, C.; Lacombe, E.; Lasserre, E.; Roby, D.; Grima-Pettenati, J. Two cinnamoyl-CoA reductase (CCR) genes from Arabidopsis thaliana are differentially expressed during development and in response to infection with pathogenic bacteria. Phytochemistry 2001, 57, 1187–1195. [Google Scholar] [CrossRef]
- Rathinam, M.; Roschitzki, B.; Grossmann, J.; Mishra, P.; Kunz, L.; Wolski, W.; Panse, C.; Tyagi, S.; Rao, U.; Schlapbach, R.; et al. Unraveling the proteomic changes involved in the resistance response of Cajanus platycarpus to herbivory by Helicoverpa armigera. Appl. Microbiol. Biotechnol. 2020, 104, 7603–7618. [Google Scholar] [CrossRef]
- Kaur, H.; Heinzel, N.; Schöttner, M.; Baldwin, I.T.; Gális, I. R2R3-NaMYB8 Regulates the Accumulation of Phenylpropanoid-Polyamine Conjugates, Which Are Essential for Local and Systemic Defense against Insect Herbivores in Nicotiana attenuata. Plant Physiol. 2010, 152, 1731–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Chen, Q.; Bu, Y.; Luo, R.; Hao, S.; Zhang, J.; Tian, J.; Yao, Y. Flavonoid Accumulation Plays an Important Role in the Rust Resistance of Malus Plant Leaves. Front. Plant Sci. 2017, 8, 1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nix, A.; A Paull, C.; Colgrave, M. The flavonoid profile of pigeonpea, Cajanus cajan: A review. SpringerPlus 2015, 4, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, P.; Stevenson, P.C.; Simmonds, M.S.J.; Sharma, H.C. Phenolic Compounds on the Pod-Surface of Pigeonpea, Cajanus cajan, Mediate Feeding Behavior of Helicoverpa armigera Larvae. J. Chem. Ecol. 2003, 29, 811–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goławska, S.; Sprawka, I.; Łukasik, I.; Golawski, A. Are naringenin and quercetin useful chemicals in pest-management strategies? J. Pest Sci. 2014, 87, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napal, G.N.D.; Palacios, S.M. Bioinsecticidal effect of the flavonoids pinocembrin and quercetin against Spodoptera frugiperda. J. Pest Sci. 2015, 88, 629–635. [Google Scholar] [CrossRef]
- Zu, Y.-G.; Fu, Y.-J.; Liu, W.; Hou, C.-L.; Kong, Y. Simultaneous Determination of Four Flavonoids in Pigeonpea [Cajanus cajan (L.) Millsp.] Leaves Using RP-LC-DAD. Chromatographia 2006, 63, 499–505. [Google Scholar] [CrossRef]
- Chen, S.; Elzaki, M.E.A.; Ding, C.; Li, Z.-F.; Wang, J.; Zeng, R.-S.; Song, Y.-Y. Plant allelochemicals affect tolerance of polyphagous lepidopteran pest Helicoverpa armigera (Hübner) against insecticides. Pestic. Biochem. Physiol. 2019, 154, 32–38. [Google Scholar] [CrossRef] [PubMed]
- War, A.R.; Paulraj, M.G.; Hussain, B.; Buhroo, A.A.; Ignacimuthu, S.; Sharma, H.C. Effect of plant secondary metabolites on legume pod borer, Helicoverpa armigera. J. Pest Sci. 2013, 86, 399–408. [Google Scholar] [CrossRef]
- Zhang, Y.-E.; Ma, H.-J.; Feng, D.-D.; Lai, X.-F.; Chen, Z.-M.; Xu, M.-Y.; Yu, Q.-Y.; Zhang, Z. Induction of Detoxification Enzymes by Quercetin in the Silkworm. J. Econ. Entomol. 2012, 105, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Khan, M.A.; Asaf, S.; Lee, I.-J.; Kim, K.-M. Overexpression of OsF3H modulates WBPH stress by alteration of phenylpropanoid pathway at a transcriptomic and metabolomic level in Oryza sativa. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- ICRISAT (International Crops Research Institute for the Semi-Arid Tropics). Pigeonpea Variety ICPL 87. In Plant Material De-Scription No.42; ICRISAT Center 93-208: Patancheru, Hyderabad, India, 1993; ISBN 92-9066-267-0. Order code: PME 042; Available online: http://oar.icrisat.org/id/eprint/565 (accessed on 28 July 2020).
- Wu, Y.; Mirzaei, M.; Pascovici, D.; Haynes, P.A.; Atwell, B.J. Proteomes of Leaf-Growing Zones in Rice Genotypes with Contrasting Drought Tolerance. Proteomics 2019, 19, e1800310. [Google Scholar] [CrossRef]
- Isaacson, T.; Damasceno, C.M.B.; Saravanan, R.S.; He, Y.; Catalá, C.; Saladié, M.; Rose, J.K.C. Sample extraction techniques for enhanced proteomic analysis of plant tissues. Nat. Protoc. 2006, 1, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Uniprot Data Base. Available online: http://www.uniprot.org/ (accessed on 10 October 2012).
- PANTHER. Classification System. Available online: http://www.pantherdb.org/ (accessed on 31 October 2019).
- KEGG. Available online: http://www.genome.jp/kegg/ (accessed on 10 January 2009).
- Daudi, A.; O’Brien, J.A. Detection of Hydrogen Peroxide by DAB Staining in Arabidopsis Leaves. Bio-Protocol 2012, 2, e263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi, N.; Mahernia, S.; Amanlou, M. Comparison of Different Methods in Quercetin Extraction from Leaves of Raphanus sativus L. Pharm. Sci. 2017, 23, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Pascovici, D.; Wu, J.X.; Chick, J.; Wu, Y.; Cooke, B.; Haynes, P.; Molloy, M.P. TMT One-Stop Shop: From Reliable Sample Preparation to Computational Analysis Platform. Adv. Struct. Saf. Stud. 2016, 1549, 45–66. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt ID | Protein Description | Proteomic LOG2FC | ANOVA (p < 0.05) | Role |
|---|---|---|---|---|
| Antioxidant, ROS Scavenger & Detoxification Enzymes | ||||
| A0A151R1K6 | Putative glutathione S-transferase | 5.674 | 0.000 | Antioxidant/ ROS scavenger/detoxifying enzyme |
| A0A151RS05 | Cytochrome P450 | 3.688 | 0.000 | Biosynthesis of primary & secondary metabolites |
| A0A151QVW9 | Peroxidase | 2.591 | 0.010 | Response to stress/Antioxidant |
| A0A151TXJ4 | Superoxide dismutase [CuZn] | 2.567 | 0.002 | Scavenging of O2 radicals/superoxide anions |
| Secondary Metabolite | ||||
| A0A151QU72 | Anthocyanidin 5,3-O-glucosyltransferase | 2.898 | 0.001 | Anthocyanin biosynthesis pathway |
| A0A151RR63 | Isoflavone-7-O-methyltransferase | 2.496 | 0.001 | Phytoalexin biosynthesis |
| A0A151RRX4 | Dirigent protein | 2.007 | 0.010 | Monolignol pathway |
| Signalling | ||||
| A0A151U2F2 | Calcium-dependent protein kinase | 2.714 | 0.004 | Signal transduction pathways |
| A0A151U2V3 | Hydroxyethylthiazole kinase | 2.538 | 0.002 | Cofactor biosynthesis pathway |
| A0A151TPW7 | Annexin | 1.810 | 0.003 | Calcium-dependent membrane-binding proteins |
| A0A151SDR5 | Jasmonate O-methyltransferase | 3.156 | 0.000 | Signalling & defence-related processes |
| Anti-Nutritional Factors | ||||
| A0A151SAI2 | Lysosomal Pro-X carboxypeptidase | 3.160 | 0.009 | Herbivore defensive protein |
| A0A151RBF5 | Subtilisin-like protease | 2.256 | 0.000 | Serine proteases (development & signalling) |
| A0A151QZF6 | Cysteine proteinase inhibitor | 2.343 | 0.001 | Herbivore defensive protein |
| Carbohydrate Metabolism | ||||
| A0A151QPT9 | Acidic endochitinase | 3.824 | 0.009 | Response to biotic & abiotic stresses |
| A0A151TII8 | Lipase | 2.999 | 0.002 | Synthesis of signalling defence lipids & precursors |
| Phytohormone | ||||
| A0A151RG77 | Cytokinin-Oglucosyltransferase | 3.730 | 0.000 | Cytokinin (biotic & abiotic stress responses) |
| A0A151RKG5 | 1-aminocyclopropane-1-carboxylate oxidase | 1.812 | 0.001 | Ethylene (development & stress responses) |
| Flavonoid Class | Possible Compounds | Ion Formula (Negative Ion Mode) | Mass-To-Charge (M/Z) | Retention Time (Minutes) | Peak Area | Fold Change (IBS 3471/ICPL 87) | |
|---|---|---|---|---|---|---|---|
| IBS 3471 | ICPL 87 | ||||||
| Flavonols | Quercetin | C15H9O7− | 301.0354 | 62.5 | 1,774,965 | 952,200 | 2 |
| Isorhamnetin | C16H11O7− | 315.051 | 64.6 | 450,240 | 114,517 | 4 | |
| 66.6 | 146,742 | 1,356,558 | 0 | ||||
| Flavanones | Naringenin | C15H11O5− | 271.0612 | 55.9 | 273,446 | 35,715 | 8 |
| 63.1 | 180,120 | 33,276 | 5 | ||||
| 64.6 | 27,846 | 6202 | 4 | ||||
| Flavones | Vitexin or Isovitexin | C21H19O10− | 431.0984 | 55.8 | 61,411,155 | 61,736,130 | 1 |
| Luteolin | C15H9O6− | 285.0405 | 63.8 | 3,130,076 | 523,375 | 6 | |
| 65.8 | 1,687,625 | 145,539 | 12 | ||||
| Sample Name | Replication | Growth Stage | TMT Reagent Labelling of Peptide |
|---|---|---|---|
| IBS 3471 1 (Cajanus scarabaeoides) | Rep 1 | Vegetative stage | 126 |
| Rep 2 | 127N | ||
| Rep 1 | Flowering/podding stage | 127C | |
| Rep 2 | 128N | ||
| Rep 3 | 128C | ||
| ICPL 87 2 (Cajanus cajan) | Rep 1 | Vegetative stage | 129N |
| Rep 2 | 129C | ||
| Rep 1 | Flowering/podding stage | 130N | |
| Rep 2 | 130C | ||
| Rep 3 | 131 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngugi-Dawit, A.; Njaci, I.; Higgins, T.J.V.; Williams, B.; Ghimire, S.R.; Mundree, S.G.; Hoang, L.T.M. Comparative TMT Proteomic Analysis Unveils Unique Insights into Helicoverpa armigera (Hübner) Resistance in Cajanus scarabaeoides (L.) Thouars. Int. J. Mol. Sci. 2021, 22, 5941. https://doi.org/10.3390/ijms22115941
Ngugi-Dawit A, Njaci I, Higgins TJV, Williams B, Ghimire SR, Mundree SG, Hoang LTM. Comparative TMT Proteomic Analysis Unveils Unique Insights into Helicoverpa armigera (Hübner) Resistance in Cajanus scarabaeoides (L.) Thouars. International Journal of Molecular Sciences. 2021; 22(11):5941. https://doi.org/10.3390/ijms22115941
Chicago/Turabian StyleNgugi-Dawit, Abigail, Isaac Njaci, Thomas J. V. Higgins, Brett Williams, Sita R. Ghimire, Sagadevan G. Mundree, and Linh Thi My Hoang. 2021. "Comparative TMT Proteomic Analysis Unveils Unique Insights into Helicoverpa armigera (Hübner) Resistance in Cajanus scarabaeoides (L.) Thouars" International Journal of Molecular Sciences 22, no. 11: 5941. https://doi.org/10.3390/ijms22115941