Changes in Aquaporin 1, 5 and 9 Gene Expression in the Porcine Oviduct According to Estrous Cycle and Early Pregnancy

 , , ,
, , ,
Abstract
:1. Introduction
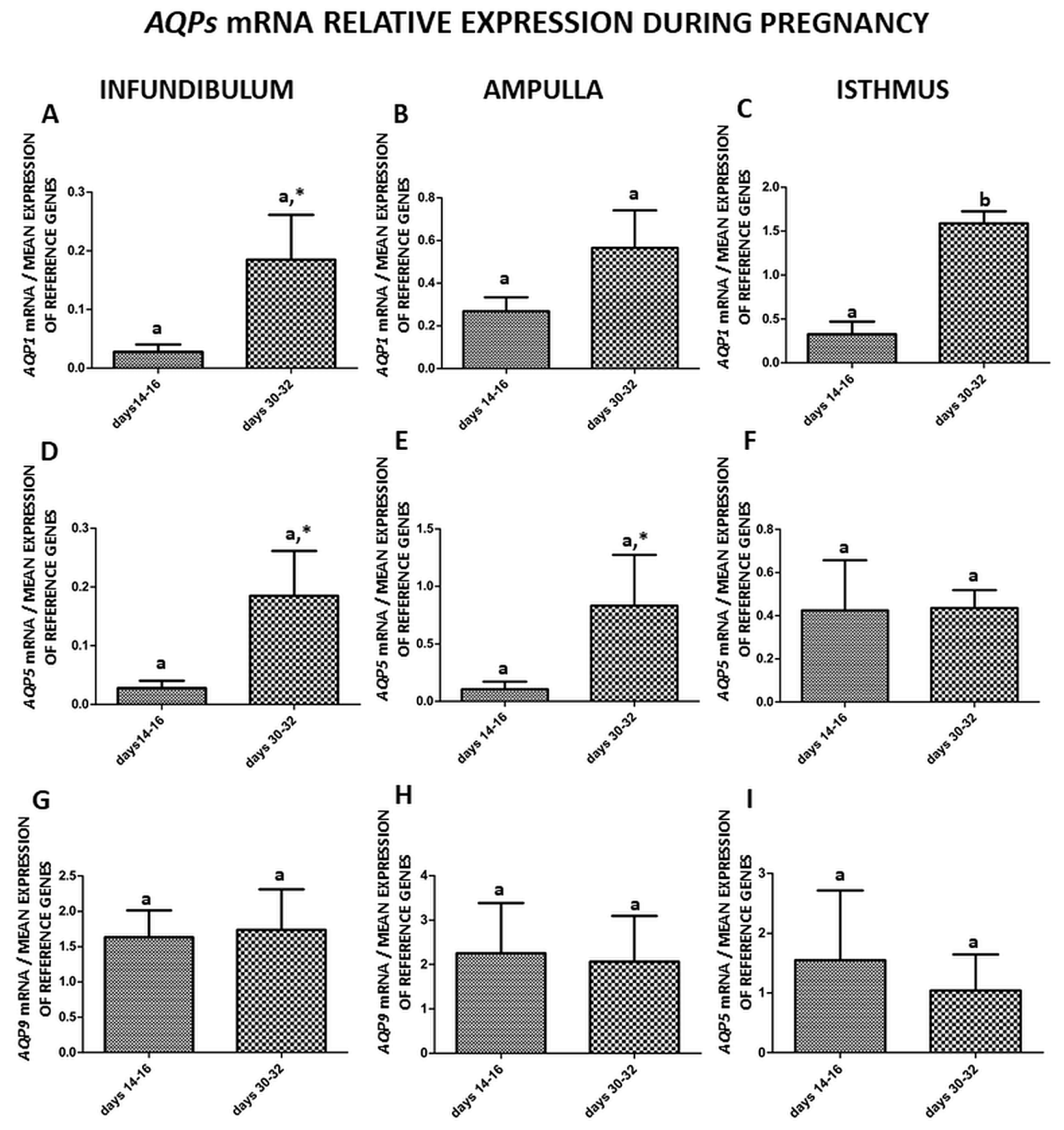
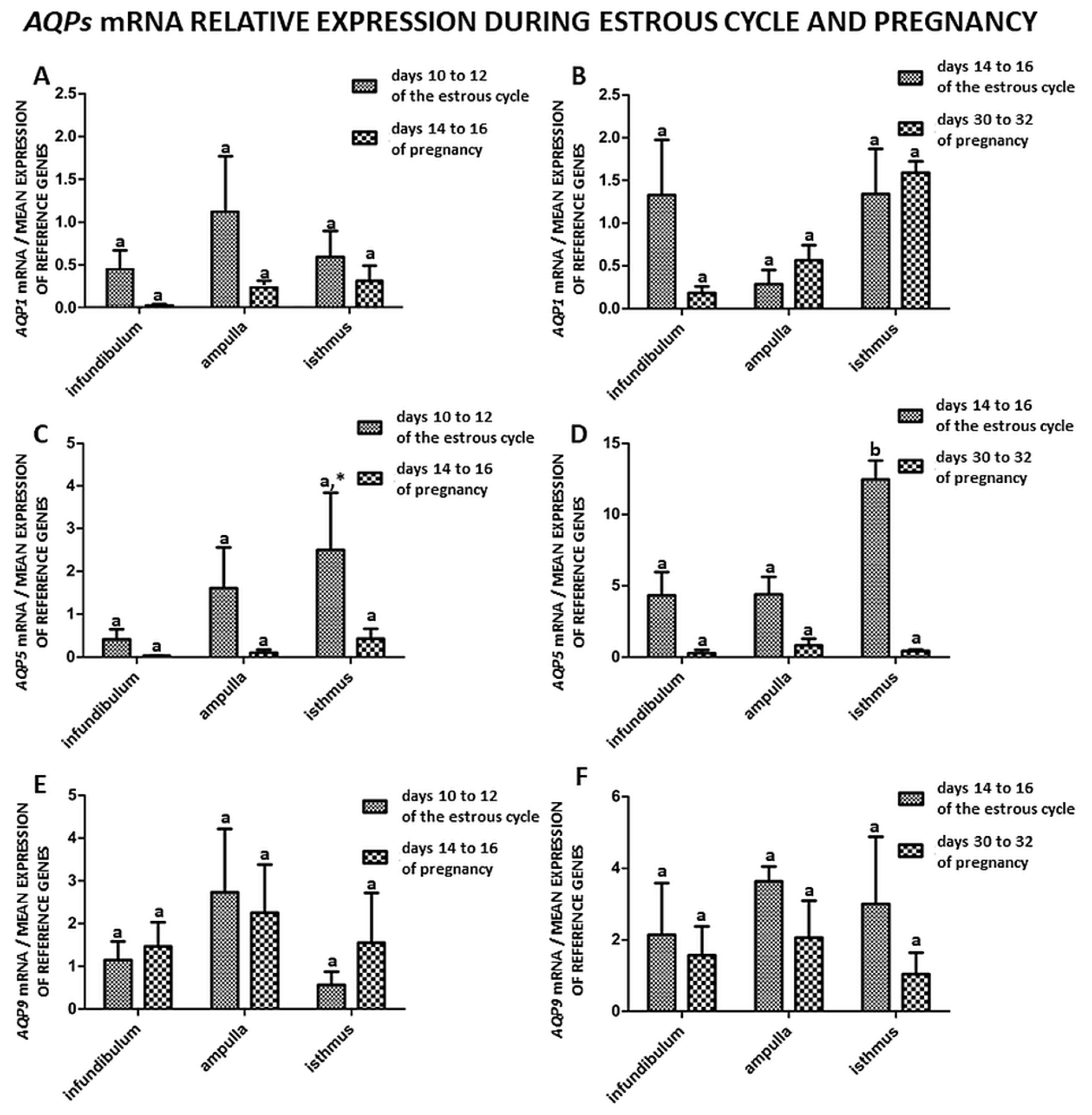
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Collection of Oviduct Tissue
4.2. Preparation of Oviduct Slices
4.3. RNA Isolation
4.4. cDNA Synthesis and Quantitative Real-Time Polymerase Chain Reaction Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pauerstein, C.J.; Eddy, C.A. The role of the oviduct in reproduction; our knowledge and our ignorance. Reproduction 1979, 55, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Black, D.L.; Davis, J.T. A blocking mechanism in the cow oviduct. J. Reprod. Fertil. 1962, 4, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johns, A.; Buchanan, J.D.; Coons, L.W. Effect of ovulation on the ionic and water content of rabbit oviduct. Biol. Reprod. 1982, 26, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verco, C.J. Fallopian tube anatomy, microanatomy, microcirculation and counter-current exchange. In The Fallopian Tube; Clinical and Surgical Aspects; Grudzinskas, J.G., Chapman, M.G., Chard, T., Eds.; Springer: London, UK, 1994; pp. 3–15. [Google Scholar]
- Buhi, W.C.; Alvarez, I.M.; Kouba, A.J. Oviductal regulation of fertilization and early embryonic development. J. Reprod. Fertil. Suppl. 1997, 52, 285–300. [Google Scholar] [PubMed]
- Brussow, K.P.; Ratky, J.; Rodriguez-Martinez, H. Fertilization and early embryonic development in the porcine fallopian tube. Reprod. Domest. Anim. Suppl. 2008, 2, 245–251. [Google Scholar] [CrossRef]
- Leese, H.; Tay, J.; Reischl, J.; Downing, S. Formation of Fallopian tubal fluid: Role of a neglected epithelium. Reproduction 2001, 121, 339–346. [Google Scholar] [CrossRef]
- Skowronski, M.T.; Kwon, T.H.; Nielsen, S. Immunolocalization of aquaporin 1, 5, and 9 in the female pig reproductive system. J. Histochem. Cytochem. 2009, 57, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Skowronski, M.T.; Skowronska, A.; Nielsen, S. Fluctuation of aquaporin 1, 5, and 9 expression in the pig oviduct during the estrous cycle and early pregnancy. J. Histochem. Cytochem. 2011, 59, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Gannon, B.J.; Warnes, G.M.; Carati, C.J.; Verco, C.J. Aquaporin-1 expression in visceral smooth muscle cells of female rat reproductive tract. J. Smooth Muscle Res. 2000, 36, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Branes, M.C.; Morales, B.; Rios, M.; Villalon, M.J. Regulation of the immunoexpression of aquaporin 9 by ovarian hormones in the rat oviductal epithelium. Am. J. Physiol. Cell Physiol. 2005, 288, 1048–1057. [Google Scholar] [CrossRef]
- Nah, W.H.; Oh, Y.S.; Hwang, J.H.; Gye, M.C. Changes in aquaporin 5 in the non-ciliated cells of mouse oviduct according to sexual maturation and oestrous cycle. Reprod. Fertil. Dev. 2015, 29, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, S.; Bosi, G.; Frattini, S.; Coizet, B.; Groppetti, D.; Pecile, A. Morphology and aquaporin immunohistochemistry of the uterine tube of Saanen goats (Capra hircus): Comparison throughout the reproductive cycle. Reprod. Domest. Anim. 2016, 51, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Zaniboni, L.; Bakst, M.R. Localization of Aquaporins in the Sperm Storage Tubules in the Turkey Oviduct. Poult. Sci. 2004, 83, 1209–1212. [Google Scholar]
- Socha, J.K.; Saito, N.; Wolak, D.; Sechman, A.; Hrabia, A. Expression of aquaporin 4 in the chicken oviduct following tamoxifen treatment. Reprod. Domest. Anim. 2018, 53, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.F.; Chen, L.Y.; Xu, K.H.; Yao, J.F.; Shi, Y.F.; Shanguan, X.J. Reduced expression of aquaporin 9 in tubal ectopic pregnancy. J. Mol. Hist. 2013, 44, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Smolinska, N.; Kiezun, M.; Dobrzyn, K.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Zaobidna, E.; Bogus-Nowakowska, K.; Wyrebek, J.; Bors, K.; et al. Expression of Chemerin and Its Receptors in the Porcine Hypothalamus and Plasma Chemerin Levels during the Oestrous Cycle and Early Pregnancy. Int. J. Mol. Sci. 2019, 20, 3887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.J.; Yu, H.M.; Koide, S.S. Regulation of water channel gene (AQP-CHIP) expression by estradiol and anordiol in rat uterus. Yao Xue Xue Bao. 1997, 32, 586–592. (In Chinese) [Google Scholar] [PubMed]
- Huang, H.F.; He, R.H.; Sun, C.C.; Zhang, Y.; Meng, Q.X.; Ma, Y.Y. Function of aquaporins in female and male reproductive systems. Hum. Reprod. Update 2006, 12, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronska, A.; Mlotkowska, P.; Eliszewski, M.; Nielsen, S.; Skowronski, M.T. Expression of aquaporin 1, 5 and 9 in the ovarian follicles of cycling and early pregnant pigs. Physiol. Res. 2015, 64, 237–245. [Google Scholar] [CrossRef]
- Skowronski, M.T. Distribution and quantitative changes in amounts of aquaporin 1, 5 and 9 in the pig uterus during the estrous cycle and early pregnancy. Reprod. Biol. Endocrinol. 2010, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1–R26. [Google Scholar] [CrossRef] [Green Version]
- Skowronski, M.T.; Mlotkowska, P.; Tanski, D.; Lepiarczyk, E.; Kempisty, B.; Jaskiewicz, L.; Pareek, C.S.; Skowronska, A. Pituitary Hormones (FSH, LH, PRL, and GH) Differentially Regulate AQP5 Expression in Porcine Ovarian Follicular Cells. Int. J. Mol. Sci. 2019, 20, 4914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronski, M.T.; Mlotkowska, P.; Tanski, D.; Lepiarczyk, E.; Oklinski, M.K.; Nielsen, S.; Skowronska, A. Pituitary Gonadotropins, Prolactin and Growth Hormone Differentially Regulate AQP1 Expression in the Porcine Ovarian Follicular Cells. Int. J. Mol. Sci. 2017, 19, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkman, A.S.; Hara-Chikuma, M.; Papadopoulos, M.C. Aquaporins-new players in cancer biology. J. Mol. Med. 2008, 86, 523–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffl, M.; Schweiger, M.; Amselgruber, W.M. Immunophenotype of porcine oviduct epithelial cells during the oestrous cycle: A double-labelling immunohistochemical study. Histochem. Cell Biol. 2004, 121, 239–244. [Google Scholar] [CrossRef]
- Wollenhaupt, K.; Kettler, A.; Brussow, K.P.; Schneider, F.; Kanitz, W.; Einspanier, R. Regulation of the expression and bioactivation of the epidermal growth factor receptor system by estradiol in pig oviduct and endometrium. Reprod. Fertil. Dev. 2001, 13, 167–176. [Google Scholar] [CrossRef]
- Wollenhaupt, K.; Welter, H.; Einspanier, R.; Manabe, N.; Brussow, K.P. Expression of epidermal growth factor receptor (EGF-R), vascular endothelial growth factor receptor (VEGF-R) and fibroblast growth factor receptor (FGF-R) systems in porcine oviduct and endometrium during the time of implantation. J. Reprod. Dev. 2004, 50, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Helguera, G.; Eghbali, M.; Sforza, D.; Minosyan, T.Y.; Toro, L.; Stefani, E. Changes in global gene expression in rat myometrium in transition from late pregnancy to parturition. Physiol. Genom. 2009, 36, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Girotti, M.; Zingg, H.H. Gene expression profiling of rat uterus at different stages of parturition. Endocrinology 2003, 144, 2254–2265. [Google Scholar] [CrossRef] [Green Version]
- Wanggren, K.; Stavreus-Evers, A.; Olsson, C.; Andersson, E.; Gemsell-Danielsson, K. Regulation of muscular contractions in the human fallopian tube through prostaglandins and progestagens. Hum. Reprod. 2008, 23, 2359–2368. [Google Scholar] [CrossRef] [Green Version]
- Antosik, P.; Kempisty, B.; Jackowska, M.; Bukowska, D.; Wozna, M.; Lianeri, M.; Brussow, K.P.; Jaskowski, J.M. Assessment of transcripts and protein contents contributing to cell cycle control and gap junction connections in morphologically variable groups of porcine cumulus-oocyte complexes. Vet. Med. 2010, 55, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Kempisty, B.; Ziolkowska, A.; Piotrowska, H.; Ciesiolka, S.; Antosik, P.; Bukowska, D.; Zawierucha, P.; Wozna, M.; Jaskowski, J.M.; Brussow, K.P.; et al. Short-term cultivation of porcine cumulus cells influences the cyclin-dependent kinase 4 (Cdk4) and connexin 43 (Cx43) protein expression -a real-time cell proliferation approach. J. Reprod. Dev. 2013, 59, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempisty, B.; Ziolkowska, A.; Piotrowska, H.; Zawierucha, P.; Antosik, P.; Bukowska, D.; Ciesiolka, S.; Jaskowski, J.M.; Brussow, K.P.; Nowicki, M.; et al. Real-time proliferation of porcine cumulus cells is related to the protein levels and cellular distribution of Cdk4 and Cx43. Theriogenology 2013, 80, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Młotkowska, P.; Tanski, D.; Eliszewski, M.; Skowronska, A.; Nielsen, S.; Skowronski, M.T. The expression profile of AQP1, AQP5 and AQP9 in granulosa and theca cells of porcine ovarian follicles during oestrous cycle and early pregnancy. J. Anim. Feed Sci. 2018, 27, 26–35. [Google Scholar] [CrossRef]
- Shimada, M.; Maeda, T.; Terada, T. Dynamic changes of connexin-43, Gap junctional protein, in outer layers of cumulus cells are regulated by PKC and PI 3-kinase during meiotic resumption in porcine oocytes. Biol. Reprod. 2001, 64, 1255–1263. [Google Scholar] [CrossRef] [Green Version]
- Ackert, C.L.; Gittens, J.E.I.; O’Brien, M.J.; Eppig, J.J.; Kidder, G.M. Intercellular Communication via Connexin43 Gap Junin the Mousections Is Required for Ovarian Folliculogenesis. Dev. Biol. 2001, 233, 258–270. [Google Scholar] [CrossRef] [Green Version]
- Gittens, J.E.I.; Kidder, G.M. Differential contributions of connexin37 and connexin43 to oogenesis revealed in chimeric reaggregated mouse ovaries. J. Cell Sci. 2005, 118, 5071–5078. [Google Scholar] [CrossRef] [Green Version]
- Vozzi, C.; Formenton, A.; Chanson, A.; Senn, A.; Sahli, R.; Shaw, P.; Nicod, P.; Germond, M.J.; Haefliger, J.A. Involvement of connexin 43 in meiotic maturation of bovine oocytem. Reproduction 2001, 122, 619–628. [Google Scholar] [CrossRef]
- McConnell, N.A.; Yunus, R.S.; Gross, S.A.; Bost, K.L.; Clemens, M.G.; Hughes, F.M., Jr. Water permeability of an ovarian antral follicle is predominantly transcellular and mediated by aquaporins. Endocrinology 2002, 8, 2905–2912. [Google Scholar] [CrossRef]
- Edashige, K.; Sakamoto, M.; Kasai, M. Expression of mRNAs of the aquaporin family in mouse oocytes and embryos. Cryobiology 2000, 40, 171–175. [Google Scholar] [CrossRef]
- Meng, Q.X.; Gao, H.J.; Xu, C.M.; Dong, M.Y.; Sheng, X.; Sheng, J.Z.; Huang, H.F. Reduced expression and function of aquaporin-3 in mouse metaphase-II oocytes induced by controlled ovarian hyperstimulation were associated with subsequent low fertilization rate. Cell Physiol. Biochem. 2005, 21, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Kolle, S.; Dubielzig, S.; Reese, S.; Wehrend, A.; Konig, P.; Kummer, W. Ciliary transport, gamete interaction, and effects of the early embryo in the oviduct: Ex vivo analyses using a new digital videomicroscopic system in the cow. Biol. Reprod. 2009, 81, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillo, V.; Sanchez-Calabuig, M.J.; Lopera-Vasquez, R.; Hamdi, M.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. Oviductal response to gametes and early embryos in mammals. Reproduction 2016, 152, R127–R141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budna-Tukan, J.; Swiatly-Blaszkiewicz, A.; Celichowski, P.; Kaluzna, S.; Konwerska, A.; Sujka-Kordowska, P.; Jankowski, M.; Kulus, M.; Jeseta, M.; Piotrowska-Kempisty, H.; et al. “Biological Adhesion” is a Significantly Regulated Molecular Process during Long-Term Primary In Vitro Culture of Oviductal Epithelial Cells (Oecs): A Transcriptomic and Proteomic Study. Int. J. Mol. Sci. 2019, 20, 3387. [Google Scholar] [CrossRef] [Green Version]
- Akins, E.L.; Morrissette, M.C. Gross ovarian changes during estrous cycle of swine. Am. J. Vet. Res. 1968, 29, 1953–1957. [Google Scholar]
- Dobrzyn, K.; Smolinska, N.; Kiezun, M.; Szeszko, K.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Kaminski, T. The in vitro effect of progesterone on the orexin system in porcine uterine tissues during early pregnancy. Acta. Vet. Scand. 2018, 60, 76. [Google Scholar] [CrossRef]
- Lord, E.; Ledoux, S.; Murphy, B.D.; Beaudry, D.; Palin, M.F. Expression of adiponectin and its receptors in swine. J. Anim. Sci. 2005, 83, 565–578. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Aquaporin Isoforms | Species | Vessels | Muscles | Luminal Epithelium | Articles |
|---|---|---|---|---|---|
| AQP1 | Rat | + | + | Gannon BJ. et al. [10] | |
| Pig | + | Skowronski MT et al. [8] | |||
| Goat | + | + | + | Arrighi S et al. [13] | |
| AQP2 | Turkey | + | Zaniboni L. and Bakst MR. [14] | ||
| AQP3 | Turkey | + | Zaniboni L. and Bakst MR. [14] | ||
| AQP4 | Goat | + | Arrighi S et al. [13] | ||
| Chicken | + | + | + | Socha JK. et al. [15] | |
| AQP5 | Rat | + | Branes MC et al. [11] | ||
| Pig | + | + | Skowronski MT et al. [8] | ||
| Mouse | + | + | Nah WH. et al. [12] | ||
| Goat | + | Arrighi S. et al. [13] | |||
| AQP6 | - | - | - | - | - |
| AQP7 | - | - | - | - | - |
| AQP8 | Rat | + | Branes MC. et al. [11] | ||
| AQP9 | Turkey | + | Zaniboni L. and Bakst M.R. [14] | ||
| Rat | + | Branes MC. et al. [11] | |||
| Pig | + | Skowronski MT. et al. [8] | |||
| Human | + | Ji YF et al. [16] | |||
| AQP10 | - | - | - | - | - |
| AQP11 | - | - | - | - | - |
| AQP12 | - | - | - | - | - |
| Name of the Gene | Primer Sequence Forward/Reverse | Amplicon Length, bp | Accession Number |
|---|---|---|---|
| Aquaporin 1 (Aqp1) | 5′-CAGCGAGTTCAAGAAGAAG-3′ 5′-GCGACACCTTCACGTTATC-3′ | 161 | NM_214454.1 |
| Aquaporin 5 (Aqp5) | 5′-CTATGAGTCCGAGGAGGATT-3′ 5′-GCTTCGCTGTCATCTGTT-3′ | 147 | NM_001110424.1 |
| Aquaporin 9 (Aqp9) | 5′-TCTGGTGGATTCCTGTAGTG-3′ 5′-GGTTTGTCCTCCGATTGTTC-3′ | 130 | NM_001112684.1 |
| Cyclophilin (PPIA) | 5′-GCACTGGTGGCAAGTCCAT-3′ 5′-AGGACCCGTATGCTTCAGGA-3′ | 71 | XM_021078519.1 |
| 18S ribosomal RNA (18S rRNA) | 5′-GGCTACCACATCCAAGGAAG-3′ 5′-TCCAATGGATCCTCGCGGAA-3′ | 149 | AK393333.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanski, D.; Skowronska, A.; Eliszewski, M.; Gromadzinski, L.; Kempisty, B.; Skowronski, M.T. Changes in Aquaporin 1, 5 and 9 Gene Expression in the Porcine Oviduct According to Estrous Cycle and Early Pregnancy. Int. J. Mol. Sci. 2020, 21, 2777. https://doi.org/10.3390/ijms21082777
Tanski D, Skowronska A, Eliszewski M, Gromadzinski L, Kempisty B, Skowronski MT. Changes in Aquaporin 1, 5 and 9 Gene Expression in the Porcine Oviduct According to Estrous Cycle and Early Pregnancy. International Journal of Molecular Sciences. 2020; 21(8):2777. https://doi.org/10.3390/ijms21082777
Chicago/Turabian StyleTanski, Damian, Agnieszka Skowronska, Maciej Eliszewski, Leszek Gromadzinski, Bartosz Kempisty, and Mariusz T. Skowronski. 2020. "Changes in Aquaporin 1, 5 and 9 Gene Expression in the Porcine Oviduct According to Estrous Cycle and Early Pregnancy" International Journal of Molecular Sciences 21, no. 8: 2777. https://doi.org/10.3390/ijms21082777