
Ecological Adaptations of Gut Microbiota Members and Their Consequences for Use as a New Generation of Probiotics

Abstract
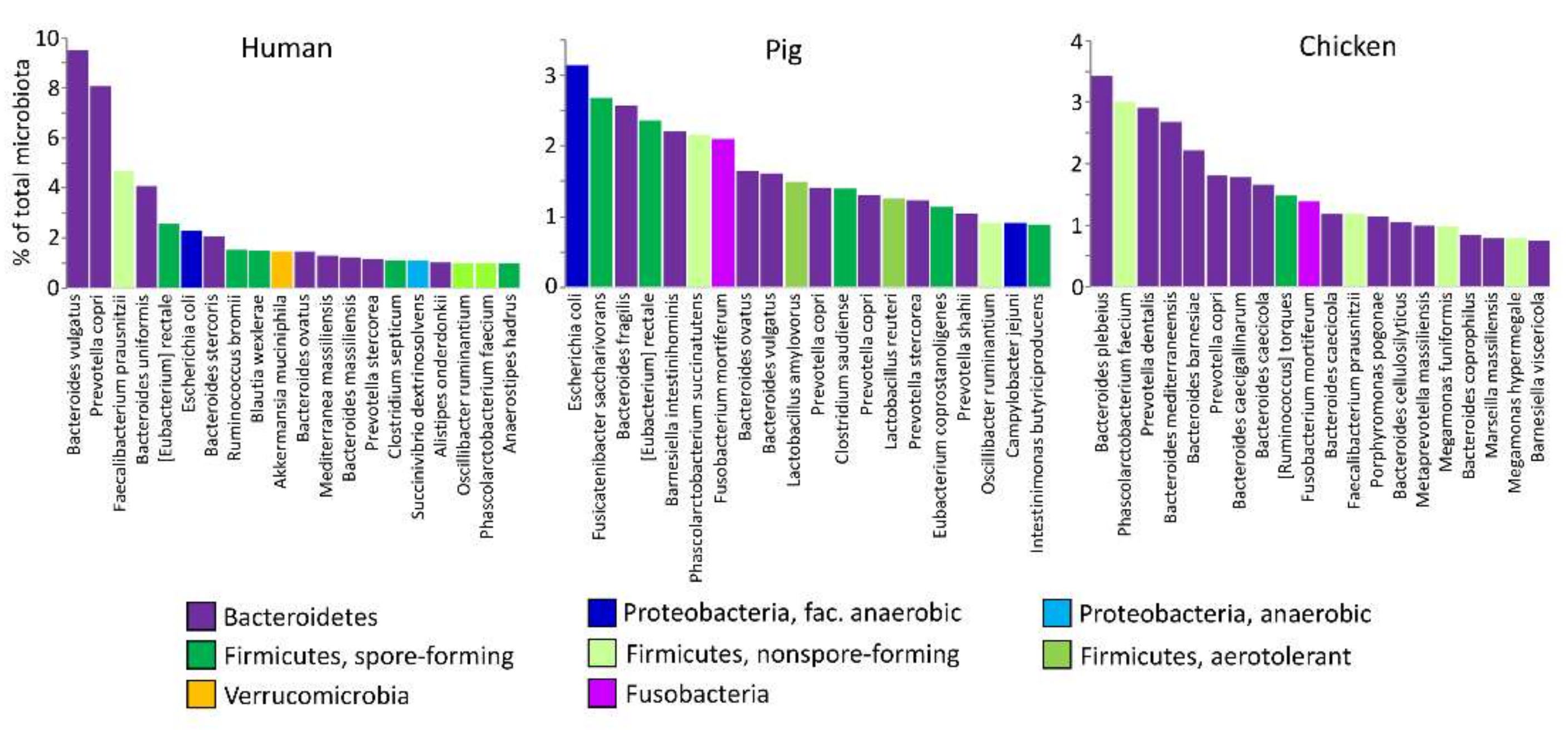
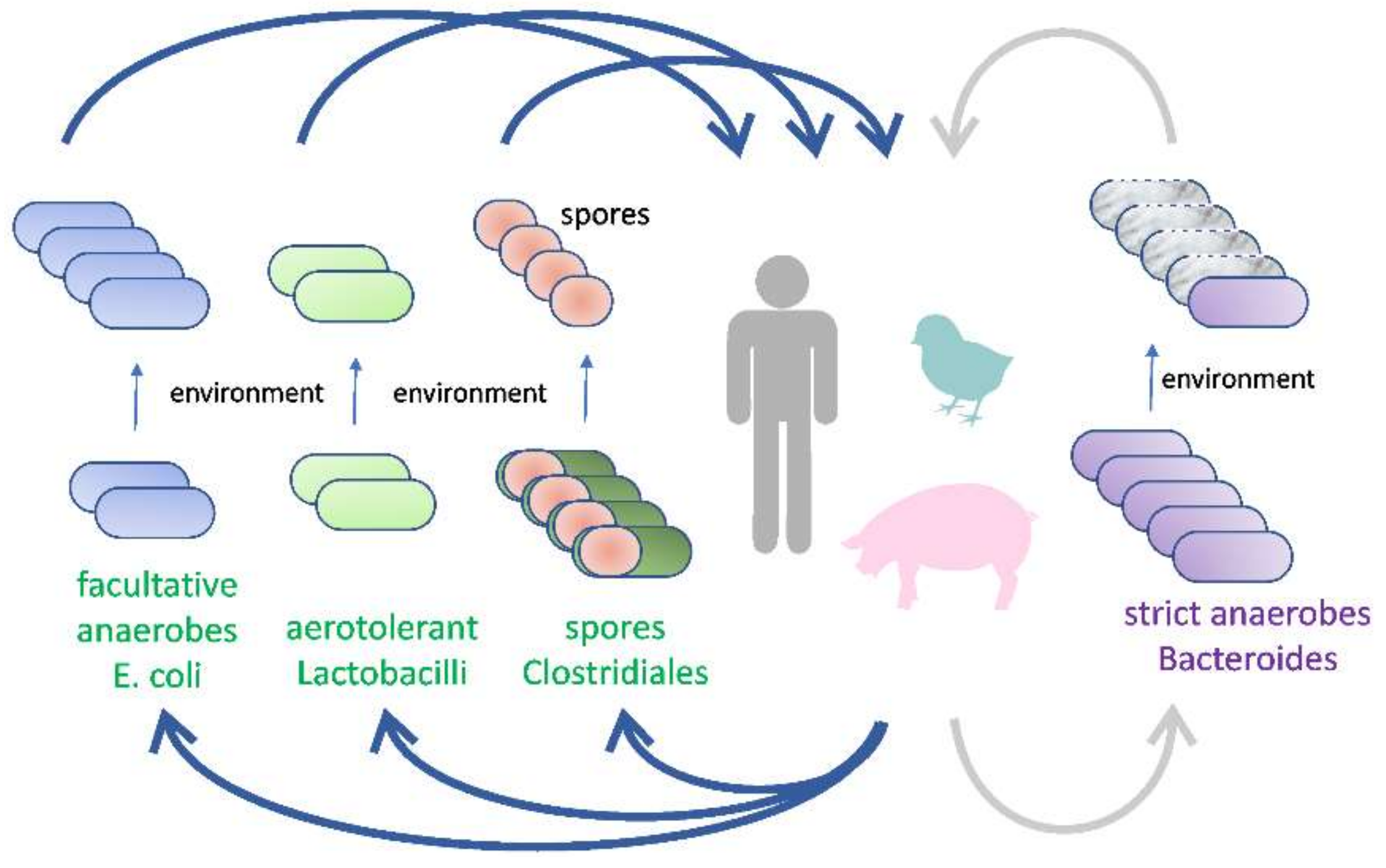
:1. Introduction
2. Novel Types of Probiotics from Gut Microbiota
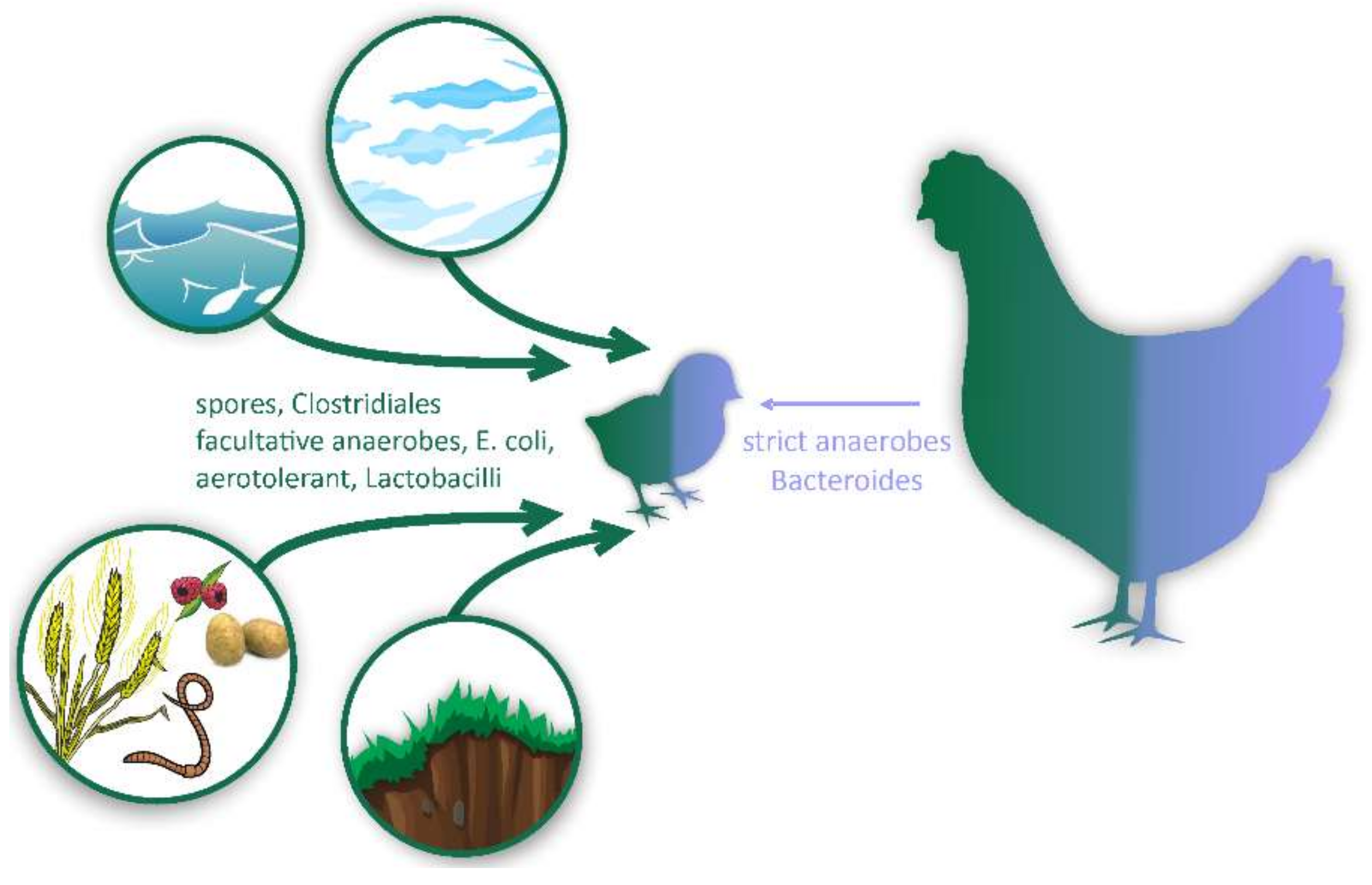
2.1. Aerotolerant Bacteria Commonly Associated with Food and Feed
2.2. Facultative Anaerobes
2.3. Spore-Forming Firmicutes
2.4. Non-Spore-Forming Gut Anaerobes
3. Final Remarks
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kubasova, T.; Davidova-Gerzova, L.; Babak, V.; Cejkova, D.; Montagne, L.; Le-Floc’H, N.; Rychlik, I. Effects of host genetics and environmental conditions on fecal microbiota composition of pigs. PLoS ONE 2018, 13, e0201901. [Google Scholar] [CrossRef] [PubMed]
- Videnska, P.; Sedlar, K.; Lukac, M.; Faldynova, M.; Gerzova, L.; Cejkova, D.; Sisak, F.; Rychlik, I. Succession and Replacement of Bacterial Populations in the Caecum of Egg Laying Hens over Their Whole Life. PLoS ONE 2014, 9, e115142. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liang, S.; Xia, Z.; Qu, J.; Liu, C.; Yang, H.; Wang, J.; Madsen, L.; Hou, Y.; Li, J.; et al. Establishment of a Macaca fascicularis gut microbiome gene catalog and comparison with the human, pig, and mouse gut microbiomes. GigaScience 2018, 7, 100. [Google Scholar] [CrossRef]
- Donnell, M.M.O.; Harris, H.M.B.; Ross, R.P.; O’Toole, P.W. Core fecal microbiota of domesticated herbivorous ruminant, hindgut fermenters, and monogastric animals. Microbiologyopen 2017, 6, e00509. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef] [PubMed]
- Rezazadeh, L.; Alipour, B.; Jafarabadi, M.A.; Behrooz, M.; Gargari, B.P. Daily consumption effects of probiotic yogurt containing Lactobacillus acidophilus La5 and Bifidobacterium lactis Bb12 on oxidative stress in metabolic syndrome patients. Clin. Nutr. ESPEN 2021, 41, 136–142. [Google Scholar] [CrossRef]
- Boyen, F.; Haesebrouck, F.; Vanparys, A.; Volf, J.; Mahu, M.; Van Immerseel, F.; Rychlik, I.; Dewulf, J.; Ducatelle, R.; Pasmans, F. Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Veter. Microbiol. 2008, 132, 319–327. [Google Scholar] [CrossRef]
- Van Immerseel, F.; De Buck, J.; Boyen, F.; Bohez, L.; Pasmans, F.; Volf, J.; Sevcik, M.; Rychlik, I.; Haesebrouck, F.; Ducatelle, R. Medium-Chain Fatty Acids Decrease Colonization and Invasion through hilA Suppression Shortly after Infection of Chickens with Salmonella enterica Serovar Enteritidis. Appl. Environ. Microbiol. 2004, 70, 3582–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebbia, S.; Lamberti, C.; Bianco, G.L.; Cirrincione, S.; Laroute, V.; Cocaign-Bousquet, M.; Cavallarin, L.; Giuffrida, M.G.; Pessione, E. Antimicrobial Potential of Food Lactic Acid Bacteria: Bioactive Peptide Decrypting from Caseins and Bacteriocin Production. Microorganisms 2020, 9, 65. [Google Scholar] [CrossRef]
- Derrien, M.; Vlieg, J.E.V.H. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahbazi, R.; Sharifzad, F.; Bagheri, R.; Alsadi, N.; Yasavoli-Sharahi, H.; Matar, C. Anti-Inflammatory and Immunomodulatory Properties of Fermented Plant Foods. Nutrients 2021, 13, 1516. [Google Scholar] [CrossRef]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Karaffová, V.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Ševčíková, Z.; Levkut, M.; Herich, R.; Revajová, V. The Probiotic Lactobacillus fermentum Biocenol CCM 7514 Moderates Campylobacter jejuni-Induced Body Weight Impairment by Improving Gut Morphometry and Regulating Cecal Cytokine Abundance in Broiler Chickens. Animals 2021, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.; Baptista, A.A.S.; Valdiviezo, M.J.; Justino, L.; Menck-Costa, M.F.; Ferraz, C.R.; da Gloria, E.M.; Verri, W.A.; Bracarense, A.P.F. Lactobacillus spp. reduces morphological changes and oxidative stress induced by deoxynivalenol on the intestine and liver of broilers. Toxicon 2020, 185, 203–212. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef] [PubMed]
- Crhanova, M.; Karasova, D.; Juricova, H.; Matiasovicova, J.; Jahodarova, E.; Kubasova, T.; Seidlerova, Z.; Cizek, A.; Rychlik, I. Systematic Culturomics Shows that Half of Chicken Caecal Microbiota Members can be Grown in Vitro Except for Two Lineages of Clostridiales and a Single Lineage of Bacteroidetes. Microorganisms 2019, 7, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, L.; Nielsen, B.; Boll, E.; Skjøt-Rasmussen, L.; Wellejus, A.; Jørgensen, L.; Lauridsen, C.; Canibe, N. Functional in vitro screening of probiotic strains for inoculation of piglets as a prophylactic measure towards Enterotoxigenic Escherichia coli infection. J. Microbiol. Methods 2021, 180, 106126. [Google Scholar] [CrossRef]
- Wang, J.; Bai, X.; Peng, C.; Yu, Z.; Li, B.; Zhang, W.; Sun, Z.; Zhang, H. Fermented milk containing Lactobacillus casei Zhang and Bifidobacterium animalis ssp. lactis V9 alleviated constipation symptoms through regulation of intestinal microbiota, inflammation, and metabolic pathways. J. Dairy Sci. 2020, 103, 11025–11038. [Google Scholar] [CrossRef] [PubMed]
- Di Napoli, M.; Di Luccia, B.; Vitiello, G.; D’Errico, G.; Carpentieri, A.; Pezzella, A.; Pizzo, E.; Notomista, E.; Varcamonti, M.; Zanfardino, A. Characterisation of EFV12 a bio-active small peptide produced by the human intestinal isolate Lactobacillus gasseri SF1109. Benef. Microbes 2020, 11, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gutierrez, E.; O’Connor, P.M.; Colquhoun, I.J.; Vior, N.M.; Rodríguez, J.M.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Production of multiple bacteriocins, including the novel bacteriocin gassericin M, by Lactobacillus gasseri LM19, a strain isolated from human milk. Appl. Microbiol. Biotechnol. 2020, 104, 3869–3884. [Google Scholar] [CrossRef] [Green Version]
- Bujalance, C.; Moreno, E.; Jiménez-Valera, M.; Ruiz-Bravo, A. A probiotic strain of Lactobacillus plantarum stimulates lymphocyte responses in immunologically intact and immunocompromised mice. Int. J. Food Microbiol. 2007, 113, 28–34. [Google Scholar] [CrossRef]
- Hrdý, J.; Alard, J.; Couturier-Maillard, A.; Boulard, O.; Boutillier, D.; Delacre, M.; Lapadatescu, C.; Cesaro, A.; Blanc, P.; Pot, B.; et al. Lactobacillus reuteri 5454 and Bifidobacterium animalis ssp. lactis 5764 improve colitis while differentially impacting dendritic cells maturation and antimicrobial responses. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anania, C.; Di Marino, V.; Olivero, F.; De Canditiis, D.; Brindisi, G.; Iannilli, F.; De Castro, G.; Zicari, A.; Duse, M. Treatment with a Probiotic Mixture Containing Bifidobacterium animalis Subsp. Lactis BB12 and Enterococcus faecium L3 for the Prevention of Allergic Rhinitis Symptoms in Children: A Randomized Controlled Trial. Nutrients 2021, 13, 1315. [Google Scholar] [CrossRef]
- Kubasova, T.; Davidova-Gerzova, L.; Merlot, E.; Medvecky, M.; Polansky, O.; Gardan-Salmon, D.; Quesnel, H.; Rychlik, I. Housing Systems Influence Gut Microbiota Composition of Sows but Not of Their Piglets. PLoS ONE 2017, 12, e0170051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Pokorna, A.; Cizek, A.; et al. Contact with adult hen affects development of caecal microbiota in newly hatched chicks. PLoS ONE 2019, 14, e0212446. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Zhu, A.; Benes, V.; Costea, P.I.; Hercog, R.; Hildebrand, F.; Huerta-Cepas, J.; Nieuwdorp, M.; Salojärvi, J.; Voigt, A.Y.; et al. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 2016, 352, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Nii, T.; Kakuya, H.; Isobe, N.; Yoshimura, Y. Lactobacillus reuteri Enhances the Mucosal Barrier Function against Heat-killed Salmonella Typhimurium in the Intestine of Broiler Chicks. J. Poult. Sci. 2020, 57, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Sorrento, A.; Castillejos, L.; López-Colom, P.; Cifuentes-Orjuela, G.; Rodríguez-Palmero, M.; Moreno-Muñoz, J.A.; Luise, D.; Trevisi, P.; Martín-Orúe, S.M. Effects of the Administration of Bifidobacterium longum subsp. infantis CECT 7210 and Lactobacillus rhamnosus HN001 and Their Synbiotic Combination with Galacto-Oligosaccharides Against Enterotoxigenic Escherichia coli F4 in an Early Weaned Piglet Model. Front. Microbiol. 2021, 12, 642549. [Google Scholar] [CrossRef]
- Spreckels, J.; Wejryd, E.; Marchini, G.; Jonsson, B.; de Vries, D.; Jenmalm, M.; Landberg, E.; Sverremark-Ekström, E.; Martí, M.; Abrahamsson, T. Lactobacillus reuteri Colonisation of Extremely Preterm Infants in a Randomised Placebo-Controlled Trial. Microorganisms 2021, 9, 915. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liang, S.; Guo, F.; Ren, Z.; Yang, X.; Long, F. Gut microbiota mediates the protective role of Lactobacillus plantarum in ameliorating deoxynivalenol-induced apoptosis and intestinal inflammation of broiler chickens. Poult. Sci. 2020, 99, 2395–2406. [Google Scholar] [CrossRef]
- Geng, T.; Su, S.; Sun, K.; Zhao, L.; Zhao, Y.; Bao, N.; Pan, L.; Sun, H. Effects of feeding a Lactobacillus plantarum JL01 diet on caecal bacteria and metabolites of weaned piglets. Lett. Appl. Microbiol. 2021, 72, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Hofeld, B.C.; Puppala, V.K.; Tyagi, S.; Ahn, K.W.; Anger, A.; Jia, S.; Salzman, N.H.; Hessner, M.J.; Widlansky, M.E. Lactobacillus plantarum 299v probiotic supplementation in men with stable coronary artery disease suppresses systemic inflammation. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Sisak, F.; Babak, V.; et al. Gut Anaerobes Capable of Chicken Caecum Colonisation. Microorganisms 2019, 7, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Arnesen, L.P.S.; Fagerlund, A.; Granum, P.E. From soil to gut:Bacillus cereusand its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medvecky, M.; Cejkova, D.; Polansky, O.; Karasova, D.; Kubasova, T.; Cizek, A.; Rychlik, I. Whole genome sequencing and function prediction of 133 gut anaerobes isolated from chicken caecum in pure cultures. BMC Genom. 2018, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, I. Composition and Function of Chicken Gut Microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Onizuka, S.; Mishima, R.; Nakayama, J. Cultural isolation of spore-forming bacteria in human feces using bile acids. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Videnska, P.; Faldynova, M.; Juricova, H.; Babak, V.; Sisak, F.; Havlickova, H.; Rychlik, I. Chicken faecal microbiota and disturbances induced by single or repeated therapy with tetracycline and streptomycin. BMC Veter. Res. 2013, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, B.; Kim, S.W.; Kwon, Y.M. Characterization of Microbiota Associated with Digesta and Mucosa in Different Regions of Gastrointestinal Tract of Nursery Pigs. Int. J. Mol. Sci. 2019, 20, 1630. [Google Scholar] [CrossRef] [Green Version]
- Slifierz, M.J.; Friendship, R.M.; Weese, J.S. Longitudinal study of the early-life fecal and nasal microbiotas of the domestic pig. BMC Microbiol. 2015, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Reid, G.; Gaudier, E.; Guarner, F.; Huffnagle, G.B.; Macklaim, J.M.; Munoz, A.M.; Martini, M.; Ringel-Kulka, T.; Sartor, B.R.; Unal, R.R.; et al. Responders and non-responders to probiotic interventions. Gut Microbes 2010, 1, 200–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, H.; Starrenburg, M.J.; Molenaar, D.; Kleerebezem, M.; Vlieg, J.E.V.H. Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution. Genome Res. 2011, 22, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Chee, W.J.Y.; Chew, S.Y.; Than, L.T.L. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microb. Cell Factories 2020, 19, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Bucher, M.G.; Zwirzitz, B.; Oladeinde, A.; Cook, K.; Plymel, C.; Zock, G.; Lakin, S.; Aggrey, S.E.; Ritz, C.; Looft, T.; et al. Reused poultry litter microbiome with competitive exclusion potential against Salmonella Heidelberg. J. Environ. Qual. 2020, 49, 869–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, H.P.; Neville, B.A.; Forster, S.C.; Lawley, T.D. Transmission of the gut microbiota: Spreading of health. Nat. Rev. Genet. 2017, 15, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Francavilla, R.; De Angelis, M.; Rizzello, C.G.; Cavallo, N.; Bello, F.D.; Gobbetti, M. Selected Probiotic Lactobacilli Have the Capacity to Hydrolyze Gluten Peptides during Simulated Gastrointestinal Digestion. Appl. Environ. Microbiol. 2017, 83, e00376-17. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Platt, G. Fermented foods—A world perspective. Food Res. Int. 1994, 27, 253–257. [Google Scholar] [CrossRef]
- Kearney, S.M.; Gibbons, S.M.; Poyet, M.; Gurry, T.; Bullock, K.; Allegretti, J.R.; Clish, C.B.; Alm, E.J. Endospores and other lysis-resistant bacteria comprise a widely shared core community within the human microbiota. ISME J. 2018, 12, 2403–2416. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, J.; Larsson, P.; Singh, B.; Pettersson, F.; Islam, N.M.; Sarkar, S.N.; Dasgupta, S.; Kirsebom, L.A. Sporulation in mycobacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 10781–10786. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Kiewiet, M.B.G.; Logtenberg, M.J.; Groeneveld, A.; Nauta, A.; Schols, H.A.; Walvoort, M.T.C.; Harmsen, H.J.M.; De Vos, P. Effects of Different Human Milk Oligosaccharides on Growth of Bifidobacteria in Monoculture and Co-culture with Faecalibacterium prausnitzii. Front. Microbiol. 2020, 11, 569700. [Google Scholar] [CrossRef]
- Karasova, D.; Crhanova, M.; Babak, V.; Jerabek, M.; Brzobohaty, L.; Matesova, Z.; Rychlik, I. Development of piglet gut microbiota at the time of weaning influences development of postweaning diarrhea—A field study. Res. Veter. Sci. 2020, 135, 59–65. [Google Scholar] [CrossRef]
- Takakura, W.; Pimentel, M. Small Intestinal Bacterial Overgrowth and Irritable Bowel Syndrome—An Update. Front. Psychiatry 2020, 11, 664. [Google Scholar] [CrossRef]
- Cukrowska, B.; Lodínová-Žádníková, R.; Enders, C.; Sonnenborn, U.; Schulze, J.; Tlaskalová-Hogenová, H. Specific Proliferative and Antibody Responses of Premature Infants to Intestinal Colonization with Nonpathogenic Probiotic E. coli Strain Nissle 1917. Scand. J. Immunol. 2002, 55, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Henker, J.; Laass, M.; Blokhin, B.M.; Bolbot, Y.K.; Maydannik, V.G.; Elze, M.; Wolff, C.; Schulze, J. The probiotic Escherichia coli strain Nissle 1917 (EcN) stops acute diarrhoea in infants and toddlers. Eur. J. Nucl. Med. Mol. Imaging 2007, 166, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faldynova, M.; Pravcova, M.; Sisak, F.; Havlickova, H.; Kolackova, I.; Cizek, A.; Karpiskova, R.; Rychlik, I. Evolution of Antibiotic Resistance in Salmonella enterica Serovar Typhimurium Strains Isolated in the Czech Republic between 1984 and 2002. Antimicrob. Agents Chemother. 2003, 47, 2002–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.-K.; Kim, W.-S.; Paik, H.-D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Mazanko, M.S.; Gorlov, I.; Prazdnova, E.V.; Makarenko, M.S.; Usatov, A.V.; Bren, A.B.; Chistyakov, V.A.; Tutelyan, A.V.; Komarova, Z.B.; Mosolova, N.I.; et al. Bacillus Probiotic Supplementations Improve Laying Performance, Egg Quality, Hatching of Laying Hens, and Sperm Quality of Roosters. Probiotics Antimicrob. Proteins 2018, 10, 367–373. [Google Scholar] [CrossRef]
- Poulsen, A.-S.R.; De Jonge, N.; Nielsen, J.L.; Højberg, O.; Lauridsen, C.; Cutting, S.M.; Canibe, N. Impact of Bacillus spp. spores and gentamicin on the gastrointestinal microbiota of suckling and newly weaned piglets. PLoS ONE 2018, 13, e0207382. [Google Scholar] [CrossRef] [Green Version]
- Von Stetten, F. Climatic influence on mesophilic Bacillus cereus and psychrotolerant Bacillus weihenstephanensis populations in tropical, temperate and alpine soil. Environ. Microbiol. 1999, 1, 503–515. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nat. Cell Biol. 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Nayfach, S.; Rodriguez-Mueller, B.; Garud, N.; Pollard, K.S. An integrated metagenomics pipeline for strain profiling reveals novel patterns of bacterial transmission and biogeography. Genome Res. 2016, 26, 1612–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avershina, E.; Larsen, M.G.; Aspholm, M.; Lindbäck, T.; Storrø, O.; Øien, T.; Johnsen, R.; Rudi, K. Culture dependent and independent analyses suggest a low level of sharing of endospore-forming species between mothers and their children. Sci. Rep. 2020, 10, 1832. [Google Scholar] [CrossRef] [PubMed]
- Browne, H.P.; Forster, S.C.; Anonye, B.O.; Kumar, N.; Neville, B.A.; Stares, M.D.; Goulding, D.; Lawley, T.D. Culturing of ‘unculturable’ human microbiota reveals novel taxa and extensive sporulation. Nat. Cell Biol. 2016, 533, 543–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forster, S.C.; Kumar, N.; Anonye, B.O.; Almeida, A.; Viciani, E.; Stares, M.D.; Dunn, M.; Mkandawire, T.; Zhu, A.; Shao, Y.; et al. A human gut bacterial genome and culture collection for improved metagenomic analyses. Nat. Biotechnol. 2019, 37, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y. Genome Diversity of Spore-Forming Firmicutes. Microbiol. Spectr. 2013, 1, TBS-0015-2012. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.; Poppleton, D.; Klingl, A.; Criscuolo, A.; Dupuy, B.; Brochier-Armanet, C.; Beloin, C.; Gribaldo, S. Phylogenomic analysis supports the ancestral presence of LPS-outer membranes in the Firmicutes. eLife 2016, 5, e14589. [Google Scholar] [CrossRef]
- Kollarcikova, M.; Faldynova, M.; Matiasovicova, J.; Jahodarova, E.; Kubasova, T.; Seidlerova, Z.; Babak, V.; Videnska, P.; Cizek, A.; Rychlik, I. Different Bacteroides Species Colonise Human and Chicken Intestinal Tract. Microorganisms 2020, 8, 1483. [Google Scholar] [CrossRef]
- Browne, H.P.; Almeida, A.; Kumar, N.; Vervier, K.; Adoum, A.T.; Viciani, E.; Dawson, N.J.R.; Forster, S.C.; Cormie, C.; Goulding, D.; et al. Host adaptation in gut Firmicutes is associated with sporulation loss and altered colonisa-tion patterns. bioRxiv 2021. [Google Scholar] [CrossRef]
- Van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; De Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.W.M.; Tijssen, J.G.P.; et al. Duodenal Infusion of Donor Feces for Recurrent Clostridium difficile. New Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Varmuzova, K.; Kubasova, T.; Davidova-Gerzova, L.; Sisak, F.; Havlickova, H.; Sebkova, A.; Faldynova, M.; Rychlik, I. Composition of Gut Microbiota Influences Resistance of Newly Hatched Chickens to Salmonella Enteritidis Infection. Front. Microbiol. 2016, 7, 957. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F.; Filion, K.B.; et al. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by Caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Form of Air Resistance | Ecological Classification | Major Taxa from Gut Microbiota | Reference |
|---|---|---|---|
| Aerotolerance | Food and feed microbiota | Lactobacilli, Actinobacteria | [14] |
| Facultative anaerobes | Ubiquitous distribution | Enterobacteriaceae, E. coli | [35,36] |
| Spore formation | Gut microbiota | Clostridiales | [32,35,37] |
| None | Gut microbiota | Bacteroidetes, Selenomonadales | [35] |
| Form of Air Resistance | Major Taxa | Vertical Transmission | Environ. Origin | Host Adaptation | Probiotic Dosage | Permanent Colonization |
|---|---|---|---|---|---|---|
| None | Bacteroidetes, Selenomonadales | Yes | No | Yes | Single | Yes |
| Aerotolerance | Lactobacilli, Actinobacteria | No | Yes | No | Repeated | No |
| Fac. anaerobes | Enterobacteriaceae | No | Yes | No | Repeated | No |
| Spores | Clostridiales | No | Yes | No | Repeated | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubasova, T.; Seidlerova, Z.; Rychlik, I. Ecological Adaptations of Gut Microbiota Members and Their Consequences for Use as a New Generation of Probiotics. Int. J. Mol. Sci. 2021, 22, 5471. https://doi.org/10.3390/ijms22115471
Kubasova T, Seidlerova Z, Rychlik I. Ecological Adaptations of Gut Microbiota Members and Their Consequences for Use as a New Generation of Probiotics. International Journal of Molecular Sciences. 2021; 22(11):5471. https://doi.org/10.3390/ijms22115471
Chicago/Turabian StyleKubasova, Tereza, Zuzana Seidlerova, and Ivan Rychlik. 2021. "Ecological Adaptations of Gut Microbiota Members and Their Consequences for Use as a New Generation of Probiotics" International Journal of Molecular Sciences 22, no. 11: 5471. https://doi.org/10.3390/ijms22115471