
Atovaquone Suppresses Triple-Negative Breast Tumor Growth by Reducing Immune-Suppressive Cells


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Atovaquone Inhibits Breast Tumor Growth
2.2. Reduction in Myeloid-Derived Suppressor Cells (MDSCs) with Atovaquone Treatment
2.3. Reduction of Regulatory T Cells (Tregs) by Atovaquone Treatment
2.4. Atovaquone Suppressed Tumor Cytokines
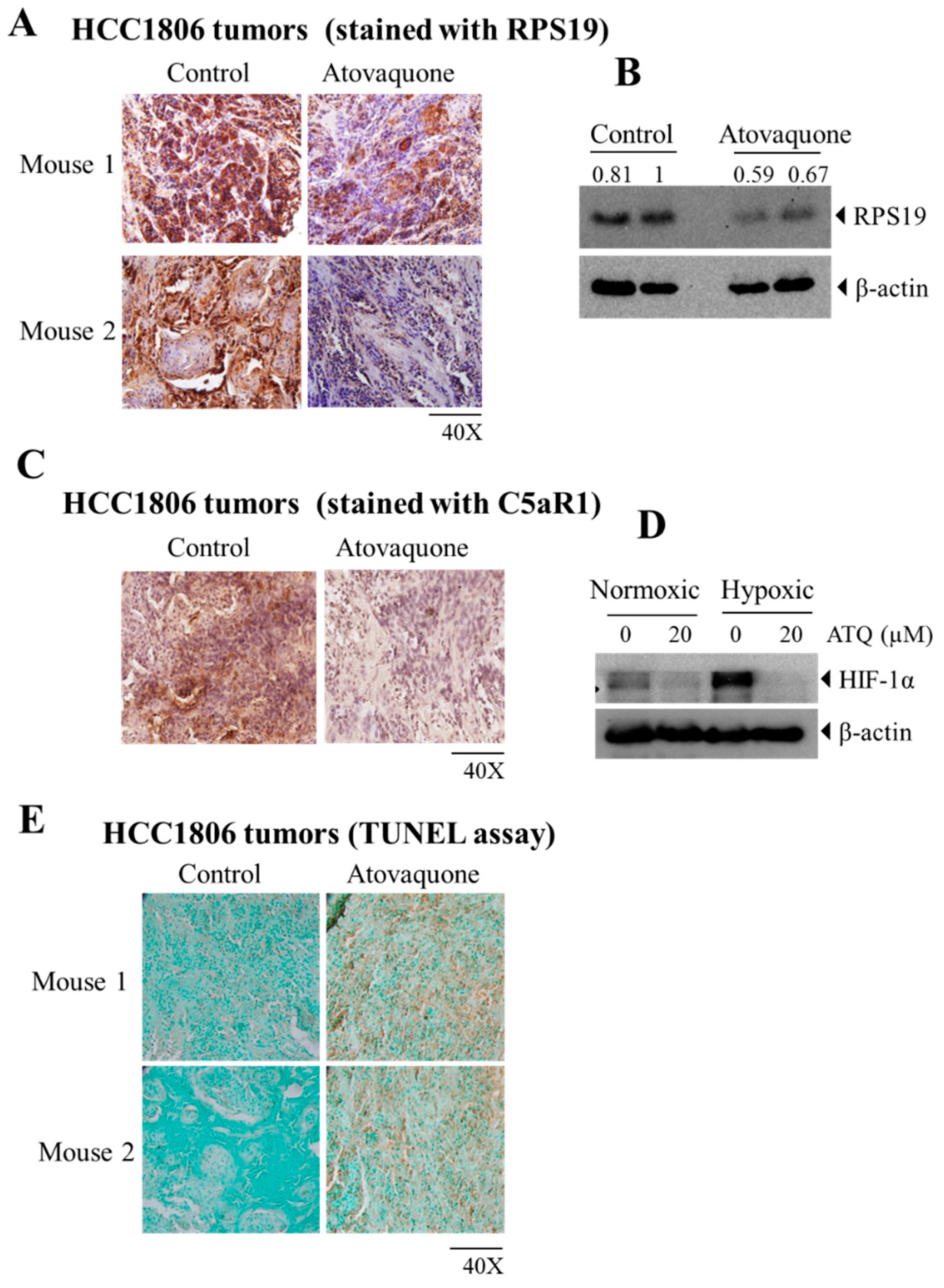
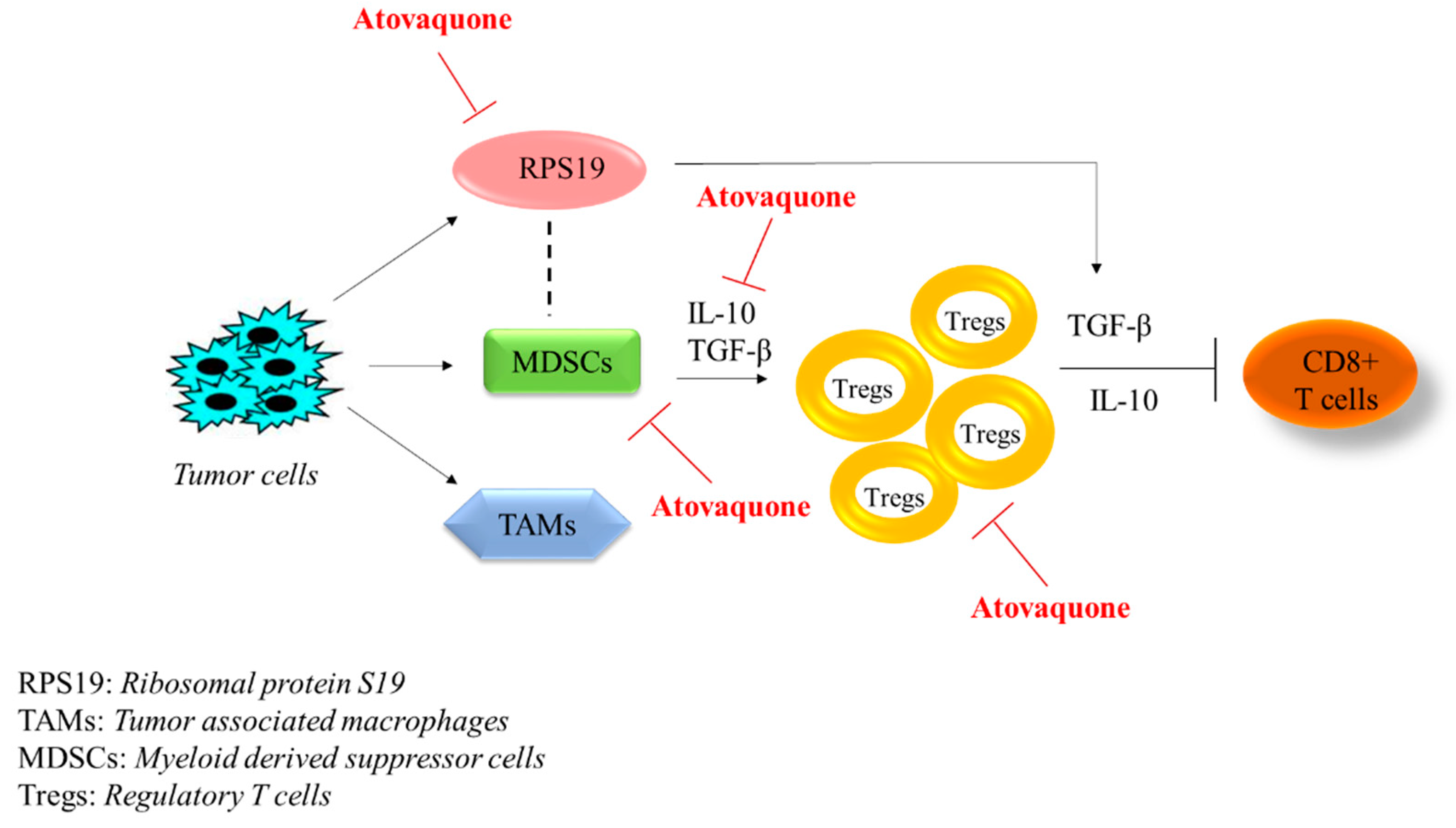
2.5. Atovaquone Reduces the Expression of the Immunosuppressive Ribosomal Protein S19 (RPS19) and Induces Apoptosis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents and Chemicals
4.3. Human PBMCs Isolation from Buffy Coat
4.4. Tumor Therapy Model
4.5. PBMC Collection from Mouse Blood and Fluorescence Cytometric Analysis
4.6. Enzyme-Linked Immune Sorbent Assay
4.7. Immunostaining for Tissue Sections
4.8. Hypoxia Treatment and Western Blot Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anders, C.K.; Carey, L.A. Biology, Metastatic Patterns, and Treatment of Patients with Triple-negative Breast Cancer. Clin. Breast Cancer 2009, 9, S73–S81. [Google Scholar] [CrossRef]
- Pareja, F.; Geyer, F.C.; Marchiò, C.; Burke, K.A.; Weigelt, B.; Reis-Filho, J.S. Triple-negative Breast Cancer: The Importance of Molecular and Histologic Subtyping, and Recognition of Low-grade Variants. NPJ Breast Cancer 2016, 2, 16036. [Google Scholar] [CrossRef] [PubMed]
- Mathe, A.; Wong-Brown, M.; Morten, B.; Forbes, J.F.; Braye, S.G.; Avery-Kiejda, K.A.; Scott, R.J. Novel Genes Associated with Lymph Node Metastasis in Triple Negative Breast Cancer. Sci. Rep. 2015, 5, 15832. [Google Scholar] [CrossRef]
- Stagg, J.; Allard, B. Immunotherapeutic Approaches in Triple-negative Breast Cancer: Latest Research and Clinical Prospects. Ther. Adv. Med Oncol. 2013, 5, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wein, L.; Loi, S. Mechanisms of Resistance of Chemotherapy in Early-stage Triple Negative Breast Cancer (TNBC). Breast 2017, 34, S27–S30. [Google Scholar] [CrossRef]
- Salimi, M. Future of Triple Negative Breast Cancer: Can Immunotherapy Treat This Deadly Subtype of Breast Cancer? Iran. Biomed. J. 2018, 22, 76. [Google Scholar]
- Katz, H.; Alsharedi, M. Immunotherapy in Triple-negative Breast Cancer. Med Oncol. 2018, 35, 13. [Google Scholar] [CrossRef]
- Yu, L.-Y.; Tang, J.; Zhang, C.-M.; Zeng, W.-J.; Yan, H.; Li, M.-P.; Chen, X.-P. New Immunotherapy Strategies in Breast Cancer. Int. J. Environ. Res. Public Health 2017, 14, 68. [Google Scholar] [CrossRef]
- Solito, S.; Pinton, L.; Mandruzzato, S. In Brief: Myeloid-derived Suppressor Cells in Cancer. J. Pathol. 2017, 242, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Marvel, D.; Gabrilovich, D.I. Myeloid-derived Suppressor Cells in the Tumor Microenvironment: Expect the Unexpected. J. Clin. Investig. 2015, 125, 3356–3364. [Google Scholar] [CrossRef] [PubMed]
- Ouzounova, M.; Lee, E.; Piranlioglu, R.; El Andaloussi, A.; Kolhe, R.; Demirci, M.F.; Marasco, D.; Asm, I.; Chadli, A.; Hassan, K.A. Monocytic and Granulocytic Myeloid Derived Suppressor Cells Differentially Regulate Spatiotemporal Tumour Plasticity during Metastatic Cascade. Nat. Commun. 2017, 8, 14979. [Google Scholar] [CrossRef]
- Umansky, V.; Blattner, C.; Gebhardt, C.; Utikal, J. The Role of Myeloid-derived Suppressor Cells (MDSC) in Cancer Progression. Vaccines 2016, 4, 36. [Google Scholar] [CrossRef] [PubMed]
- Shou, D.; Wen, L.; Song, Z.; Yin, J.; Sun, Q.; Gong, W. Suppressive Role of Myeloid-derived Suppressor Cells (MDSCs) in the Microenvironment of Breast Cancer and Targeted Immunotherapies. Oncotarget 2016, 7, 64505. [Google Scholar] [CrossRef] [Green Version]
- Plitas, G.; Konopacki, C.; Wu, K.; Bos, P.D.; Morrow, M.; Putintseva, E.V.; Chudakov, D.M.; Rudensky, A.Y. Regulatory T Cells Exhibit Distinct Features in Human Breast Cancer. Immunity 2016, 45, 1122–1134. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.A.E.; Oda, J.M.M.; Amarante, M.K.; Voltarelli, J.C. Regulatory T Cells and Breast Cancer: Implications for Immunopathogenesis. Cancer Metastasis Rev. 2010, 29, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Finotello, F.; Trajanoski, Z. New Strategies for Cancer Immunotherapy: Targeting Regulatory T Cells. Genome Med. 2017, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Liao, J.; Liu, J.; Huang, D.; He, C.; Chen, F.; Yang, L.; Wu, W.; Chen, J.; Lin, L. Blocking the Recruitment of Naive CD4+ T Cells Reverses Immunosuppression in Breast Cancer. Cell Res. 2017, 27, 461. [Google Scholar] [CrossRef]
- Vander Ark, A.; Cao, J.; Li, X. TGF-β Receptors: In and beyond TGF-β Signaling. Cell. Signal. 2018, 52, 112–120. [Google Scholar] [CrossRef]
- Prasad, S.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Srivastava, S.K. Cancer Cells Stemness: A Doorstep to Targeted Therapy. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1866, 165424. [Google Scholar] [CrossRef]
- Massagué, J. TGFβ in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachman, K.E.; Park, B.H. Duel Nature of TGF-β Signaling: Tumor Suppressor vs. Tumor Promoter. Curr. Opin. Oncol. 2005, 17, 49–54. [Google Scholar] [CrossRef]
- Dennis, K.L.; Blatner, N.R.; Gounari, F.; Khazaie, K. Current Status of IL-10 and Regulatory T-cells in Cancer. Curr. Opin. Oncol. 2013, 25, 637. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.G.; Lee, H.; Gupta, N.; Ramachandran, S.; Kaushik, I.; Srivastava, S.; Kim, S.-H.; Srivastava, S.K. Role of Forkhead Box Class O Proteins in Cancer Progression and Metastasis. Semin. Cancer Biol. 2018, 50, 142–151. [Google Scholar] [CrossRef]
- Mannino, M.H.; Zhu, Z.; Xiao, H.; Bai, Q.; Wakefield, M.R.; Fang, Y. The Paradoxical Role of IL-10 in Immunity and Cancer. Cancer Lett. 2015, 367, 103–107. [Google Scholar] [CrossRef]
- Park, M.-J.; Lee, S.-H.; Kim, E.-K.; Lee, E.-J.; Baek, J.-A.; Park, S.-H.; Kwok, S.-K.; Cho, M.-L. Interleukin-10 Produced by Myeloid-derived Suppressor Cells is Critical for the Induction of Tregs and Attenuation of Rheumatoid Inflammation in Mice. Sci. Rep. 2018, 8, 3753. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Pan, P.-Y.; Li, Q.; Sato, A.I.; Levy, D.E.; Bromberg, J.; Divino, C.M.; Chen, S.-H. Gr-1+ CD115+ Immature Myeloid Suppressor Cells Mediate the Development of Tumor-induced T Regulatory Cells and T-cell Anergy in Tumor-bearing Host. Cancer Res. 2006, 66, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Sheikhpour, E.; Noorbakhsh, P.; Foroughi, E.; Farahnak, S.; Nasiri, R.; Neamatzadeh, H. A Survey on the Role of Interleukin-10 in Breast Cancer: A Narrative. Rep. Biochem. Mol. Biol. 2018, 7, 30–37. [Google Scholar]
- Liu, C.; Wang, Q.; Sun, B.; Meng, X.; Li, L.; Yang, L.; Cong, Y.; Liu, J.; Xuan, L.; Huang, Y. Low BMI is Correlated with Increased TGF-β and IL-10 mRNA Levels in the Peripheral Blood of Breast Cancer Patients. IUBMB Life 2018, 70, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Ostapchuk, Y.O.; Cetin, E.A.; Perfilyeva, Y.V.; Yilmaz, A.; Skiba, Y.A.; Chirkin, A.P.; Omarbaeva, N.A.; Talaeva, S.G.; Belyaev, N.N.; Deniz, G. Peripheral Blood NK Cells Expressing HLA-G, IL-10 and TGF-β in Healthy Donors and Breast Cancer Patients. Cell. Immunol. 2015, 298, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-a.; Dukhanina, O.; Tang, B.; Mamura, M.; Letterio, J.J.; MacGregor, J.; Patel, S.C.; Khozin, S.; Liu, Z.-y.; Green, J. Lifetime Exposure to a Soluble TGF-β Antagonist Protects Mice against Metastasis without Adverse Side Effects. J. Clin. Investig. 2002, 109, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, R.S.; Dumont, N.; Ritter, C.A.; Dugger, T.C.; Brantley, D.M.; Chen, J.; Easterly, E.; Roebuck, L.R.; Ryan, S.; Gotwals, P.J. Blockade of TGF-β Inhibits Mammary Tumor Cell Viability, Migration, and Metastases. J. Clin. Investig. 2002, 109, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Akhurst, R.J. TGF-β Antagonists: Why Suppress a Tumor Suppressor? J. Clin. Investig. 2002, 109, 1533–1536. [Google Scholar] [CrossRef] [PubMed]
- Neuzillet, C.; Tijeras-Raballand, A.; Cohen, R.; Cros, J.; Faivre, S.; Raymond, E.; de Gramont, A. Targeting the TGFβ Pathway for Cancer Therapy. Pharmacol. Ther. 2015, 147, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiura, H.; Shibuya, Y.; Yamamoto, T. S19 Ribosomal Protein Cross-linked Dimer Causes Monocyte-predominant Infiltration by Means of Molecular Mimicry to Complement C5a. Lab. Investig. J. Tech. Methods Pathol. 1998, 78, 1615–1623. [Google Scholar]
- Markiewski, M.M.; DeAngelis, R.A.; Benencia, F.; Ricklin-Lichtsteiner, S.K.; Koutoulaki, A.; Gerard, C.; Coukos, G.; Lambris, J.D. Modulation of the Antitumor Immune Response by Complement. Nat. Immunol. 2008, 9, 1225. [Google Scholar] [CrossRef] [Green Version]
- Markiewski, M.M.; Lambris, J.D. Unwelcome Complement. Cancer Res. 2009, 69, 6367–6370. [Google Scholar] [CrossRef] [Green Version]
- Safaeian, M.; Hildesheim, A.; Gonzalez, P.; Yu, K.; Porras, C.; Li, Q.; Rodriguez, A.C.; Sherman, M.E.; Schiffman, M.; Wacholder, S. Single Nucleotide Polymorphisms in the PRDX3 and RPS19 and Risk of HPV Persistence and Cervical Precancer/Cancer. PLoS ONE 2012, 7, e33619. [Google Scholar] [CrossRef] [Green Version]
- Chien, C.-C.; Tu, T.-C.; Huang, C.-J.; Yang, S.-H.; Lee, C.-L. Lowly Expressed Ribosomal Protein s19 in the Feces of Patients with Colorectal Cancer. ISRN Gastroenterol. 2012, 2012, 394545. [Google Scholar] [CrossRef] [Green Version]
- Markiewski, M.M.; Vadrevu, S.K.; Sharma, S.K.; Chintala, N.K.; Ghouse, S.; Cho, J.-H.; Fairlie, D.P.; Paterson, Y.; Astrinidis, A.; Karbowniczek, M. The Ribosomal Protein S19 Suppresses Antitumor Immune Responses via the Complement C5a Receptor 1. J. Immunol. 2017, 198, 2989–2999. [Google Scholar] [CrossRef] [Green Version]
- Labiano, S.; Palazon, A.; Melero, I. Immune Response Regulation in the Tumor Microenvironment by Hypoxia. Semin. Oncol. 2015, 42, 378–386. [Google Scholar] [CrossRef]
- Triner, D.; Shah, Y.M. Hypoxia-inducible Factors: A Central Link between Inflammation and Cancer. J. Clin. Investig. 2016, 126, 3689–3698. [Google Scholar] [CrossRef]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1α and HIF2α: Sibling Rivalry in Hypoxic Tumour Growth and Progression. Nat. Rev. Cancer 2012, 12, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Noman, M.Z.; Hasmim, M.; Messai, Y.; Terry, S.; Kieda, C.; Janji, B.; Chouaib, S. Hypoxia: A Key Player in Antitumor Immune Response. A Review in the Theme: Cellular Responses to Hypoxia. Am. J. Physiol. Cell Physiol. 2015, 309, C569–C579. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Gabrilovich, D.I. Hypoxia-inducible Factors in Regulation of Immune Responses in Tumour Microenvironment. Immunology 2014, 143, 512–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Z.; Yan, X.; Lu, L.; Su, C.; He, Y. Atovaquone Enhances Doxorubicin’s Efficacy via Inhibiting Mitochondrial Respiration and STAT3 in Aggressive Thyroid Cancer. J. Bioenerg. Biomembr. 2018, 50, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, X.; Shan, W.; Liu, Q.; Wang, C.; Zheng, J.; Yao, H.; Tang, R.; Zheng, J. Anti-malarial Atovaquone Exhibits Anti-tumor Effects by Inducing DNA Damage in Hepatocellular Carcinoma. Am. J. Cancer Res. 2018, 8, 1697. [Google Scholar]
- Xiang, M.; Kim, H.; Ho, V.T.; Walker, S.R.; Bar-Natan, M.; Liu, S.; Toniolo, P.A.; Kroll, Y.; Jones, N.; Giaccone, Z.T. Gene Expression-based Discovery of Atovaquone as a STAT3 Inhibitor and Anti-cancer Agent. Blood 2016, 128, 1845–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, T.M.; Fokas, E.; Kunz-Schughart, L.A.; Folkes, L.K.; Anbalagan, S.; Huether, M.; Kelly, C.J.; Pirovano, G.; Buffa, F.M.; Hammond, E.M. The Anti-malarial Atovaquone Increases Radiosensitivity by Alleviating Tumour Hypoxia. Nat. Commun. 2016, 7, 12308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Srivastava, S.K. Atovaquone: An Antiprotozoal Drug Suppresses Primary and Resistant Breast Tumor Growth by Inhibiting HER2/β-Catenin Signaling. Mol. Cancer Ther. 2019, 18, 1708–1720. [Google Scholar] [CrossRef]
- Gupta, N.; Gupta, P.; Srivastava, S.K. Penfluridol Overcomes Paclitaxel Resistance in Metastatic Breast Cancer. Sci Rep. 2019, 9, 5066. [Google Scholar] [CrossRef]
- Goedegebuure, P.; Mitchem, B.J.; Porembka, R.M.; Tan, C.B.M.; Belt, B.A.; Wang-Gillam, A.; Gillanders, E.W.; Hawkins, G.W.; Linehan, C.D. Myeloid-derived Suppressor Cells: General Characteristics and Relevance to Clinical Management of Pancreatic Cancer. Curr. Cancer Drug Targets 2011, 11, 734–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.; Tanikawa, T.; Li, W.; Zhao, L.; Vatan, L.; Szeliga, W.; Wan, S.; Wei, S.; Wang, Y.; Liu, Y.; et al. Myeloid-Derived Suppressor Cells Endow Stem-like Qualities to Breast Cancer Cells through IL6/STAT3 and NO/NOTCH Cross-talk Signaling. Cancer Res. 2016, 76, 3156–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowitz, J.; Wesolowski, R.; Papenfuss, T.; Brooks, T.R.; Carson, W.E. Myeloid-derived Suppressor Cells in Breast Cancer. Breast Cancer Res. Treat. 2013, 140, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Vikas, P.; Borcherding, N.; Zhang, W. The Clinical Promise of Immunotherapy in Triple-negative Breast Cancer. Cancer Manag. Res. 2018, 10, 6823–6833. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Qi, Y.; Kong, X.; Zhai, J.; Li, Y.; Song, Y.; Wang, J.; Feng, X.; Fang, Y. Immunological Therapy: A Novel Thriving Area for Triple-negative Breast Cancer Treatment. Cancer Lett. 2019, 442, 409–428. [Google Scholar] [CrossRef]
- Marrufo, A.M.; Mathew, S.O.; Chaudhary, P.; Malaer, J.D.; Vishwanatha, J.K.; Mathew, P.A. Blocking LLT1 (CLEC2D, OCIL)-NKRP1A (CD161) Interaction Enhances Natural Killer Cell-mediated Lysis of Triple-negative Breast Cancer Cells. Am. J. Cancer Res. 2018, 8, 1050–1063. [Google Scholar]
- Pan, P.-Y.; Ma, G.; Weber, K.J.; Ozao-Choy, J.; Wang, G.; Yin, B.; Divino, C.M.; Chen, S.-H. Immune Stimulatory Receptor CD40 is Required for T-cell Suppression and T Regulatory Cell Activation Mediated by Myeloid-derived Suppressor Cells in Cancer. Cancer Res. 2010, 70, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Wang, Y.; Yan, F.; Zhang, P.; Li, H.; Zhao, H.; Yan, C.; Yan, F.; Ren, X. Noncanonical NF-κB Activation Mediates STAT3-stimulated IDO Upregulation in Myeloid-derived Suppressor Cells in Breast Cancer. J. Immunol. 2014, 193, 2574–2586. [Google Scholar] [CrossRef] [Green Version]
- Mace, T.A.; King, S.A.; Ameen, Z.; Elnaggar, O.; Young, G.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K.; Knobloch, T.J.; Weghorst, C.M. Bioactive Compounds or Metabolites from Black Raspberries Modulate T Lymphocyte Proliferation, Myeloid Cell Differentiation and Jak/STAT Signaling. Cancer Immunol. Immunother. 2014, 63, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Danishmalik, S.N.; Sin, J.-I. Therapeutic Tumor Control of HER2 DNA Vaccines is Achieved by an Alteration of Tumor Cells and Tumor Microenvironment by Gemcitabine and Anti-Gr-1 Ab Treatment in a HER2-expressing Tumor Model. DNA Cell Biol. 2017, 36, 801–811. [Google Scholar] [CrossRef]
- Wesolowski, R.; Markowitz, J.; Carson, W.E. Myeloid Derived Suppressor Cells–A New Therapeutic Target in the Treatment of Cancer. J. Immunother. Cancer 2013, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The Nature of Myeloid-derived Suppressor Cells in the Tumor Microenvironment. Trends Immunol. 2016, 37, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Kapanadze, T.; Gamrekelashvili, J.; Ma, C.; Chan, C.; Zhao, F.; Hewitt, S.; Zender, L.; Kapoor, V.; Felsher, D.W.; Manns, M.P. Regulation of Accumulation and Function of Myeloid Derived Suppressor Cells in Different Murine Models of Hepatocellular Carcinoma. J. Hepatol. 2013, 59, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Ostrand-Rosenberg, S.; Sinha, P. Myeloid-derived Suppressor Cells: Linking Inflammation and Cancer. J. Immunol. 2009, 182, 4499–4506. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Varney, M.L.; Saxena, S.; Wu, L.; Singh, R.K. Induction of CXCR2 Ligands, Stem Cell-like Phenotype, and Metastasis in Chemotherapy-resistant Breast Cancer Cells. Cancer Lett. 2016, 372, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Younos, I.; Donkor, M.; Hoke, T.; Dafferner, A.; Samson, H.; Westphal, S.; Talmadge, J. Tumor-and Organ-dependent Infiltration by Myeloid-derived Suppressor Cells. Int. Immunopharmacol. 2011, 11, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.; Gupta, N.; Zang, T.; Wangtrakluldee, P.; Srivastava, S.K.; Penning, T.M.; Trippier, P.C. AKR1C3 Inhibitor KV-37 Exhibits Antineoplastic Effects and Potentiates Enzalutamide in Combination Therapy in Prostate Adenocarcinoma Cells. Mol. Cancer Ther. 2018, 17, 1833–1845. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Wright, S.E.; Srivastava, S.K. PEITC Treatment Suppresses Myeloid Derived Tumor Suppressor Cells to Inhibit Breast Tumor Growth. Oncoimmunology 2015, 4, e981449. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Wright, S.; Srivastava, S.K. Immune Consequences of Penfluridol Treatment Associated with Inhibition of Glioblastoma Tumor Growth. Oncotarget 2017, 8, 47632. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Gupta, N.; Fofaria, N.M.; Ranjan, A.; Srivastava, S.K. HER2-mediated GLI2 Stabilization Promotes Anoikis Resistance and Metastasis of Breast Cancer Cells. Cancer Lett. 2019, 442, 68–81. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, N.; Gaikwad, S.; Kaushik, I.; Wright, S.E.; Markiewski, M.M.; Srivastava, S.K. Atovaquone Suppresses Triple-Negative Breast Tumor Growth by Reducing Immune-Suppressive Cells. Int. J. Mol. Sci. 2021, 22, 5150. https://doi.org/10.3390/ijms22105150
Gupta N, Gaikwad S, Kaushik I, Wright SE, Markiewski MM, Srivastava SK. Atovaquone Suppresses Triple-Negative Breast Tumor Growth by Reducing Immune-Suppressive Cells. International Journal of Molecular Sciences. 2021; 22(10):5150. https://doi.org/10.3390/ijms22105150
Chicago/Turabian StyleGupta, Nehal, Shreyas Gaikwad, Itishree Kaushik, Stephen E. Wright, Maciej M. Markiewski, and Sanjay K. Srivastava. 2021. "Atovaquone Suppresses Triple-Negative Breast Tumor Growth by Reducing Immune-Suppressive Cells" International Journal of Molecular Sciences 22, no. 10: 5150. https://doi.org/10.3390/ijms22105150