Preparing for the KIL: Receptor Analysis of Pseudomonas syringae pv. porri Phages and Their Impact on Bacterial Virulence

 ,
,
Abstract
:1. Introduction
2. Results
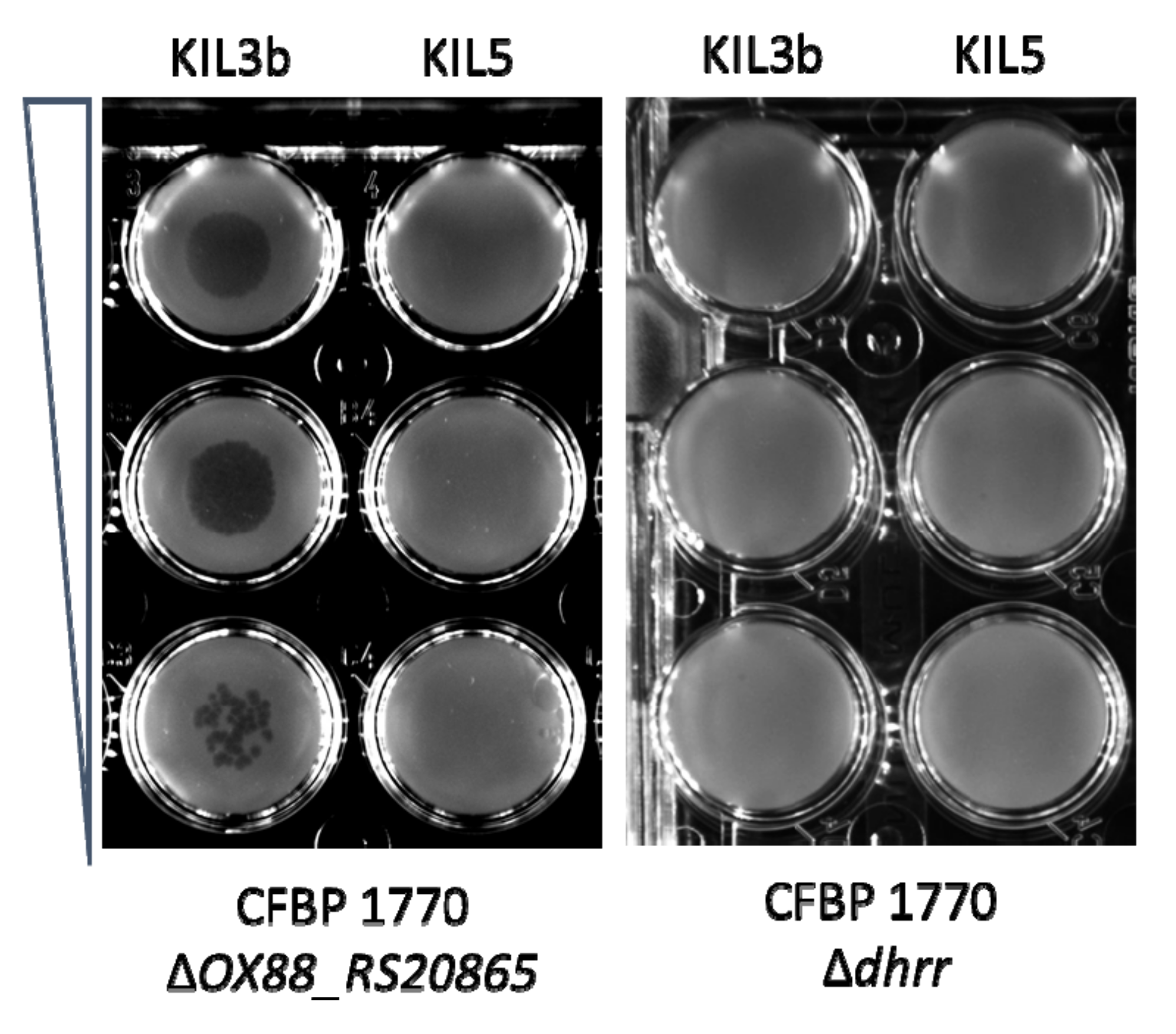
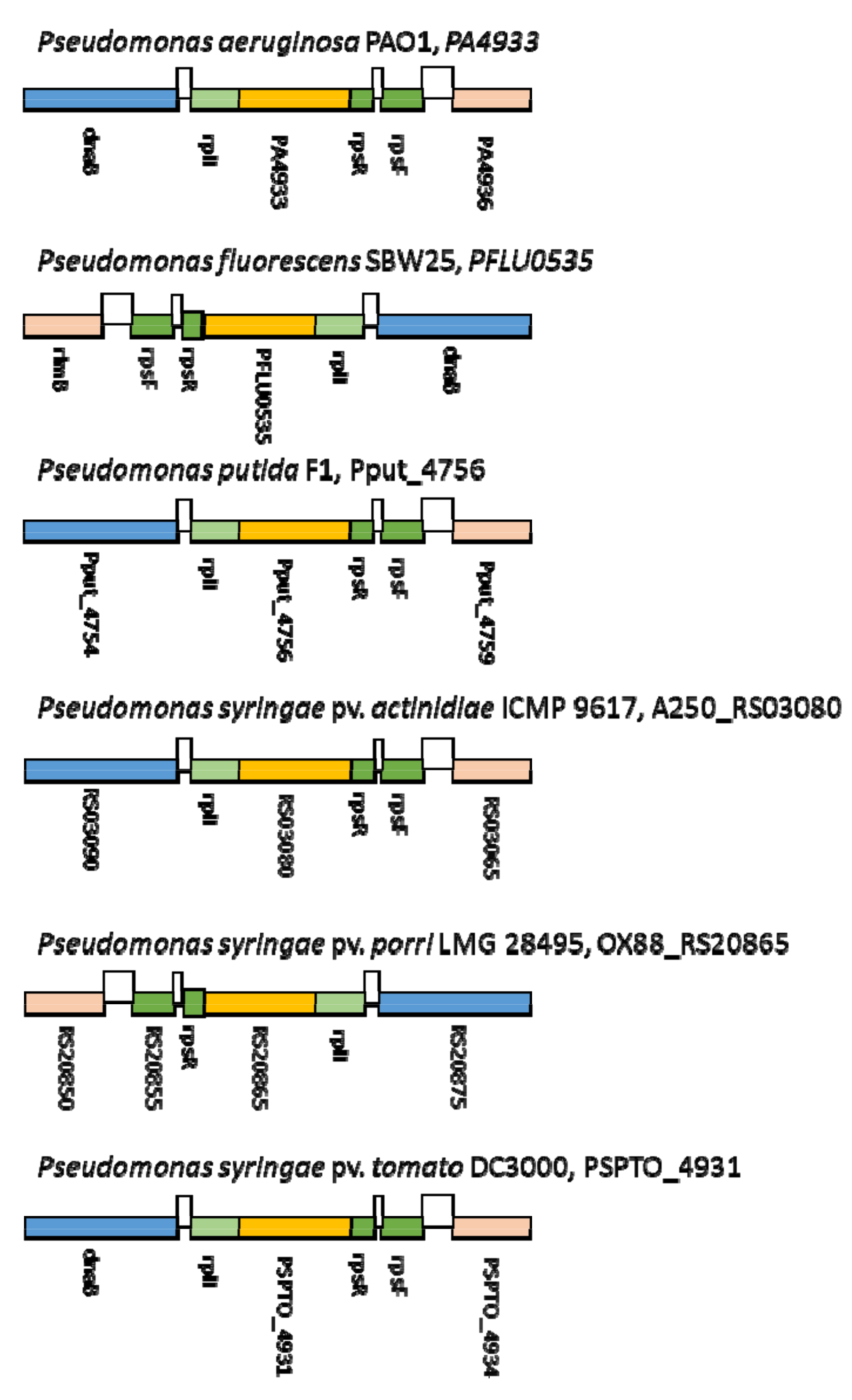
2.1. Subsection Identification of Genes Involved in the Attachment of KIL3b and KIL5 to Pseudomonas syringae pv. porri
2.2. Phage-Resistant Pspo Mutants Display a Markedly Reduced Virulence
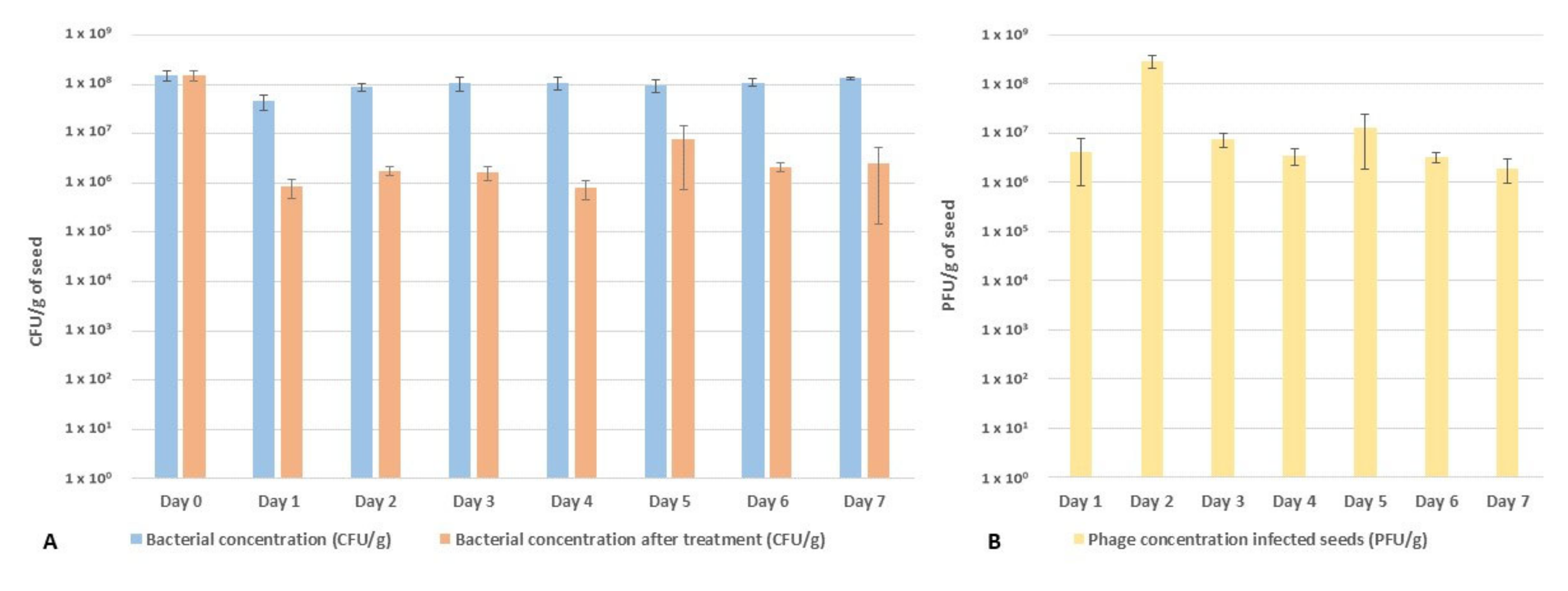
2.3. Performance of the Phage Cocktail in Reducing Pspo Titers during Seed Priming
2.4. Screening and Full Genome SNP Analysis of Phage-Resistant Pspo Strains
3. Discussion
3.1. Elucidation of the Differences in Bacterial Receptors for Phages KIL3b and KIL5
3.2. Impact of the Bacterial Receptors on Phage-Based Biocontrol Assays
4. Materials and Methods
4.1. Microbiological Manipulations
4.2. Tn5 Knock-Out Mutants of CFBP 1770 and Selection of Phage-Resistant Clones
4.3. Complementation of Phage Resistance and Phage Adsorption Assay
4.4. Virulence Assay in Leek Plants
4.5. Seed Bioassay
4.6. Sequencing of Natural Phage-Resistant Clones of Pseudomonas syringae pv. porri
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage | KO Mutant | Protein | Accession Number | e-Value |
|---|---|---|---|---|
| KIL3b | CFBP 1770_R3a | dTDP-4-dehydrorhamnose reductase | KPY21988.1 | 6 × 10−144 |
| CFBP 1770_R3b | Carbamoyltransferase | KOP57525.1 | 9 × 10−75 | |
| CFBP 1770_R3c | dTDP-4-dehydrorhamnose reductase | KOP54965.1 | 2 × 10−98 | |
| CFBP 1770_R3d | Carbamoyltransferase | KOP57525.1 | 9 × 10−75 | |
| CFBP 1770_R3e | Carbamoyltransferase | KOP57525.1 | 8 × 10−75 | |
| CFBP 1770_R3f | dTDP-glucose 4,6-dehydratase | KOP54966.1 | 3 × 10−108 | |
| CFBP 1770_R3g | GDP-6-deoxy-D-lyxo-4-hexulose reductase | KPY25409.1 | 2 × 10−8 | |
| KIL5 | CFBP 1770_R5a | Hypothetical protein ALP22_101621 | RMU82419.1 | 5 × 10−91 |
| CFBP 1770_R5b | Hypothetical membrane protein | KOP53361.1 | 4 × 10−57 | |
| CFBP 1770_R5c | Hypothetical membrane protein | KOP53361.1 | 5 × 10−50 | |
| CFBP 1770_R5d | Hypothetical membrane protein | KOP53361.1 | 6 × 10−64 |
References
- Mallmann, W.; Hemstreet, C. Isolation ofan inhibitory substance from plants. Agric. Res. 1924, 28, 599–602. [Google Scholar]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtappels, D.; Lavigne, R.; Huys, I.; Wagemans, J. Protection of Phage Applications in Crop Production: A Patent Landscape. Viruses 2019, 11, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellington, E.M.; Boxall, A.B.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Barra Caracciolo, A. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Bondarczuk, K.; Piotrowska-Seget, Z. Molecular basis of active copper resistance mechanisms in Gram-negative bacteria. Cell Biol. Toxicol. 2013, 29, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.K.; Király, L.; Schwarczinger, I. Phage therapy for plant disease control with a focus on fire blight. Cent. Eur. J. Biol. 2011, 7, 1–12. [Google Scholar] [CrossRef]
- Monteil, C.L.; Yahara, K.; Studholme, D.J.; Mageiros, L.; Méric, G.; Swingle, B.; Morris, C.E.; Vinatzer, B.A.; Sheppard, S.K. Population-genomic insights into emergence, crop adaptation and dissemination of Pseudomonas syringae pathogens. Microb. Genom. 2016, 2, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lelliott, R.A. A New Bacterial Disease of Leeks. Plant Pathol. 1952, 1, 84. [Google Scholar] [CrossRef]
- Rombouts, S.; Van Vaerenbergh, J.; Volckaert, A.; Baeyen, S.; De Langhe, T.; Declercq, B.; Lavigne, R.; Maes, M. Isolation and characterization of Pseudomonas syringae pv. porri from leek in Flanders. Eur. J. Plant Pathol. 2016, 144, 185–198. [Google Scholar] [CrossRef]
- Samson, R.; Shafik, H.; Benjama, A.; Gardan, L. Description of the bacterium causing blight of leek as Pseudomonas syringae pv. porri (pv. nov.). Phytopathology 1998, 88, 844–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, D.H.; Cother, E.J.; Hailstones, D.L.; Flack, M.; Oxspring, L.; Hall, B. Characterisation of Pseudomonas syringae strains associated with a leaf disease of leek in Australia. Eur. J. Plant Pathol. 2006, 115, 419–430. [Google Scholar] [CrossRef]
- Rombouts, S.; Volckaert, A.; Venneman, S.; Declercq, B.; Vandenheuvel, D.; Allonsius, C.N.; Van Malderghem, C.; Jang, H.B.; Briers, Y.; Noben, J.P.; et al. Characterization of novel bacteriophages for biocontrol of bacterial blight in leek caused by Pseudomonas syringae pv. porri. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.; Ward, S.; Hyman, P. More is better: Selecting for broad host range bacteriophages. Front. Microbiol. 2016, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hyman, P.; Abedon, S.T. Bacteriophage Host Range and Bacterial Resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, M.; Bai, J.; Lim, J.-A.; Heu, S.; Ryu, S. Colanic Acid Is a Novel Phage Receptor of Pectobacterium carotovorum subsp. carotovorum Phage POP72. Front. Microbiol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Roach, D.R.; Sjaarda, D.R.; Castle, A.J.; Svircev, A.M. Host exopolysaccharide quantity and composition impact erwinia: Amylovora bacteriophage pathogenesis. Appl. Environ. Microbiol. 2013, 79, 3249–3256. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.-H.; Wu, H.-C.; Tseng, Y.-H. Mutation in the Xanthomonas campestris xanA Gene Required for Synthesis of Xanthan and Lipopolysaccharide Drastically Reduces the Efficiency of Bacteriophage φL7 Adsorption. Biochem. Biophys. Res. Commun. 2002, 291, 338–343. [Google Scholar] [CrossRef]
- Narulita, E.; Addy, H.S.; Kawasaki, T.; Fujie, M.; Yamada, T. The involvement of the PilQ secretin of type IV pili in phage infection in Ralstonia solanacearum. Biochem. Biophys. Res. Commun. 2016, 469, 868–872. [Google Scholar] [CrossRef]
- Epicentre EZ-Tn5TM <KAN-2> Tnp TransposomeTM Kit. 2012, pp. 1–9. Available online: https://www.lucigen.com/docs/manuals/MA138E-EZ-Tn5-KAN-2-Transposome.pdf (accessed on 20 April 2020).
- Zdorovenko, E.L.; Zatonskii, G.V.; Kocharova, N.A.; Shashkov, A.S.; Knirel, Y.A.; Ovod, V.V. Structure of the O polysaccharides and serological classification of Pseudomonas syringae pv. porri from genomospecies 4. Eur. J. Biochem. 2003, 270, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskella, B.; Brockhurst, M.A. Bacteria–phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Creuzenet, C.; Lam, J.S. Topological and functional characterization of WbpM, an inner membrane UDP-GlcNAc C6 dehydratase essential for lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Mol. Microbiol. 2001, 41, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Schoonejans, E.; Expert, D.; Toussaint, A. Characterization and virulence properties of Erwinia chrysanthemi lipopolysaccharide-defective, phi EC2-resistant mutants. J. Bacteriol. 1987, 169, 4011–4017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumrall, E.T.; Shen, Y.; Keller, A.P.; Rismondo, J.; Pavlou, M.; Eugster, M.R.; Boulos, S.; Disson, O.; Thouvenot, P.; Kilcher, S.; et al. Phage resistance at the cost of virulence: Listeria monocytogenes serovar 4b requires galactosylated teichoic acids for InlB-mediated invasion. PLoS Pathog. 2019, 15, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippov, A.A.; Sergueev, K.V.; He, Y.; Huang, X.-Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.M.; Nikolich, M.P. Bacteriophage-Resistant Mutants in Yersinia pestis: Identification of Phage Receptors and Attenuation for Mice. PLoS ONE 2011, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Olszak, T.; Danis-Wlodarczyk, K.; Arabski, M.; Gula, G.; Maciejewska, B.; Wasik, S.; Lood, C.; Higgins, G.; Harvey, B.J.; Lavigne, R.; et al. Pseudomonas aeruginosa PA5oct jumbo phage impacts planktonic and biofilm population and reduces its host virulence. Viruses 2019, 11, 1089. [Google Scholar] [CrossRef] [Green Version]
- Cumby, N.; Reimer, K.; Mengin-Lecreulx, D.; Davidson, A.R.; Maxwell, K.L. The phage tail tape measure protein, an inner membrane protein and a periplasmic chaperone play connected roles in the genome injection process of E.coli phage HK97. Mol. Microbiol. 2015, 96, 437–447. [Google Scholar] [CrossRef]
- Williams, N.; Fox, D.K.; Shea, C.; Roseman, S. Pel, the protein that permits lambda DNA penetration of Escherichia coli, is encoded by a gene in ptsM and is required for mannose utilization by the phosphotransferase system. Proc. Natl. Acad. Sci. USA 1986, 83, 8934–8938. [Google Scholar] [CrossRef] [Green Version]
- Scandella, D.; Arber, W. Phage λ DNA injection into Escherichia coli pel− mutants is restored by mutations in phage genes V or H. Virology 1976, 69, 206–215. [Google Scholar] [CrossRef]
- Torres-Barceló, C. Phage Therapy Faces Evolutionary Challenges. Viruses 2018, 10, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svircev, A.; Roach, D.; Castle, A. Framing the future with bacteriophages in agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.B.; Jackson, L.E.; Balogh, B.; Obradovic, A.; Iriarte, F.B.; Momol, M.T. Bacteriophages for plant disease control. Annu. Rev. Phytopathol. 2007, 45, 245–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, Y.E.; Saleh, A.A.; Al-Saleh, M.A. Management of asiatic citrus canker under field conditions in Saudi Arabia using bacteriophages and acibenzolar-s-methyl. Plant Dis. 2017, 101, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Koike, S.T.; Barak, J.D.; Henderson, D.M.; Gilbertson, R.L. Bacterial blight of leek: A new disease in California caused by Pseudomonas syringae. Plant Dis. 1999, 83, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Wright, R.C.T.; Friman, V.P.; Smith, M.C.M.; Brockhurst, M.A. Resistance evolution against phage combinations depends on the timing and order of exposure. MBio 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Adriaenssens, E.M.; Klumpp, J.; Lavigne, R. Flaumdravirus; International committee on Taxonomy of Viruses: Washington, DC, USA, 2018. [Google Scholar]
- Wagemans, J.; Blasdel, B.G.; Van den Bossche, A.; Uytterhoeven, B.; De Smet, J.; Paeshuyse, J.; Cenens, W.; Aertsen, A.; Uetz, P.; Delattre, A.-S.; et al. Functional elucidation of antibacterial phage ORFans targeting Pseudomonas aeruginosa. Cell. Microbiol. 2014, 16, 1822–1835. [Google Scholar] [CrossRef]
- Singer, T.; Burke, E. High-Throughput TAIL-PCR as a Tool to Identify DNA Flanking Insertions. In Methods in Molecular Biology, vol. 236: Plant Functional Genomics: Methods and Protocols; Grotewold, E., Ed.; Humana Press: Totowa, NJ, USA, 2003; pp. 241–271. [Google Scholar]
- States, D.J.; Gish, W. Comined use of sequence similarity and codon bias for coding region identification. J. Comput. Biol. 1994, 1, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.; Damron, F.H.; Mima, T.; Schweizer, H.P.; Yu, H.D. PBAD-based shuttle vectors for functional analysis of toxic and highly regulated genes in Pseudomonas and Burkholderia spp. and other bacteria. Appl. Environ. Microbiol. 2008, 74, 7422–7426. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; Edwards, R.; Nash, J.H.E.; Mahadevan, P.; Seto, D.; Ackermann, H.-W.; Lavigne, R.; Kropinski, A.M. Integration of genomic and proteomic analyses in the classification of the Siphoviridae family. Virology 2015, 477, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Van der Linden, I.; Cottyn, B.; Uyttendaele, M.; Vlaemynck, G.; Maes, M.; Heyndrickx, M. Long-term survival of Escherichia coli O157:H7 and Salmonella enterica on butterhead lettuce seeds, and their subsequent survival and growth on the seedlings. Int. J. Food Microbiol. 2013, 161, 214–219. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of neural network basecalling tools for Oxford Nanopore sequencing. Genome Biol. 2019, 20, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucl. Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Snippy: Fast Bacterial Variant Calling from NGS Reads. Available online: https://github.com/tseemann/snippy (accessed on 3 March 2020).





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holtappels, D.; Kerremans, A.; Busschots, Y.; Van Vaerenbergh, J.; Maes, M.; Lavigne, R.; Wagemans, J. Preparing for the KIL: Receptor Analysis of Pseudomonas syringae pv. porri Phages and Their Impact on Bacterial Virulence. Int. J. Mol. Sci. 2020, 21, 2930. https://doi.org/10.3390/ijms21082930
Holtappels D, Kerremans A, Busschots Y, Van Vaerenbergh J, Maes M, Lavigne R, Wagemans J. Preparing for the KIL: Receptor Analysis of Pseudomonas syringae pv. porri Phages and Their Impact on Bacterial Virulence. International Journal of Molecular Sciences. 2020; 21(8):2930. https://doi.org/10.3390/ijms21082930
Chicago/Turabian StyleHoltappels, Dominique, Alison Kerremans, Yoni Busschots, Johan Van Vaerenbergh, Martine Maes, Rob Lavigne, and Jeroen Wagemans. 2020. "Preparing for the KIL: Receptor Analysis of Pseudomonas syringae pv. porri Phages and Their Impact on Bacterial Virulence" International Journal of Molecular Sciences 21, no. 8: 2930. https://doi.org/10.3390/ijms21082930