
The Potential Protective Role of RUNX1 in Nonalcoholic Fatty Liver Disease

 ,
,  , ,
, ,
Abstract
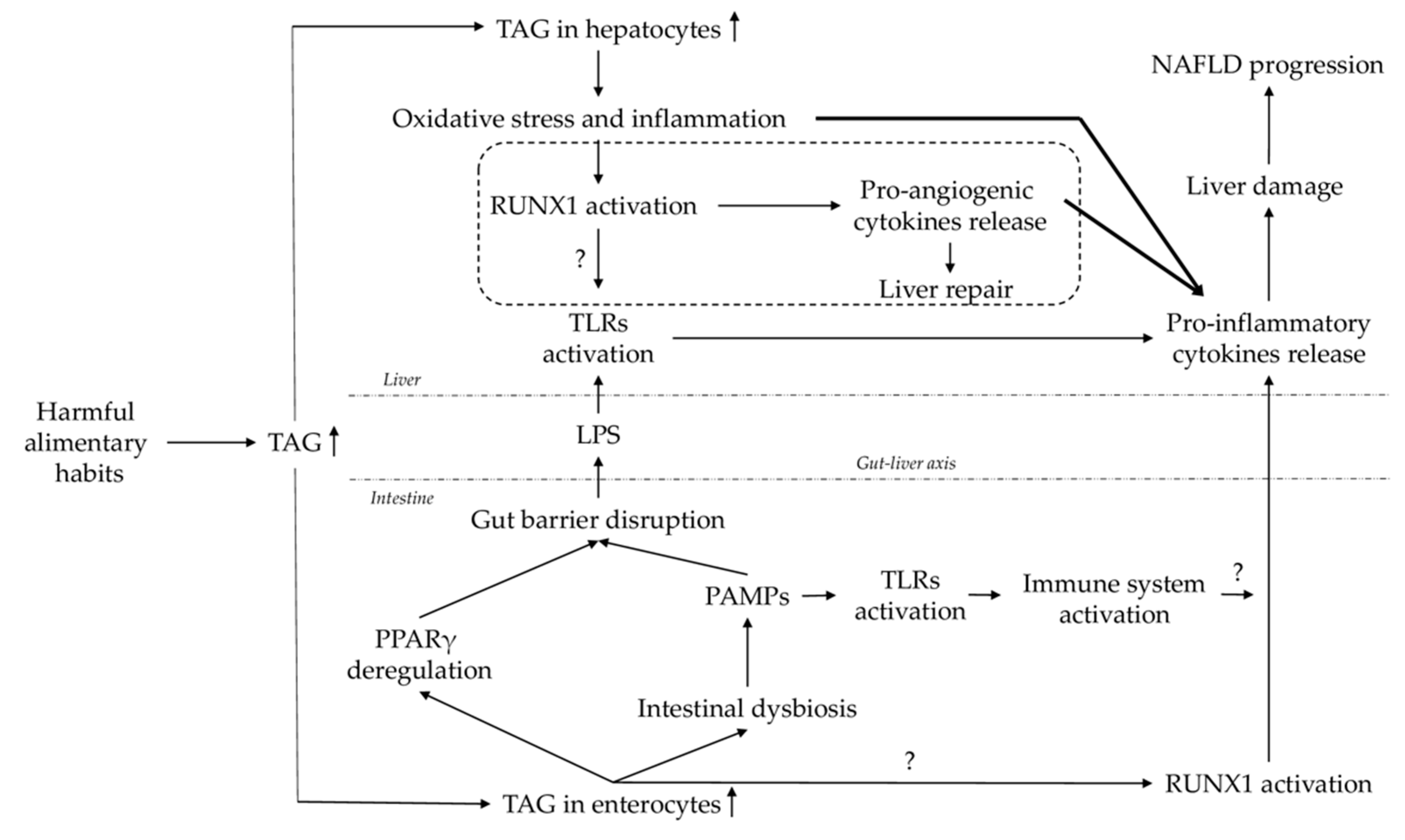
:1. Introduction
2. Results
2.1. Baseline Characteristics of Subjects
2.2. Evaluation of the Relative mRNA Abundance of RUNX1 in the Liver and Jejunum According to Hepatic Histology
2.3. Evaluation of the Immunohistochemical Analysis of RUNX1 in the Liver According to Hepatic Histology
2.4. Evaluation of Relative mRNA Abundance of Some Lipid Metabolism-Related Genes and TLRs in the Liver and Jejunum According to Hepatic Histology
2.5. Correlations of RUNX1 Relative Hepatic Expression with Different NAFLD-Related Parameters
2.6. Correlations of the RUNX1 Relative Jejunal Expression with Different NAFLD-Related Parameters
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Sample Size
4.3. Liver Pathology
4.4. Biochemical Analyses
4.5. Gene Expression in the Liver and Jejunum
4.6. Immunohistochemistry Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| ALT | Alanine aminotransferase |
| AST | Aspartate aminotransferase |
| BMI | Body mass index |
| DBP | Diastolic blood pressure |
| FAS | Fatty acid synthase |
| FXR | Farnesoid X receptor |
| GGT | Gamma-glutamyltransferase |
| HbA1c | Glycosylated hemoglobin |
| HCC | Hepatocellular carcinoma |
| HDL-C | High density lipoprotein cholesterol |
| HOMA1-IR | Homeostatic model assessment method 1 of insulin resistance |
| IL | Interleukin |
| LDH | Lactate dehydrogenase |
| LDL-C | Low density lipoprotein cholesterol |
| LXR | Liver X receptor |
| MetS | Metabolic syndrome |
| MO | Morbid obesity |
| NAFLD | Nonalcoholic fatty liver disease |
| NASH | Nonalcoholic steatohepatitis |
| NL | Normal liver |
| PPARγ | Peroxisome proliferator-activated receptor γ |
| RUNX1 | Runt-related transcription factor 1 |
| SBP | Systolic blood pressure |
| SREBP1c | Sterol regulatory element binding protein-1c |
| SS | Simple steatosis |
| TNF-α | Tumor necrosis factor α |
| TLR | Toll-like receptor |
References
- Cotter, T.G.; Rinella, M. Nonalcoholic Fatty Liver Disease 2020: The State of the Disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The Diagnosis and Management of Nonalcoholic Fatty Liver Disease: Practice Guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef]
- Fazel, Y.; Koenig, A.B.; Sayiner, M.; Goodman, Z.D.; Younossi, Z.M. Epidemiology and Natural History of Non-Alcoholic Fatty Liver Disease. Metabolism 2016, 65, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Neuman, M.G.; Cohen, L.B.; Nanau, R.M. Biomarkers in Nonalcoholic Fatty Liver Disease. Can. J. Gastroenterol. Hepatol. 2014, 28, 607–618. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and Validation of a Histological Scoring System for Nonalcoholic Fatty Liver Disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The Multiple-Hit Pathogenesis of Non-Alcoholic Fatty Liver Disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R. Evolution of Inflammation in Nonalcoholic Fatty Liver Disease: The Multiple Parallel Hits Hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Aragonès, G.; Colom-Pellicer, M.; Aguilar, C.; Guiu-Jurado, E.; Martínez, S.; Sabench, F.; Antonio Porras, J.; Riesco, D.; Del Castillo, D.; Richart, C.; et al. Circulating Microbiota-Derived Metabolites: A “liquid Biopsy? Int. J. Obes. 2020, 44, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Liaskou, E.; Wilson, D.V.; Oo, Y.H. Innate Immune Cells in Liver Inflammation. Mediat. Inflamm. 2012, 2012, 949157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.; Xu, C.; Yu, C.; Li, Y. Role of NLRP3 Inflammasome in the Progression of NAFLD to NASH. Can. J. Gastroenterol. Hepatol. 2016, 2016, 6489012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Xu, D.; Xue, B.; Liu, B.; Li, J.; Huang, J. Upregulation of RUNX1 Suppresses Proliferation and Migration through Repressing VEGFA Expression in Hepatocellular Carcinoma. Pathol. Oncol. Res. 2020, 26, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- North, T.E.; de Bruijn, M.F.T.R.; Stacy, T.; Talebian, L.; Lind, E.; Robin, C.; Binder, M.; Dzierzak, E.; Speck, N.A. Runx1 Expression Marks Long-Term Repopulating Hematopoietic Stem Cells in the Midgestation Mouse Embryo. Immunity 2002, 16, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Iwatsuki, K.; Tanaka, K.; Kaneko, T.; Kazama, R.; Okamoto, S.; Nakayama, Y.; Ito, Y.; Satake, M.; Takahashi, S.-I.; Miyajima, A.; et al. Runx1 Promotes Angiogenesis by Downregulation of Insulin-like Growth Factor-Binding Protein-3. Oncogene 2005, 24, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Semela, D.; Bruix, J.; Colle, I.; Pinzani, M.; Bosch, J. Angiogenesis in Liver Disease. J. Hepatol. 2009, 50, 604–620. [Google Scholar] [CrossRef]
- Kaur, S.; Rawal, P.; Siddiqui, H.; Rohilla, S.; Sharma, S.; Tripathi, D.M.; Baweja, S.; Hassan, M.; Vlaic, S.; Guthke, R.; et al. Increased Expression of RUNX1 in Liver Correlates with NASH Activity Score in Patients with Non-Alcoholic Steatohepatitis (NASH). Cells 2019, 8, 1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulon, S. Evaluation of Inflammatory and Angiogenic Factors in Patients with Non-Alcoholic Fatty Liver Disease. Cytokine 2012, 59, 442–449. [Google Scholar] [CrossRef]
- Coulon, S.; Legry, V.; Heindryckx, F.; Van Steenkiste, C.; Casteleyn, C.; Olievier, K.; Libbrecht, L.; Carmeliet, P.; Jonckx, B.; Stassen, J.-M.; et al. Role of Vascular Endothelial Growth Factor in the Pathophysiology of Nonalcoholic Steatohepatitis in Two Rodent Models. Hepatology 2013, 57, 1793–1805. [Google Scholar] [CrossRef]
- Kitade, M.; Yoshiji, H.; Kojima, H.; Ikenaka, Y.; Noguchi, R.; Kaji, K.; Yoshii, J.; Yanase, K.; Namisaki, T.; Yamazaki, M.; et al. Neovascularization and Oxidative Stress in the Progression of Non-Alcoholic Steatohepatitis. Mol. Med. Rep. 2008. [Google Scholar] [CrossRef] [Green Version]
- Tailleux, A.; Wouters, K.; Staels, B. Roles of PPARs in NAFLD: Potential Therapeutic Targets. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2012, 1821, 809–818. [Google Scholar] [CrossRef]
- Luo, M.-C.; Zhou, S.-Y.; Feng, D.-Y.; Xiao, J.; Li, W.-Y.; Xu, C.-D.; Wang, H.-Y.; Zhou, T. Runt-Related Transcription Factor 1 (RUNX1) Binds to P50 in Macrophages and Enhances TLR4-Triggered Inflammation and Septic Shock. J. Biol. Chem. 2016, 291, 22011–22020. [Google Scholar] [CrossRef] [Green Version]
- Kiziltas, S. Toll-like Receptors in Pathophysiology of Liver Diseases. WJH 2016, 8, 1354–1369. [Google Scholar] [CrossRef] [PubMed]
- Villard, A.; Boursier, J.; Andriantsitohaina, R. Bacterial and Eukaryotic Extracellular Vesicles and Non-Alcoholic Fatty Liver Disease: New Players in the Gut-Liver Axis? Am. J. Physiol. Gastrointest. Liver Physiol. 2021. [Google Scholar] [CrossRef]
- Rodríguez-Acebes, S.; Palacios, N.; Botella-Carretero, J.I.; Olea, N.; Crespo, L.; Peromingo, R.; Gómez-Coronado, D.; Lasunción, M.A.; Vázquez, C.; Martínez-Botas, J. Gene Expression Profiling of Subcutaneous Adipose Tissue in Morbid Obesity Using a Focused Microarray: Distinct Expression of Cell-Cycle- and Differentiation-Related Genes. BMC Med. Genom. 2010, 3, 61. [Google Scholar] [CrossRef] [Green Version]
- Auguet, T.; Berlanga, A.; Guiu-Jurado, E.; Martinez, S.; Porras, J.; Aragonès, G.; Sabench, F.; Hernandez, M.; Aguilar, C.; Sirvent, J.; et al. Altered Fatty Acid Metabolism-Related Gene Expression in Liver from Morbidly Obese Women with Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 22173–22187. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Kodama, Y.; Inokuchi, S.; Schnabl, B.; Aoyama, T.; Ohnishi, H.; Olefsky, J.M.; Brenner, D.A.; Seki, E. Toll-Like Receptor 9 Promotes Steatohepatitis by Induction of Interleukin-1β in Mice. Gastroenterology 2010, 139, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mridha, A.R.; Haczeyni, F.; Yeh, M.M.; Haigh, W.G.; Ioannou, G.N.; Barn, V.; Ajamieh, H.; Adams, L.; Hamdorf, J.M.; Teoh, N.C.; et al. TLR9 Is Up-Regulated in Human and Murine NASH: Pivotal Role in Inflammatory Recruitment and Cell Survival. Clin. Sci. 2017, 131, 2145–2159. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolic Syndrome and Altered Gut Microbiota in Mice Lacking Toll-Like Receptor 5. Science 2010, 328, 228–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Olivos, N.E. The Nuclear Receptor FXR, but Not LXR, up-Regulates Bile Acid Transporter Expression in Non-Alcoholic Fatty Liver Disease. Anal. Hepatol. 2015, 14, 7487–7493. [Google Scholar] [CrossRef]
- Kato, K.; Satoh, H.; Endo, Y.; Yamada, D.; Midorikawa, S.; Sato, W.; Mizuno, K.; Fujita, T.; Tsukamoto, K.; Watanabe, T. Thiazolidinediones Down-Regulate Plasminogen Activator Inhibitor Type 1 Expression in Human Vascular Endothelial Cells: A Possible Role for PPAR␥ in Endothelial Function. Biochem. Biophys. Res. Commun. 1999, 258, 5. [Google Scholar] [CrossRef]
- Liu, H.B.; Hu, Y.S.; Medcalf, R.L.; Simpson, R.W.; Dear, A.E. Thiazolidinediones Inhibit TNFα Induction of PAI-1 Independent of PPARγ Activation. Biochem. Biophys. Res. Commun. 2005, 334, 30–37. [Google Scholar] [CrossRef]
- Ye, P.; Hu, X.; Zhao, Y. The Increase in Plasminogen Activator Inhibitor Type-1 Expression by Stimulation of Activators for Peroxisome Proliferator-Activated Receptors in Human Endothelial Cells. Chin. Med. Sci. J. 2002, 17, 112–116. [Google Scholar]
- Józkowicz, A.; Huk, I.; Nigisch, A.; Weigel, G.; Dietrich, W.; Motterlini, R.; Dulak, J. Heme Oxygenase and Angiogenic Activity of Endothelial Cells: Stimulation by Carbon Monoxide and Inhibition by Tin Protoporphyrin-IX. Antioxid. Redox Signal. 2003, 5, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Ghoshal, U.; Ranjan, P.; Ghoshal, U.C. Expression of Toll-like Receptors, Pro-, and Anti-Inflammatory Cytokines in Relation to Gut Microbiota in Irritable Bowel Syndrome: The Evidence for Its Micro-Organic Basis. J. Neurogastroenterol. Motil. 2018, 24, 628–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamdar, K.; Nguyen, V.; DePaolo, R.W. Toll-like Receptor Signaling and Regulation of Intestinal Immunity. Virulence 2013, 4, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | NL | NAFLD | SS | NASH |
|---|---|---|---|---|
| (n = 28) | (n = 41) | (n = 24) | (n = 17) | |
| Weight (kg) | 116.50 (107.25–130.50) | 112.40 (106.00–128.00) | 113.20 (108.33–128.00) | 112.00 (104.65–125.00) |
| BMI (kg/m2) | 43.30 (40.94–46.47) | 44.46 (40.84–46.60) | 44.35 (40.82–46.83) | 44.46 (40.76–46.03) |
| Glucose (mg/dL) | 85.50 (76.25–93.00) | 93.00 (84.00–105.00) * | 93.50 (85.75–107.00) # | 91.00 (82.50–101.20) |
| Insulin (mUI/L) | 9.43 (5.59–16.21) | 9.63 (5.88–14.52) | 11.27 (7.81–14.51) | 6.57 (5.09–23.04) |
| HOMA1-IR | 2.07 (1.06–3.35) | 2.14 (1.27–3.97) | 2.70 (1.59–4.77) | 1.57 (1.05–4.12) |
| HbA1c (%) | 5.40 (5.30–5.70) | 5.60 (5.20–6.05) | 5.60 (5.25–6.03) | 5.50 (5.20–6.10) |
| Cholesterol (mg/dL) | 171.88 ± 36.20 | 179.07 ± 38.80 | 174.42 ± 35.41 | 185.28 ± 43.39 |
| HDL-C (mg/dL) | 40.84 ± 9.89 | 41.04 ± 10.95 | 42.56 ± 12.38 | 38.89 ± 8.47 |
| LDL-C (mg/dL) | 108.16 ± 27.94 | 104.48 ± 30.86 | 104.39 ± 31.21 | 104.62 ± 31.58 |
| Triglycerides (mg/dL) | 106.00 (89.00–136.00) | 132.00 (91.00–189.00) | 129.50 (85.75–175.50) | 140.00 (106.00–247.00) |
| AST (UI/L) | 20.50 (15.75–36.25) | 23.50 (17.00–41.75) | 23.00 (17.00–35.00) | 24.00 (17.00–43.00) |
| ALT (UI/L) | 22.00 (16.00–27.00) | 31.00 (21.00–37.00) | 31.00 (23.00–35.75) # | 30.00 (15.50–40.00) |
| GGT (UI/L) | 18.00 (16.00–27.00) | 22.00 (16.00–27.00) | 21.00 (16.25–30.50) | 25.00 (15.00–27.00) |
| ALP (Ul/L) | 60.42 ± 13.09 | 70.67 ± 13.01 * | 75.80 ± 11.66 # | 62.77 ± 11.16 & |
| LDH (Ul/L) | 378.65 ± 54.30 | 421.00 ± 82.47 | 428.08 ± 79.38 | 411.80 ± 89.76 |
| SBP (mmHg) | 119.00 ± 18.26 | 117.29 ± 13.86 | 120.09 ± 13.41 | 113.44 ± 13.96 |
| DBP (mmHg) | 63.00 (57.75–75.75) | 62.00 (59.00–71.25) | 62.00 (59.00–72.50) | 65.50 (56.75–70.75) |
| IL-1β (pg/mL) | 4.14 ± 2.07 | 5.10 ± 2.93 | 4.91 ± 2.15 | 5.38 ± 3.84 |
| IL-6 (pg/mL) | 4.24 ± 2.51 | 3.65 ± 1.83 | 3.40 ± 1.54 | 4.01 ± 2.20 |
| IL-8 (pg/mL) | 2.84 (2.30–3.48) ¬ | 3.61 (2.82–4.54) * | 3.47 (2.56–4.09) | 4.18 (3.12–5.29) & |
| TNF-α (pg/mL) | 10.07 (7.92–11.71) | 10.44 (7.50–12.26) | 9.73 (7.36–11.23) | 11.31 (7.80–12.57) |
| IL-10 (pg/mL) | 3.99 (1.44–7.30) | 2.92 (1.49–5.95) | 2.79 (1.78–9.96) | 2.93 (1.16–5.87) |
| Adiponectin (ng/mL) | 14,209.54 ± 6519.51 | 12,187.96 ± 6707.07 | 13,263.39 ± 7238.02 | 10,709.23 ± 5795.64 |
| Resistin (ng/mL) | 26.16 (19.72–34.94) | 26.27 (19.03–32.84) | 26.27 (20.95–32.84) | 27.10 (18.56–43.83) |
| Genes | RUNX1 mRNA Jejunal R.E. | |
|---|---|---|
| rho | p Value | |
| TLR4 mRNA jejunal R.E. | 0.421 | 0.001 |
| TLR5 mRNA jejunal R.E. | 0.302 | 0.031 |
| PPARγ mRNA jejunal R.E. | 0.352 | 0.008 |
| SCORE | Proportion | Distribution |
|---|---|---|
| 1 | ↓ | scattered |
| 2 | ↓ | clustered |
| 3 | ↑ | clustered |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertran, L.; Pastor, A.; Portillo-Carrasquer, M.; Binetti, J.; Aguilar, C.; Martínez, S.; Vives, M.; Sabench, F.; Porras, J.A.; Riesco, D.; et al. The Potential Protective Role of RUNX1 in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2021, 22, 5239. https://doi.org/10.3390/ijms22105239
Bertran L, Pastor A, Portillo-Carrasquer M, Binetti J, Aguilar C, Martínez S, Vives M, Sabench F, Porras JA, Riesco D, et al. The Potential Protective Role of RUNX1 in Nonalcoholic Fatty Liver Disease. International Journal of Molecular Sciences. 2021; 22(10):5239. https://doi.org/10.3390/ijms22105239
Chicago/Turabian StyleBertran, Laia, Angela Pastor, Marta Portillo-Carrasquer, Jessica Binetti, Carmen Aguilar, Salomé Martínez, Margarita Vives, Fàtima Sabench, José Antonio Porras, David Riesco, and et al. 2021. "The Potential Protective Role of RUNX1 in Nonalcoholic Fatty Liver Disease" International Journal of Molecular Sciences 22, no. 10: 5239. https://doi.org/10.3390/ijms22105239