
The Molecular Basis of COVID-19 Pathogenesis, Conventional and Nanomedicine Therapy




Abstract
:1. Introduction
2. Pathogenesis and Therapeutic Targets
2.1. Coronavirus Structure
2.2. Coronavirus Life Cycle
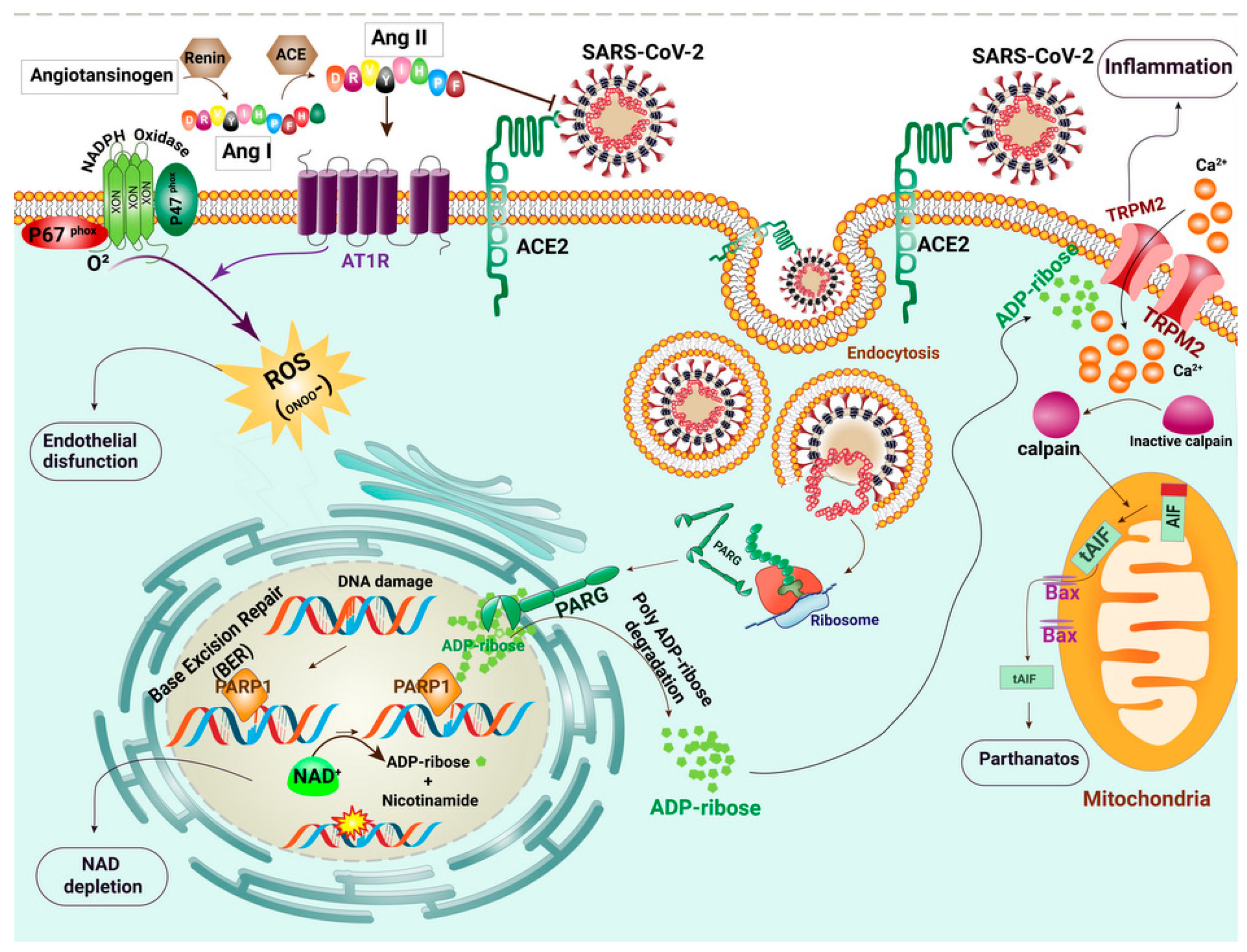
2.3. The Renin–Angiotensin System (RAS) Signaling Pathway
2.4. Oxidative Stress and Cell Death
2.5. Cytokine Storm
2.6. Endothelial Dysfunction
3. Potential Therapeutics for Management of COVID-19
3.1. Vitamin D
3.2. Thalidomide
3.3. Trehalose
3.4. N-acetylcysteine
3.5. Tannins
3.6. Flufenamic Acid/Mefenamic Acid and Clotrimazole
3.7. NAD+ and Niacin
3.8. Losartan
3.9. Remdesivir
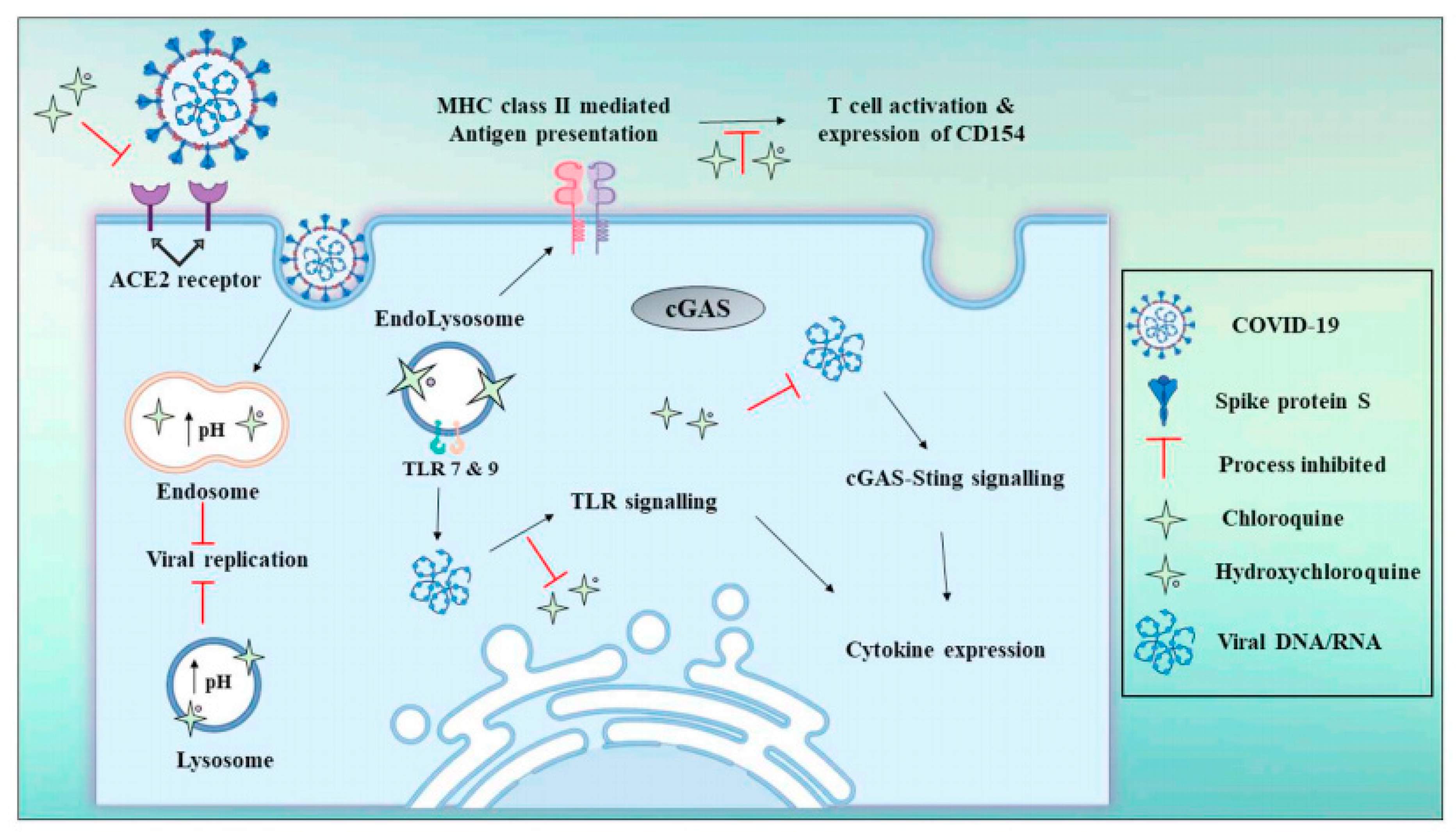
3.10. Chloroquine and Hydroxychloroquine
3.11. Monoclonal Antibodies and Recombinant Proteins
3.12. Delivery Systems: Role of Nanomedicine
3.12.1. Organic Nanoparticles
3.12.2. Inorganic Nanoparticles
4. Future Perspective
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Guo, J.; Wang, C.; Luo, F.; Yu, X.; Zhang, W.; Li, J.; Zhao, D.; Xu, D.; Gong, Q.; et al. Clinical characteristics and intrauterine vertical transmission potential of COVID-19 infection in nine pregnant women: A retrospective review of medical records. Lancet 2020, 395, 809–815. [Google Scholar] [CrossRef] [Green Version]
- Manski, C.F.; Molinari, F. Estimating the COVID-19 infection rate: Anatomy of an inference problem. J. Econ. 2021, 220, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Xing, K.; Tu, X.-Y.; Liu, M.; Liang, Z.-W.; Chen, J.-N.; Li, J.-J.; Jiang, L.-G.; Xing, F.-Q.; Jiang, Y. Efficacy and safety of COVID-19 vaccines: A systematic review. Zhongguo Dang Dai Er Ke Za Zhi 2021, 23, 221–228. [Google Scholar]
- Karpinski, T.M.; Ozarowski, M.; Seremak-Mrozikiewicz, A.; Wolski, H.; Wlodkowic, D. The 2020 race towards SARS-CoV-2 specific vaccines. Theranostics 2021, 11, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Mo, P.; Xing, Y.; Xiao, Y.; Deng, L.; Zhao, Q.; Wang, H.; Xiong, Y.; Cheng, Z.; Gao, S.; Liang, K.; et al. Clinical charac-teristics of refractory COVID-19 pneumonia in Wuhan, China. Clin. Infect. Dis. 2020, ciaa270. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.-Y.; Zhao, R.; Gao, L.-J.; Gao, X.-F.; Wang, D.-P.; Cao, J.-M. SARS-CoV-2: Structure, Biology, and Structure-Based Therapeutics Development. Front. Cell. Infect. Microbiol. 2020, 10, 587269. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 183, 1735. [Google Scholar] [CrossRef]
- Abdel-Moneim, A. COVID-19 Pandemic and Male Fertility: Clinical Manifestations and Pathogenic Mechanisms. Biochemistry (Moscow) 2021, 86, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zafer, M.M.; El-Mahallawy, H.A.; Ashour, H.M. Severe COVID-19 and Sepsis: Immune Pathogenesis and Laboratory Markers. Microorganisms 2021, 9, 159. [Google Scholar] [CrossRef]
- Soltani, S.; Tabibzadeh, A.; Zakeri, A.; Zakeri, A.M.; Latifi, T.; Shabani, M.; Pouremamali, A.; Erfani, Y.; Pakzad, I.; Malekifar, P.; et al. COVID-19 associated central nervous system manifestations, mental and neurological symptoms: A systematic review and meta-analysis. Rev. Neurosci. 2021, 32, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Gavriatopoulou, M.; Korompoki, E.; Fotiou, D.; Ntanasis-Stathopoulos, I.; Psaltopoulou, T.; Kastritis, E.; Terpos, E.; Dimopoulos, M.A. Organ-specific manifestations of COVID-19 infection. Clin. Exp. Med. 2020, 20, 493–506. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- V’Kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Nakagawa, K.; Lokugamage, K.; Makino, S. Viral and Cellular mRNA Translation in Coronavirus-Infected Cells. Adv. Appl. Microbiol. 2016, 96, 165–192. [Google Scholar] [CrossRef]
- Kim, D.; Lee, J.Y.; Yang, J.S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e910. [Google Scholar] [CrossRef]
- Raj, R. Analysis of non-structural proteins, NSPs of SARS-CoV-2 as targets for computational drug designing. Biochem. Biophys. Rep. 2021, 25, 100847. [Google Scholar]
- Pelkmans, L.; Helenius, A. Insider information: What viruses tell us about endocytosis. Curr. Opin. Cell Biol. 2003, 15, 414–422. [Google Scholar] [CrossRef]
- Wit, E.D.; Doremalen, N.V.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Belouzard, S.; Chu, V.C.; Whittaker, G.R. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc. Natl. Acad. Sci. USA 2009, 106, 5871–5876. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y. Potent binding of 2019 novel coro-navirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wevers, B.A.; Hoek, L.v.d. Renin–angiotensin system in human coronavirus pathogenesis. Futur. Virol. 2010, 5, 145–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millet, J.K.; Whittaker, G.R. Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc. Natl. Acad. Sci. USA 2014, 111, 15214–15219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Genet. 2009, 7, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Villamil-Gómez, W.E.; Sánchez, Á.; Gelis, L.; Silvera, L.A.; Barbosa, J.; Otero-Nader, O.; Bonilla-Salgado, C.D.; Rodríguez-Morales, A.J. Fatal human coronavirus 229E (HCoV-229E) and RSV–Related pneumonia in an AIDS patient from Colombia. Travel Med. Infect. Dis. 2020, 36, 101573. [Google Scholar] [CrossRef] [PubMed]
- Bayati, A.; Kumar, R.; Francis, V.; McPherson, P.S. SARS-CoV-2 uses clathrin-mediated endocytosis to gain access into cells. BioRxiv 2020. [Google Scholar] [CrossRef]
- Yang, N.; Shen, H.-M. Targeting the Endocytic Pathway and Autophagy Process as a Novel Therapeutic Strategy in COVID-19. Int. J. Biol. Sci. 2020, 16, 1724–1731. [Google Scholar] [CrossRef]
- Hartenian, E.; Nandakumar, D.; Lari, A.; Ly, M.; Tucker, J.M.; Glaunsinger, B.A. The molecular virology of Corona-viruses. J. Biol. Chem. 2020, 295, 12910–12934. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.; Hsieh, F.; Baronas, E.; Godbout, K.; Gosselin, M.; Stagliano, N.; Donovan, M.; Woolf, B.; Robison, K.; Jeyaseelan, R.; et al. A Novel Angiotensin-Converting Enzyme–Related Carboxypeptidase (ACE2) Converts Angiotensin I to Angiotensin 1-9. Circ. Res. 2000, 87, E1–E9. [Google Scholar] [CrossRef]
- Fountain, J.H.; Lappin, S.L. Physiology, Renin Angiotensin System; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhao, Z.; Wang, Y.; Zhou, Y.; Ma, Y.; Zuo, W. Single-Cell RNA Expression Profiling of ACE2, the putative receptor of Wuhan COVID-19. BioRxiv 2020. [Google Scholar] [CrossRef]
- Martins, L.D.B.U.; Jabour, L.G.P.P.; Vieira, C.C.; Nery, L.C.C.; Dias, R.F.; Silva, A.C.S.E. Renin-angiotensin system (RAS) and immune system profile in specific subgroups with COVID-19. Curr. Med. Chem. 2020, 27, 1–28. [Google Scholar] [CrossRef]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef] [Green Version]
- Gurwitz, D. Angiotensin receptor blockers as tentative SARS-CoV-2 therapeutics. Drug Dev. Res. 2020, 81, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Wu, J.Q.; Kosten, T.R.; Zhang, X.Y. Free radicals, antioxidant defense systems, and schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 46, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Kryston, T.B.; Georgiev, A.B.; Pissis, P.; Georgakilas, A.G. Role of oxidative stress and DNA damage in human car-cinogenesis. Mutat. Res. 2011, 711, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Mathews, M.T.; Berk, B. PARP-1 Inhibition Prevents Oxidative and Nitrosative Stress–Induced Endothelial Cell Death via Transactivation of the VEGF Receptor 2. Arter. Thromb. Vasc. Biol. 2008, 28, 711–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewald, M.E.; Chen, Y.; Kuny, C.; Maejima, T.; Lease, R.; Ferraris, D.; Aikawa, M.; Sullivan, C.S.; Perlman, S.; Fehr, A.R. The coronavirus macrodomain is required to prevent PARP-mediated inhibition of virus replication and enhancement of IFN expression. PLoS Pathog. 2019, 15, e1007756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrabi, S.A.; Umanah, G.K.E.; Chang, C.; Stevens, D.A.; Karuppagounder, S.S.; Gagné, J.-P.; Poirier, G.G.; Dawson, V.L.; Dawson, T.M. Poly(ADP-ribose) polymerase-dependent energy depletion occurs through inhibition of glycolysis. Proc. Natl. Acad. Sci. USA 2014, 111, 10209–10214. [Google Scholar] [CrossRef] [Green Version]
- Lange, I.; Yamamoto, S.; Partida-Sanchez, S.; Mori, Y.; Fleig, A.; Penner, R. TRPM2 functions as a lysosomal Ca2+-release channel in beta cells. Sci. Signal. 2009, 2, ra23. [Google Scholar] [CrossRef] [Green Version]
- Perraud, A.-L.; Fleig, A.; Dunn, C.A.; Bagley, L.A.; Launay, P.; Schmitz, C.; Stokes, A.J.; Zhu, Q.; Bessman, M.J.; Penner, R.; et al. ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nat. Cell Biol. 2001, 411, 595–599. [Google Scholar] [CrossRef]
- Ishii, M.; Shimizu, S.; Hagiwara, T.; Wajima, T.; Miyazaki, A.; Mori, Y.; Kiuchi, Y. Extracellular-Added ADP-Ribose Increases Intracellular Free Ca2+ Concentration through Ca2+ Release from Stores, but Not Through TRPM2-Mediated Ca2+ Entry, in Rat β-Cell Line RIN-5F. J. Pharmacol. Sci. 2006, 101, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Sumoza-Toledo, A.; Penner, R. TRPM2: A multifunctional ion channel for calcium signalling. J. Physiol. 2011, 589, 1515–1525. [Google Scholar] [CrossRef]
- Gadotti, A.C.; Lipinski, A.L.; Vasconcellos, F.T.; Marqueze, L.F.; Cunha, E.B.; Campos, A.C.; Oliveira, C.F.; Amaral, A.N.; Baena, C.P.; Telles, J.P.; et al. Susceptibility of the patients infected with Sars-Cov2 to oxidative stress and possible interplay with severity of the disease. Free Radic. Biol. Med. 2021, 165, 184–190. [Google Scholar] [CrossRef]
- Boshtam, M.; Asgary, S.; Kouhpayeh, S.; Shariati, L.; Khanahmad, H. Aptamers Against Pro- and Anti-Inflammatory Cytokines: A Review. Inflammation 2017, 40, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdeh, M.S.; Kouhpayeh, S.; Sabzehei, F.; Khanahmad, H.; Salehi, M.; Mohammadi, Z.; Shariati, L.; Hejazi, Z.; Rabiei, P.; Manian, M. Zinc Finger Nuclease: A New Approach to Overcome Beta-Lactam Antibiotic Resistance. Jundishapur J. Microbiol. 2016, 9, 29384. [Google Scholar] [CrossRef] [Green Version]
- Shariati, L.; Modarressi, M.H.; Tabatabaiefar, M.A.; Kouhpayeh, S.; Hejazi, Z.; Shahbazi, M.; Sabzehei, F.; Salehi, M.; Khanahmad, H. Engineered zinc-finger nuclease to generate site-directed modification in the KLF1 gene for fetal hemoglobin induction. J. Cell. Biochem. 2019, 120, 8438–8446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zhang, X.R.; Ju, Z.Y.; He, W.F. Advances in the research of cytokine storm mechanism induced by Corona Virus Disease 2019 and the corresponding immunotherapies. Zhonghua Shao Shang Za Zhi 2020, 36, E005. [Google Scholar]
- Dastoli, S.; Bennardo, L.; Patruno, C.; Nistico, S.P. Are erythema multiforme and urticaria related to a better outcome of COVID-19? Dermatol. Ther. 2020, 33, e13681. [Google Scholar] [CrossRef]
- Caricchio, R.; Gallucci, M.; Dass, C.; Zhang, X.; Gallucci, S.; Fleece, D.; Bromberg, M.; Criner, G.J. Preliminary predictive criteria for COVID-19 cytokine storm. Ann. Rheum. Dis. 2021, 80, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Choudhury, A.; Das, N.C.; Patra, R.; Mukherjee, S. In silico analyses on the comparative sensing of SARS-CoV-2 mRNA by the intracellular TLRs of humans. J. Med. Virol. 2021, 93, 2476–2486. [Google Scholar] [CrossRef]
- Shimabukuro-Vornhagen, A.; Gödel, P.; Subklewe, M.; Stemmler, H.J.; Schlößer, H.A.; Schlaak, M.; Kochanek, M.; Böll, B.; Von Bergwelt-Baildon, M.S. Cytokine release syndrome. J. Immunother. Cancer 2018, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J.; Collaboration, H.A.S. COVID-19: Con-sider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033. [Google Scholar] [CrossRef]
- Moore, J.B.; June, C.H. Cytokine release syndrome in severe COVID-19. Science 2020, 368, 473–474. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Koh, D.W. Roles of Poly(ADP-Ribose) Glycohydrolase in DNA Damage and Apoptosis. Int. Rev. Cell Mol. Biol. 2013, 304, 227–281. [Google Scholar] [CrossRef]
- Guo, F.; Sun, Y.B.; Su, L.; Li, S.; Liu, Z.F.; Li, J.; Hu, X.T. Losartan attenuates paraquat-induced pulmonary fibrosis in rats. Hum. Exp. Toxicol. 2014, 34, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.A.; Canna, S.W.; Schulert, G.S.; Volpi, S.; Lee, P.Y.; Kernan, K.F.; Caricchio, R.; Mahmud, S.; Hazen, M.M.; Halyabar, O.; et al. On the Alert for Cytokine Storm: Immunopathology in COVID-19. Arthritis Rheumatol. 2020, 72, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.K. Linking endothelial dysfunction with endothelial cell activation. J. Clin. Investig. 2013, 123, 540–541. [Google Scholar] [CrossRef] [PubMed]
- Endemann, D.H.; Schiffrin, E.L. Endothelial dysfunction. J. Am. Soc. Nephrol. 2004, 15, 1983–1992. [Google Scholar] [CrossRef]
- McMahon, S.; Grondin, F.; McDonald, P.P.; Richard, D.E.; Dubois, C.M. Hypoxia-enhanced expression of the proprotein convertase furin is mediated by hypoxia-inducible factor-1: Impact on the bioactivation of proproteins. J. Biol. Chem. 2005, 280, 6561–6569. [Google Scholar] [CrossRef] [Green Version]
- Sega, F.V.D.; Fortini, F.; Spadaro, S.; Ronzoni, L.; Zucchetti, O.; Manfrini, M.; Mikus, E.; Fogagnolo, A.; Torsani, F.; Pavasini, R.; et al. Time course of endothelial dysfunction markers and mortality in COVID-19 patients: A pilot study. Clin. Transl. Med. 2021, 11, e283. [Google Scholar] [CrossRef]
- Chen, X.; Touyz, R.M.; Park, J.B.; Schiffrin, E.L. Antioxidant Effects of Vitamins C and E Are Associated with Altered Activation of Vascular NADPH Oxidase and Superoxide Dismutase in Stroke-Prone SHR. Hypertension 2001, 38, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Annuk, M.; Zilmer, M.; Lind, L.; Linde, T.; Fellström, B. Oxidative Stress and Endothelial Function in Chronic Renal Failure. J. Am. Soc. Nephrol. 2001, 12, 2747–2752. [Google Scholar] [CrossRef] [PubMed]
- Heitzer, T.; Schlinzig, T.; Krohn, K.; Meinertz, T.; Münzel, T. Endothelial Dysfunction, Oxidative Stress, and Risk of Cardiovascular Events in Patients With Coronary Artery Disease. Circulation 2001, 104, 2673–2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasdan, G.; Benchetrit, S.; Rashid, G.; Green, J.; Bernheim, J.; Rathaus, M. Endothelial dysfunction and hypertension in 5/6 nephrectomized rats are mediated by vascular superoxide. Kidney Int. 2002, 61, 586–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, J.T.; Krishnan, V.; Chang, C.-Y.; Engle, S.M.; Casalini, G.; Rodgers, G.H.; Bivi, N.; Nickoloff, B.J.; Konrad, R.J.; de Bono, S.; et al. Characterization of the cytokine storm reflects hyperinflammatory endothelial dysfunction in COVID-19. J. Allergy Clin. Immunol. 2021, 147, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Furchgott, R.F. The 1996 Albert Lasker Medical Research Awards. The discovery of endothelium-derived relaxing factor and its importance in the identification of nitric oxide. JAMA 1996, 276, 1186–1188. [Google Scholar] [CrossRef]
- Sena, C.M.; Pereira, A.M.; Seiça, R. Endothelial dysfunction—A major mediator of diabetic vascular disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2013, 1832, 2216–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Caterina, R.; Libby, P.; Peng, H.B.; Thannickal, V.J.; Rajavashisth, T.B.; Gimbrone, M.A.; Shin, W.S.; Liao, J.K. Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J. Clin. Investig. 1995, 96, 60–68. [Google Scholar] [CrossRef]
- Zeiher, A.M.; Fisslthaler, B.; Schray-Utz, B.; Busse, R. Nitric oxide modulates the expression of monocyte chemoat-tractant protein 1 in cultured human endothelial cells. Circ. Res. 1995, 76, 980–986. [Google Scholar] [CrossRef]
- Eberhardt, W.; Beeg, T.; Beck, K.-F.; Walpen, S.; Gauer, S.; Böhles, H.; Pfeilschifter, J. Nitric oxide modulates expression of matrix metalloproteinase-9 in rat mesangial cells. Kidney Int. 2000, 57, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Uemura, S.; Matsushita, H.; Li, W.; Glassford, A.J.; Asagami, T.; Lee, K.H.; Harrison, D.G.; Tsao, P.S. Diabetes mellitus enhances vascular matrix metalloproteinase activity: Role of oxidative stress. Circ. Res. 2001, 88, 1291–1298. [Google Scholar] [CrossRef] [Green Version]
- Dhananjayan, R.; Koundinya, K.S.S.; Malati, T.; Kutala, V.K. Endothelial Dysfunction in Type 2 Diabetes Mellitus. Indian J. Clin. Biochem. 2016, 31, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Radomski, M.W.; Palmer, R.M.; Moncada, S. The role of nitric oxide and cGMP in platelet adhesion to vascular en-dothelium. Biochem. Biophys. Res. Commun. 1987, 148, 1482–1489. [Google Scholar] [CrossRef]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [Green Version]
- Sabioni, L.; De Lorenzo, A.; Lamas, C.; Muccillo, F.; Castro-Faria-Neto, H.C.; Estato, V.; Tibirica, E. Systemic micro-vascular endothelial dysfunction and disease severity in COVID-19 patients: Evaluation by laser Doppler perfusion moni-toring and cytokine/chemokine analysis. Microvasc. Res. 2021, 134, 104119. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Anderson, T.J. Fundamentals of Endothelial Function for the Clinical Cardiologist. Circulation 2002, 105, 546–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.; Jiang, J.; Su, L.; Shu, T.; Liu, H.; Lai, S.; Ghiladi, R.A.; Wang, J. The role of NO in COVID-19 and potential therapeutic strategies. Free Radic. Biol. Med. 2021, 163, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Dattola, A.; Silvestri, M.; Bennardo, L.; Passante, M.; Scali, E.; Patruno, C.; Nisticò, S.P. Role of Vitamins in Skin Health: A Systematic Review. Curr. Nutr. Rep. 2020, 9, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Gomez, J.M.; Castillo, M.E.; Bouillon, R. Vitamin D Receptor stimulation to reduce Acute Respiratory Distress Syndrome (ARDS) in patients with Coronavirus SARS-CoV-2 infections: Revised Ms SBMB 2020_166. J. Steroid Biochem. Mol. Biol. 2020, 202, 105719. [Google Scholar] [CrossRef]
- Rhodes, J.M.; Subramanian, S.; Laird, E.; Griffin, G.; Kenny, R.A. Perspective: Vitamin D deficiency and COVID-19 severity—Plausibly linked by latitude, ethnicity, impacts on cytokines, ACE2 and thrombosis. J. Intern. Med. 2021, 289, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, J.; Chen, J.; Luo, Q.; Zhang, Q.; Zhang, H. Vitamin D alleviates lipopolysaccharide-induced acute lung injury via regulation of the renin-angiotensin system. Mol. Med. Rep. 2017, 16, 7432–7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilz, S.; Tomaschitz, A.; Ritz, E.; Pieber, T.R. Vitamin D status and arterial hypertension: A systematic review. Nat. Rev. Cardiol. 2009, 6, 621–630. [Google Scholar] [CrossRef]
- Andersen, L.B.; Przybyl, L.; Haase, N.; von Versen-Höynck, F.; Qadri, F.; Jørgensen, J.S.; Sørensen, G.L.; Fruekilde, P.; Poglitsch, M.; Szijarto, I.; et al. Vitamin D Depletion Aggravates Hypertension and Target-Organ Damage. J. Am. Heart Assoc. 2015, 4, 001417. [Google Scholar] [CrossRef] [Green Version]
- Mabley, J.G.; Wallace, R.; Pacher, P.; Murphy, K.; Szabo, C. Inhibition of poly(adenosine diphosphate-ribose) poly-merase by the active form of vitamin D. Int. J. Mol. Med. 2007, 19, 947–952. [Google Scholar]
- Dastan, F.; Tabarsi, P.; Marjani, M.; Moniri, A.; Hashemian, S.M.R.; Tavakoli-Ardakani, M.; Saffaei, A. Thalidomide against Coronavirus Disease 2019 (COVID-19): A Medicine with a Thousand Faces. Iran. J. Pharm. Res. 2020, 19, 1–2. [Google Scholar] [PubMed]
- Wang, G.; Ma, H.; Wang, J.; Khan, M.F. Contribution of poly(ADP-ribose)polymerase-1 activation and apoptosis in trichloroethene-mediated autoimmunity. Toxicol. Appl. Pharmacol. 2019, 362, 28–34. [Google Scholar] [CrossRef]
- Martinon, D.; Borges, V.F.; Gomez, A.C.; Shimada, K. Potential Fast COVID-19 Containment With Trehalose. Front. Immunol. 2020, 11, 1623. [Google Scholar] [CrossRef]
- Lee, H.J.; Yoon, Y.S.; Lee, S.J. Mechanism of neuroprotection by trehalose: Controversy surrounding autophagy in-duction. Cell Death Dis. 2018, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, M.; Pal, R.; Nelvagal, H.R.; Lotfi, P.; Stinnett, G.R.; Seymour, M.L.; Chaudhury, A.; Bajaj, L.; Bondar, V.V.; Bremner, L. mTORC1-independent TFEB activation via Akt inhibition promotes cellular clearance in neurodegenerative storage diseases. Nat. Commun. 2017, 8, 1–19. [Google Scholar]
- De Flora, S.; Balansky, R.; La Maestra, S. Rationale for the use of N-acetylcysteine in both prevention and adjuvant therapy of COVID-19. FASEB J. 2020, 34, 13185–13193. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Yang, X.; Huang, A.; Chen, Z. The potential mechanism of N-acetylcysteine in treating COVID-19. Curr. Pharm. Biotechnol. 2020, 21, 1–9. [Google Scholar] [CrossRef]
- Amarowicz, R.; Pegg, R.B. Tree Nuts and Peanuts as a Source of Natural Antioxidants in our Daily Diet. Curr. Pharm. Des. 2020, 26, 1898–1916. [Google Scholar] [CrossRef] [PubMed]
- Yeo, J.; Lee, J.; Yoon, S.; Kim, W.J. Tannic acid-based nanogel as an efficient anti-inflammatory agent. Biomater. Sci. 2020, 8, 1148–1159. [Google Scholar] [CrossRef]
- Orlowski, P.; Żmigrodzka, M.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Czupryn, M.; Antos-Bielska, M.; Szemraj, J.; Celichowski, G.; Grobelny, J.; Krzyzowska, M. Tannic acid-modified silver nanoparticles for wound healing: The importance of size. Int. J. Nanomed. 2018, ume 13, 991–1007. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, I.; Zhu, W.; Mohammed, H.H.H.; Dutta, K.; Li, C. Tannins inhibit SARS-CoV-2 through binding with catalytic dyad residues of 3CLpro: An in silico approach with 19 structural different hydrolysable tannins. J. Food Biochem. 2020, 44, e13432. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, M.; Zhan, K.; Yang, W.; Zhang, L.; Wang, K.; Yu, P.; Zhang, L. Selective inhibition of TRPM2 channel by two novel synthesized ADPR analogues. Chem. Biol. Drug Des. 2018, 91, 552–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, C.; Mankoo, H.; Wei, L.; An, X.; Li, C.; Li, D.; Jiang, L. TRPM2 channel: A novel target for alleviating ischaemia-reperfusion, chronic cerebral hypo-perfusion and neonatal hypoxic-ischaemic brain damage. J. Cell. Mol. Med. 2020, 24, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Li, K.; Yan, Q.; Koizumi, S.; Shi, L.; Takahashi, S.; Zhu, Y.; Matsue, H.; Takeda, M.; Kitamura, M.; et al. Non-steroidal anti-inflammatory drug flufenamic acid is a potent activator of AMP-activated protein kinase. J. Pharmacol. Exp. Ther. 2011, 339, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, S.; Haneda, M.; Kanasaki, K.; Sugimoto, T.; Araki, S.; Isshiki, K.; Isono, M.; Uzu, T.; Guarente, L.; Kashiwagi, A.; et al. SIRT1 inhibits transforming growth factor beta-induced apoptosis in glomerular mesangial cells via Smad7 deacetyla-tion. J. Biol. Chem. 2007, 282, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, H.; Ishii, M.; Goto, A.; Sugimoto, T. Role of serotonin type 2 receptors in regulation of aldosterone pro-duction. Am. J. Physiol. 1985, 249, E234–E238. [Google Scholar]
- Miller, R.; Wentzel, A.; Richards, G. COVID-19: NAD+ deficiency may predispose the aged, obese and type2 diabetics to mortality through its effect on SIRT1 activity. Med. Hypotheses 2020, 144, 110044. [Google Scholar] [CrossRef]
- Gharote, M.A. Role of poly (ADP) ribose polymerase-1 inhibition by nicotinamide as a possible additive treatment to modulate host immune response and prevention of cytokine storm in COVID-19. Indian J. Med Sci. 2020, 72, 25–28. [Google Scholar] [CrossRef]
- Cotton, S.A.; Herrick, A.L.; Jayson, M.I.; Freemont, A.J. TGF beta--a role in systemic sclerosis? J. Pathol. 1998, 184, 4–6. [Google Scholar] [CrossRef]
- McKay, S.; de Jongste, J.C.; Saxena, P.R.; Sharma, H.S. Angiotensin II induces hypertrophy of human airway smooth muscle cells: Expression of transcription factors and transforming growth factor-beta1. Am. J. Respir. Cell Mol. Biol. 1998, 18, 823–833. [Google Scholar] [CrossRef]
- Sanderson, N.; Factor, V.; Nagy, P.; Kopp, J.; Kondaiah, P.; Wakefield, L.; Roberts, A.B.; Sporn, M.B.; Thorgeirsson, S.S. Hepatic expression of mature transforming growth factor beta 1 in transgenic mice results in multiple tissue lesions. Proc. Natl. Acad. Sci. USA 1995, 92, 2572–2576. [Google Scholar] [CrossRef] [Green Version]
- Stula, M.; Pinto, Y.M.; Gschwend, S.; Teisman, A.C.; Van Gilst, W.H.; Böhm, M.; Dietz, R.; Paul, M. Interaction of the Renin-Angiotensin System and the Endothelin System in Cardiac Hypertrophy. J. Cardiovasc. Pharmacol. 1998, 31, S403–S405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ünübol, M.; Yavaşoğlu, I.; Acar, B.; Kadıköylü, G.; Bolaman, Z. The effect of losartan on platelet aggregation and hematological parameters in patients with newly diagnosed hypertension. Meandros Med. J. 2015, 16, 91–96. [Google Scholar] [CrossRef]
- Cardoso, P.R.G.; Matias, K.A.; Dantas, A.T.; Marques, C.D.L.; Pereira, M.C.; Duarte, A.; Rego, M.; Pitta, I.D.R.; Pitta, M. Losartan, but not Enalapril and Valsartan, Inhibits the Expression of IFN-gamma, IL-6, IL-17F and IL-22 in PBMCs from Rheumatoid Arthritis Patients. Open Rheumatol. J. 2018, 12, 160–170. [Google Scholar] [CrossRef]
- Shen, L.; Mo, H.; Cai, L.; Kong, T.; Zheng, W.; Ye, J.; Qi, J.; Xiao, Z. Losartan prevents sepsis-induced acute lung injury and decreases activation of nuclear factor kappaB and mitogen-activated protein kinases. Shock 2009, 31, 500–506. [Google Scholar] [CrossRef]
- Lanza, K.; Perez, L.G.; Costa, L.B.; Cordeiro, T.M.; Palmeira, V.A.; Ribeiro, V.T.; Simoes, E.S.A.C. Covid-19: The ren-in-angiotensin system imbalance hypothesis. Clin. Sci. (Lond.) 2020, 134, 1259–1264. [Google Scholar] [CrossRef]
- Sun, M.; Yang, J.; Sun, Y.; Su, G. Inhibitors of RAS Might Be a Good Choice for the Therapy of COVID-19 Pneumonia. Zhonghua jie he he hu xi za zhi = Chin. J. Tuberc. Respir. Dis. 2020, 43, E014. [Google Scholar]
- Bengtson, C.D.; Montgomery, R.N.; Nazir, U.; Satterwhite, L.; Kim, M.D.; Bahr, N.C.; Castro, M.; Baumlin, N.; Salathe, M. An Open Label Trial to Assess Safety of Losartan for Treating Worsening Respiratory Illness in COVID-19. Front. Med. 2021, 8, 152. [Google Scholar] [CrossRef]
- Chen, G.-L.; Zeng, B.; Eastmond, S.; E Elsenussi, S.; Boa, A.; Xu, S.-Z. Pharmacological comparison of novel synthetic fenamate analogues with econazole and 2-APB on the inhibition of TRPM2 channels. Br. J. Pharmacol. 2012, 167, 1232–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, K.; Benham, C.D.; McNulty, S.; Randall, A.D. Flufenamic acid is a pH-dependent antagonist of TRPM2 channels. Neuropharmacology 2004, 47, 450–460. [Google Scholar] [CrossRef]
- Kraft, R.; Grimm, C.; Frenzel, H.; Harteneck, C. Inhibition of TRPM2 cation channels by N -(p -amylcinnamoyl)anthranilic acid. Br. J. Pharmacol. 2006, 148, 264–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, K.; McNulty, S.; Randall, A.D. Inhibition of TRPM2 channels by the antifungal agents clotrimazole and econazole. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 370, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, H.-J.; Germann, T.; Gillen, C.; Hatt, H.; Jostock, R. Characterization of the mouse cold-menthol receptor TRPM8 and vanilloid receptor type-1 VR1 using a fluorometric imaging plate reader (FLIPR) assay. Br. J. Pharmacol. 2004, 141, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Togashi, K.; Inada, H.; Tominaga, M. Inhibition of the transient receptor potential cation channel TRPM2 by 2-aminoethoxydiphenyl borate (2-APB). Br. J. Pharmacol. 2008, 153, 1324–1330. [Google Scholar] [CrossRef]
- Jiang, H.; Zeng, B.; Chen, G.-L.; Bot, D.; Eastmond, S.; Elsenussi, S.E.; Atkin, S.L.; Boa, A.; Xu, S.-Z. Effect of non-steroidal anti-inflammatory drugs and new fenamate analogues on TRPC4 and TRPC5 channels. Biochem. Pharmacol. 2012, 83, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Kolisek, M.; Beck, A.; Fleig, A.; Penner, R. Cyclic ADP-Ribose and Hydrogen Peroxide Synergize with ADP-Ribose in the Activation of TRPM2 Channels. Mol. Cell 2005, 18, 61–69. [Google Scholar] [CrossRef]
- Moreau, C.; Kirchberger, T.; Swarbrick, J.M.; Bartlett, S.J.; Fliegert, R.; Yorgan, T.; Bauche, A.; Harneit, A.; Guse, A.H.; Potter, B.V.L. Structure–Activity Relationship of Adenosine 5′-diphosphoribose at the Transient Receptor Potential Melastatin 2 (TRPM2) Channel: Rational Design of Antagonists. J. Med. Chem. 2013, 56, 10079–10102. [Google Scholar] [CrossRef]
- Togashi, K.; Hara, Y.; Tominaga, T.; Higashi, T.; Konishi, Y.; Mori, Y.; Tominaga, M. TRPM2 activation by cyclic ADP-ribose at body temperature is involved in insulin secretion. EMBO J. 2006, 25, 1804–1815. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-C.; Deng, Q.-X.; Dai, S.-X. Remdesivir for severe acute respiratory syndrome coronavirus 2 causing COVID-19: An evaluation of the evidence. Travel Med. Infect. Dis. 2020, 35, 101647. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.-S.; Lee, P.-I.; Hsueh, P.-R. Treatment options for COVID-19: The reality and challenges. J. Microbiol. Immunol. Infect. 2020, 53, 436–443. [Google Scholar] [CrossRef]
- Harrison, C. Coronavirus puts drug repurposing on the fast track. Nat. Biotechnol. 2020, 38, 379–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the repli-cation of SARS-CoV-2 in vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef]
- Favalli, E.G.; Ingegnoli, F.; De Lucia, O.; Cincinelli, G.; Cimaz, R.; Caporali, R. COVID-19 infection and rheumatoid arthritis: Faraway, so close! Autoimmun. Rev. 2020, 19, 102523. [Google Scholar] [CrossRef]
- Devaux, C.A.; Rolain, J.-M.; Colson, P.; Raoult, D. New insights on the antiviral effects of chloroquine against coro-navirus: What to expect for COVID-19? Int. J. Antimicrobe. Agents 2020, 55, 105938. [Google Scholar] [CrossRef]
- Ibáñez, S.; Martínez, O.; Valenzuela, F.; Silva, F.; Valenzuela, O. Hydroxychloroquine and chloroquine in COVID-19: Should they be used as standard therapy? Clin. Rheumatol. 2020, 39, 2461–2465. [Google Scholar] [CrossRef]
- Colson, P.; Rolain, J.-M.; Lagier, J.-C.; Brouqui, P.; Raoult, D. Chloroquine and hydroxychloroquine as available weapons to fight COVID-19. Int. J. Antimicrob. Agents 2020, 55, 105932. [Google Scholar] [CrossRef]
- Pandey, A.; Nikam, A.N.; Shreya, A.B.; Mutalik, S.P.; Gopalan, D.; Kulkarni, S.; Padya, B.S.; Fernandes, G.; Mutalik, S.; Prassl, R. Potential therapeutic targets for combating SARS-CoV-2: Drug repurposing, clinical trials and recent advancements. Life Sci. 2020, 256, 117883. [Google Scholar] [CrossRef]
- Olchanski, N.; Hansen, R.N.; Pope, E.; D’Cruz, B.; Fergie, J.; Goldstein, M.; Krilov, L.R.; McLaurin, K.K.; Nabrit-Stephens, B.; Oster, G.; et al. Palivizumab Prophylaxis for Respiratory Syncytial Virus: Examining the Evidence Around Value. Open Forum Infect. Dis. 2018, 5, ofy031. [Google Scholar] [CrossRef] [Green Version]
- Haim, H.; Si, Z.; Madani, N.; Wang, L.; Courter, J.R.; Princiotto, A.; Kassa, A.; Degrace, M.; McGee-Estrada, K.; Mefford, M.; et al. Soluble CD4 and CD4-Mimetic Compounds Inhibit HIV-1 Infection by Induction of a Short-Lived Activated State. PLoS Pathog. 2009, 5, e1000360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roma-Rodrigues, C.; Pombo, I.; Raposo, L.; Pedrosa, P.; Fernandes, A.R.; Baptista, P. Nanotheranostics Targeting the Tumor Microenvironment. Front. Bioeng. Biotechnol. 2019, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Bidram, E.; Esmaeili, Y.; Amini, A.; Sartorius, R.; Tay, F.R.; Shariati, L.; Makvandi, P. Nanobased Platforms for Diagnosis and Treatment of COVID-19: From Benchtop to Bedside. ACS Biomater. Sci. Eng. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Madanayake, N.H.; Rienzie, R.; Adassooriya, N.M. Nanoparticles in Nanotheranostics Applications. In Nanotheranostics; Springer: Berlin/Heidelberg, Germany, 2019; pp. 19–40. [Google Scholar]
- Lanfranco, R.; Jana, P.K.; Tunesi, L.; Cicuta, P.; Mognetti, B.M.; Di Michele, L.; Bruylants, G. Kinetics of Nanoparticle–Membrane Adhesion Mediated by Multivalent Interactions. Langmuir 2019, 35, 2002–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, X.Y.; Sena-Torralba, A.; Álvarez-Diduk, R.; Muthoosamy, K.; Merkoçi, A. Nanomaterials for Nanotheranostics: Tuning Their Properties According to Disease Needs. ACS Nano 2020, 14, 2585–2627. [Google Scholar] [CrossRef] [PubMed]
- Palazzolo, S.; Bayda, S.; Hadla, M.; Caligiuri, I.; Corona, G.; Toffoli, G.; Rizzolio, F. The Clinical Translation of Organic Nanomaterials for Cancer Therapy: A Focus on Polymeric Nanoparticles, Micelles, Liposomes and Exosomes. Curr. Med. Chem. 2018, 25, 4224–4268. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.W.; Song, M.G. Organic nanomaterials: Liposomes, albumin, dendrimer, polymeric nanoparticles. In Radionanomedicine; Springer: Berlin/Heidelberg, Germany, 2018; pp. 105–123. [Google Scholar] [CrossRef]
- Öncel, M.Ö.Ö.; Garipcan, B.; Inci, F. Biomedical Applications: Liposomes and Supported Lipid Bilayers for Diagnostics, Theranostics, Imaging, Vaccine Formulation, and Tissue Engineering. In Biomimetic Lipid Membranes: Fundamentals, Applications, and Commercialization; Springer: Berlin/Heidelberg, Germany, 2019; pp. 193–212. [Google Scholar]
- Serrano, G.; Mullor, J.L.; Sanchez, A.V.; Kochergina, I.; Albors, A.; Serrano, J.M.; Hueso, G.; Oroval, M.; Mir, S.; Mor, I. Liposomal Lactoferrin Effect in Preventing SARS-CoV-2 Binding in HACAT Keratinocytes. Int. J. Res. Health Sci. 2020, 8, 16–21. [Google Scholar] [CrossRef]
- Serrano, G.; Kochergina, I.; Albors, A.; Diaz, E.; Oroval, M.; Hueso, G.; Serrano, J.M. Liposomal Lactoferrin as Potential Preventative and Cure for COVID-19. Int. J. Res. Health Sci. 2020, 8, 08–15. [Google Scholar] [CrossRef]
- Han, J.; Zhao, D.; Li, D.; Wang, X.; Jin, Z.; Zhao, K. Polymer-Based Nanomaterials and Applications for Vaccines and Drugs. Polymers 2018, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Yadav, H.K.; Almokdad, A.A.; Shaluf, S.I.; Debe, M.S. Polymer-Based Nanomaterials for Drug-Delivery Carriers. In Nanocarriers for Drug Delivery; Elsevier: Amsterdam, The Netherlands, 2019; pp. 531–556. [Google Scholar]
- Jamaledin, R.; Sartorius, R.; Di Natale, C.; Vecchione, R.; De Berardinis, P.; Netti, P.A. Recombinant Filamentous Bacteriophages Encapsulated in Biodegradable Polymeric Microparticles for Stimulation of Innate and Adaptive Immune Responses. Microorganisms 2020, 8, 650. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.; Patil, A.; Raza, B.; Reurink, D.; Hengel, S.V.D.; Rutjes, S.; Husman, A.D.R.; Roesink, H.; de Vos, W. Cationically modified membranes using covalent layer-by-layer assembly for antiviral applications in drinking water. J. Membr. Sci. 2019, 570–571, 494–503. [Google Scholar] [CrossRef]
- Mehta, P.; Kadam, S.; Pawar, A.; Bothiraja, C. Dendrimers for pulmonary delivery: Current perspectives and future challenges. New J. Chem. 2019, 43, 8396–8409. [Google Scholar] [CrossRef]
- Jones, S.T.; Cagno, V.; Janeček, M.; Ortiz, D.; Gasilova, N.; Piret, J.; Gasbarri, M.; Constant, D.A.; Han, Y.; Vuković, L.; et al. Modified cyclodextrins as broad-spectrum antivirals. Sci. Adv. 2020, 6, eaax9318. [Google Scholar] [CrossRef] [Green Version]
- Braga, S.S. Cyclodextrins: Emerging Medicines of the New Millennium. Biomolecules 2019, 9, 801. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Lu, J.; Liu, L.; Dong, N.; Fang, L.; Xiao, S.; Han, H. Antiviral Activity of Graphene Oxide–Silver Nanocomposites by Preventing Viral Entry and Activation of the Antiviral Innate Immune Response. ACS Appl. Bio Mater. 2018, 1, 1286–1293. [Google Scholar] [CrossRef]
- Ye, S.; Shao, K.; Li, Z.; Guo, N.; Zuo, Y.; Li, Q.; Lu, Z.; Chen, L.; He, Q.; Han, H. Antiviral Activity of Graphene Oxide: How Sharp Edged Structure and Charge Matter. ACS Appl. Mater. Interfaces 2015, 7, 21571–21579. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.X.; Li, C.M.; Li, Y.F.; Wang, J.; Huang, C.Z. Synergistic antiviral effect of curcumin functionalized graphene oxide against respiratory syncytial virus infection. Nanoscale 2017, 9, 16086–16092. [Google Scholar] [CrossRef]
- Donskyi, I.S.; Azab, W.; Cuellar-Camacho, J.L.; Guday, G.; Lippitz, A.; Unger, W.E.; Osterrieder, K.; Adeli, M.; Haag, R. Functionalized nanographene sheets with high antiviral activity through synergistic electrostatic and hydrophobic interactions. Nanoscale 2019, 11, 15804–15809. [Google Scholar] [CrossRef]
- Ziem, B.; Rahn, J.; Donskyi, I.; Silberreis, K.; Cuellar, L.; Dernedde, J.; Keil, G.; Mettenleiter, T.C.; Haag, R. Polyvalent 2D Entry Inhibitors for Pseudorabies and African Swine Fever Virus. Macromol. Biosci. 2017, 17, 1600499. [Google Scholar] [CrossRef]
- Deokar, A.R.; Nagvenkar, A.P.; Kalt, I.; Shani, L.; Yeshurun, Y.; Gedanken, A.; Sarid, R. Graphene-Based “Hot Plate” for the Capture and Destruction of the Herpes Simplex Virus Type 1. Bioconjugate Chem. 2017, 28, 1115–1122. [Google Scholar] [CrossRef]
- Dubrovskaya, V.; Tran, K.; Ozorowski, G.; Guenaga, J.; Wilson, R.; Bale, S.; Cottrell, C.; Turner, H.L.; Seabright, G.; O’Dell, S.; et al. Vaccination with Glycan-Modified HIV NFL Envelope Trimer-Liposomes Elicits Broadly Neutralizing Antibodies to Multiple Sites of Vulnerability. Immunity 2019, 51, 915–929.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, J.; Xie, F.; Xu, Y. When liposomes met antibodies: Drug delivery and beyond. Adv. Drug Deliv. Rev. 2020, 154-155, 151–162. [Google Scholar] [CrossRef]
- Palmieri, V.; Papi, M. Can graphene take part in the fight against COVID-19? Nano Today 2020, 33, 100883. [Google Scholar] [CrossRef]
- Salomon, N.; Vascotto, F.; Selmi, A.; Vormehr, M.; Quinkhardt, J.; Bukur, T.; Schrörs, B.; Löewer, M.; Diken, M.; Türeci, Ö.; et al. A liposomal RNA vaccine inducing neoantigen-specific CD4+ T cells augments the antitumor activity of local radiotherapy in mice. OncoImmunology 2020, 9, 1771925. [Google Scholar] [CrossRef] [PubMed]
- Evelyn Roopngam, P. Liposome and polymer-based nanomaterials for vaccine applications. Nanomed. J. 2019, 6, 1–10. [Google Scholar]
- Veeranarayanan, S.; Maekawa, T. External stimulus responsive inorganic nanomaterials for cancer theranostics. Adv. Drug Deliv. Rev. 2019, 138, 18–40. [Google Scholar] [CrossRef]
- Bose, A.; Wong, T.W. Nanotechnology-Enabled Drug Delivery for Cancer Therapy. Nanotechnol. Appl. Tissue Eng. 2015, 173–193. [Google Scholar] [CrossRef]
- Manivannan, S.; Ponnuchamy, K. Quantum dots as a promising agent to combat COVID-19. Appl. Organomet. Chem. 2020, 34, 174. [Google Scholar] [CrossRef] [PubMed]
- López-Lorente, Á.I.; Valcárcel, M. Analytical Nanoscience and Nanotechnology. In Comprehensive Analytical Chemistry; Elsevier: Amsterdam, The Netherlands, 2014; Volume 66, pp. 3–35. [Google Scholar]
- Halbus, A.F.; Horozov, T.S.; Paunov, V.N. Colloid particle formulations for antimicrobial applications. Adv. Colloid Interface Sci. 2017, 249, 134–148. [Google Scholar] [CrossRef]
- Huo, C.; Xiao, J.; Xiao, K.; Zou, S.; Wang, M.; Qi, P.; Liu, T.; Hu, Y. Pre-Treatment with Zirconia Nanoparticles Reduces Inflammation Induced by the Pathogenic H5N1 Influenza Virus. Int. J. Nanomed. 2020, 15, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The Role of Zinc in Antiviral Immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Hao, X.; Liang, X.; Zhang, Q.; Zhang, C.; Zhou, G.; Shen, S.; Jia, G.; Zhang, J. Inorganic Nanomaterials as Carriers for Drug Delivery. J. Biomed. Nanotechnol. 2016, 12, 1–27. [Google Scholar] [CrossRef]
- Lee, M.-Y.; Yang, J.-A.; Jung, H.S.; Beack, S.; Choi, J.E.; Hur, W.; Koo, H.; Kim, K.; Yoon, S.K.; Hahn, S.K. Hyaluronic acid–gold nanoparticle/interferon α complex for targeted treatment of hepatitis C virus infection. ACS Nano 2012, 6, 9522–9531. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Nayak, M.; Sahoo, G.C.; Pandey, K.; Sarkar, M.C.; Ansari, Y.; Das, V.; Topno, R.; Bhawna; Madhukar, M.; et al. Iron oxide nanoparticles based antiviral activity of H1N1 influenza A virus. J. Infect. Chemother. 2019, 25, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Abo-Zeid, Y.; Ismail, N.S.; McLean, G.R.; Hamdy, N.M. A molecular docking study repurposes FDA approved iron oxide nanoparticles to treat and control COVID-19 infection. Eur. J. Pharm. Sci. 2020, 153, 105465. [Google Scholar] [CrossRef]
- Salleh, A.; Naomi, R.; Utami, N.D.; Mohammad, A.W.; Mahmoudi, E.; Mustafa, N.; Fauzi, M.B. The Potential of Silver Nanoparticles for Antiviral and Antibacterial Applications: A Mechanism of Action. Nanomaterials 2020, 10, 1566. [Google Scholar] [CrossRef] [PubMed]
- Villeret, B.; Dieu, A.; Straube, M.; Solhonne, B.; Miklavc, P.; Hamadi, S.; Le Borgne, R.; Mailleux, A.; Norel, X.; Aerts, J.; et al. Silver Nanoparticles Impair Retinoic Acid-Inducible Gene I-Mediated Mitochondrial Antiviral Immunity by Blocking the Autophagic Flux in Lung Epithelial Cells. ACS Nano 2018, 12, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Shahabadi, N.; Khorshidi, A.; Zhaleh, H.; Kashanian, S. Synthesis, characterization, cytotoxicity and DNA binding studies of Fe3O4@ SiO2 nanoparticles coated by an antiviral drug lamivudine. J. Drug Deliv. Sci. Technol. 2018, 46, 55–65. [Google Scholar] [CrossRef]
- Croci, R.; Bottaro, E.; Chan, K.W.K.; Watanabe, S.; Pezzullo, M.; Mastrangelo, E.; Nastruzzi, C. Liposomal Systems as Nanocarriers for the Antiviral Agent Ivermectin. Int. J. Biomater. 2016, 2016, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.X.; Michiels, J.; Ariën, K.K.; New, R.; Vanham, G.; Roitt, I. Inhibition of HIV Virus by Neutralizing Vhh Attached to Dual Functional Liposomes Encapsulating Dapivirine. Nanoscale Res. Lett. 2016, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Vaillant, A. Nucleic acid polymers: Broad spectrum antiviral activity, antiviral mechanisms and optimization for the treatment of hepatitis B and hepatitis D infection. Antivir. Res. 2016, 133, 32–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz-Fernandez, M.A.; Vacas-Córdoba, E.; Malý, M.; De La Mata, F.J.; Gomez, R.; Pion, M. Antiviral mechanism of polyanionic carbosilane dendrimers against HIV-1. Int. J. Nanomed. 2016, 11, 1281–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fröhlich, T.; Hahn, F.; Belmudes, L.; Leidenberger, M.; Friedrich, O.; Kappes, B.; Couté, Y.; Marschall, M.; Tsogoeva, S.B. Synthesis of artemisinin-derived dimers, trimers and dendrimers: Investigation of their antimalarial and antiviral activities including putative mechanisms of action. Chem. Eur. J. 2018, 24, 8103–8113. [Google Scholar] [CrossRef] [PubMed]
- Paull, J.R.; Castellarnau, A.; Luscombe, C.A.; Fairley, J.K.; Heery, G.P. Astodrimer sodium, dendrimer antiviral, in-hibits replication of SARS-CoV-2 in vitro. Biorxiv 2020. [Google Scholar] [CrossRef]
- Chen, Y.-N.; Hsueh, Y.-H.; Hsieh, C.-T.; Tzou, D.-Y.; Chang, H.-T. Antiviral Activity of Graphene–Silver Nanocomposites against Non-Enveloped and Enveloped Viruses. Int. J. Environ. Res. Public Health 2016, 13, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacheo, A.; Hodek, J.; Witt, D.; Mangiatordi, G.F.; Ong, Q.K.; Kocabiyik, O.; DePalo, N.; Fanizza, E.; Laquintana, V.; Denora, N.; et al. Multi-sulfonated ligands on gold nanoparticles as virucidal antiviral for Dengue virus. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Peña-González, C.E.; García-Broncano, P.; Ottaviani, M.F.; Cangiotti, M.; Fattori, A.; Hierro-Oliva, M.; González-Martín, M.L.; Pérez-Serrano, J.; Gómez, R.; Muñoz-Fernández, M.Á. Dendronized anionic gold nanoparticles: Synthesis, characterization, and antiviral activity. Chem. Eur. J. 2016, 22, 2987–2999. [Google Scholar] [CrossRef]
- Levina, A.S.; Repkova, M.N.; Bessudnova, E.V.; Filippova, E.I.; Mazurkova, N.A.; Zarytova, V.F. High antiviral effect of TiO2·PL–DNA nanocomposites targeted to conservative regions of (−) RNA and (+) RNA of influenza A virus in cell culture. Beilstein J. Nanotechnol. 2016, 7, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Shahzad, K.; Mushtaq, S.; Ali, I.; Rafe, M.H.; Fazal-Ul-Karim, S.M. Antibacterial and antiviral potential of colloidal Titanium dioxide (TiO2) nanoparticles suitable for biological applications. Mater. Res. Express 2019, 6, 105409. [Google Scholar] [CrossRef]
- Ghaffari, H.; Tavakoli, A.; Moradi, A.; Tabarraei, A.; Bokharaei-Salim, F.; Zahmatkeshan, M.; Farahmand, M.; Javanmard, D.; Kiani, S.J.; Esghaei, M.; et al. Inhibition of H1N1 influenza virus infection by zinc oxide nanoparticles: Another emerging application of nanomedicine. J. Biomed. Sci. 2019, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T. Antiviral Activities of Cu2+ Ions in Viral Prevention, Replication, RNA Degradation, and for Antiviral Efficacies of Lytic Virus, ROS-Mediated Virus, Copper Chelation. World Sci. News 2018, 99, 148–168. [Google Scholar]
- Tavakoli, A.; Hashemzadeh, M.S. Inhibition of herpes simplex virus type 1 by copper oxide nanoparticles. J. Virol. Methods 2020, 275, 113688. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Shabaan, M.T.; Hassan, L.; Morsi, H.H. Antiviral activity of algae biosynthesized silver and gold nanoparticles against Herps Simplex (HSV-1) virus in vitro using cell-line culture technique. Int. J. Environ. Health Res. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Agnihothram, S.S.; Vermudez, S.A.; Mullis, L.; Townsend, T.A.; Manjanatha, M.G.; Azevedo, M.P. Silicon Dioxide Impedes Antiviral Response and Causes Genotoxic Insult During Calicivirus Replication. J. Nanosci. Nanotechnol. 2016, 16, 7720–7730. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanomaterial | Antiviral Mechanism | Targeted Viruses | References |
|---|---|---|---|
| Liposomes | Alter the structure of lipids and proteins of viruses via their composition (fatty acids, surfactants, alcohol) | DENV, SARS-CoV-2, HIV, Influenza | [186,187] |
| Polymers | Absorb negative charged virus particles via their positive charge and inactivate viruses | HBV, HDV, HSV, Influenza, CoVs | [155,188] |
| Dendrimers | Strong multivalent interactions with viruses via their highly-branched 3D structure and inactivate viruses | HIV, HCV, SARS-CoV-2 | [189,190,191] |
| Cyclodextrin (CD) | Directly inactivate virus and inhibit attachment | HIV, RSV, HSV-2, DENV-2, ZIKV | [159] |
| Graphene oxide (GO) | Inactivate virus function because of the interactions between the sharp edges of the GO layers and the virus particles | RSV, CoVs, HPV | [163,192] |
| Quantum dots (QDs) | Inhibit virus entry at the early stage via interaction with the spike protein of viruses and block genomic replication | SARS-CoV-2, HIV-1 | [174] |
| Gold NPs (AuNPs) | Prevent viral attachment and penetration | DENV, HCV, HPV, CoVs | [193,194] |
| Iron oxide NPs (IONPs) | Interaction with the spike protein receptor binding domain (S1-RBD) and block virus replication | HIV, Influenza A, SARS-CoV-2 | [181,182] |
| Titanium oxide NPs (TiO2NPs) | Generate ROS leading to impair lipid membranes and damage DNA structure of virus particles | CoVs, Influenza A | [195,196] |
| Zirconium oxide NPs (ZrO2NPs) | Stimulation of innate immunity and promote the expression of cytokines | H5N1 influenza virus, Influenza A | [177] |
| Zinc oxide NPs (ZnONPs) | Releasing of Zn2+, generate ROS and block viral replication and translation | HIV, HSV, HPV, H1N1 influenza | [197] |
| Copper oxide NPs (CuO2NPs) | Releasing of Cu2+, oxidation of viral proteins and degradation of viral genome | HSV | [198,199] |
| Silver NPs (AgNPs) | Iron release (Ag+), increase ROS and oxidation viral proteins, bind to the nucleic acid (DNA or RNA) of the virus | HSV, SARS-CoV-2 | [184,200] |
| Silicon dioxide NPs (SiO2NPs) | Reduce the amount of progeny virus | Influenza virus A | [201] |
| Therapeutic Agent | Current Status | Rout of Administration | Purpose of Study | Clinical Trial Identification |
|---|---|---|---|---|
| Remdesivir | Phase III | Intravenous | Antiviral drug to manage COVID-19 | NCT04678739 NCT04647695 NCT04582266 NCT04738045 |
| Chloroquine | Phase I/II/IV | Oral | Antimalarial drug to treat mild Asymptomatic and Symptomatic cases of COVID-19 | NCT04443270 NCT04328493 NCT04627467 |
| Lopinavir/ ritonavir | Phase IV/II | Intravenous | Antiviral drug to treat COVID-19 in terminally sick patients with cancer and immune suppression |
NCT04738045 NCT04455958 |
| Azithromycin | Phase III/IV | Oral | Antibiotic drug to treat COVID-19 patients |
NCT04365231 NCT04359316 |
| Hydroxy-chloroquine | Phase II/I | Oral | Antimalarial drug to treat ambulatory patients with mild and severe COVID-19 |
NCT04340544 NCT04351620 |
| Arbidol | Phase IV | Oral | Antiviral drug to treat COVID-19 patients | NCT04350684 |
| Isotretinoin | Phase III | Oral | Retinoid drug to evaluate the safety and efficacy in treatment of COVID-19 |
NCT04663906 NCT04353180 |
| Lenalidomide | Phase IV | Oral | Antiangiogenic drug to treat mild COVID-19 patients | NCT04361643 |
| Chlorpromazine | Phase III | Oral | Antipsychotics drug to manage COVID-19 subjects | NCT04366739 |
| Canakinumab | Phase III | Intravenous | Anti-human-IL-1β monoclonal antibody to study the efficacy in treating COVID-19 patients | NCT04362813 |
| Ruxolitinib | Phase III | Oral | Antiviral drug to assay the efficacy in COVID-19 patients with cytokine storm | NCT04362137 |
| Dexamethasone | Phase IV | Intravenous | Steroid drug to treat COVID-19 patients | NCT04325061 |
| Favipiravir | Phase III | Oral | Antiviral drug to treat COVID-19 patients | NCT04336904 |
| Sildenafil citrate | Phase III | Oral | Phosphodiesterase inhibitor drug to treat COVID-19 patients | NCT04304313 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouhpayeh, S.; Shariati, L.; Boshtam, M.; Rahimmanesh, I.; Mirian, M.; Esmaeili, Y.; Najaflu, M.; Khanahmad, N.; Zeinalian, M.; Trovato, M.; et al. The Molecular Basis of COVID-19 Pathogenesis, Conventional and Nanomedicine Therapy. Int. J. Mol. Sci. 2021, 22, 5438. https://doi.org/10.3390/ijms22115438
Kouhpayeh S, Shariati L, Boshtam M, Rahimmanesh I, Mirian M, Esmaeili Y, Najaflu M, Khanahmad N, Zeinalian M, Trovato M, et al. The Molecular Basis of COVID-19 Pathogenesis, Conventional and Nanomedicine Therapy. International Journal of Molecular Sciences. 2021; 22(11):5438. https://doi.org/10.3390/ijms22115438
Chicago/Turabian StyleKouhpayeh, Shirin, Laleh Shariati, Maryam Boshtam, Ilnaz Rahimmanesh, Mina Mirian, Yasaman Esmaeili, Malihe Najaflu, Negar Khanahmad, Mehrdad Zeinalian, Maria Trovato, and et al. 2021. "The Molecular Basis of COVID-19 Pathogenesis, Conventional and Nanomedicine Therapy" International Journal of Molecular Sciences 22, no. 11: 5438. https://doi.org/10.3390/ijms22115438