The Surprising Effect of Phenformin on Cutaneous Darkening and Characterization of Its Underlying Mechanism by a Forward Chemical Genetics Approach

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phenformin Substantially Darkens Human Skin in a Dose-Dependent Manner
2.2. Phenformin Suppresses Melanosome Degradation in Epidermal Cells
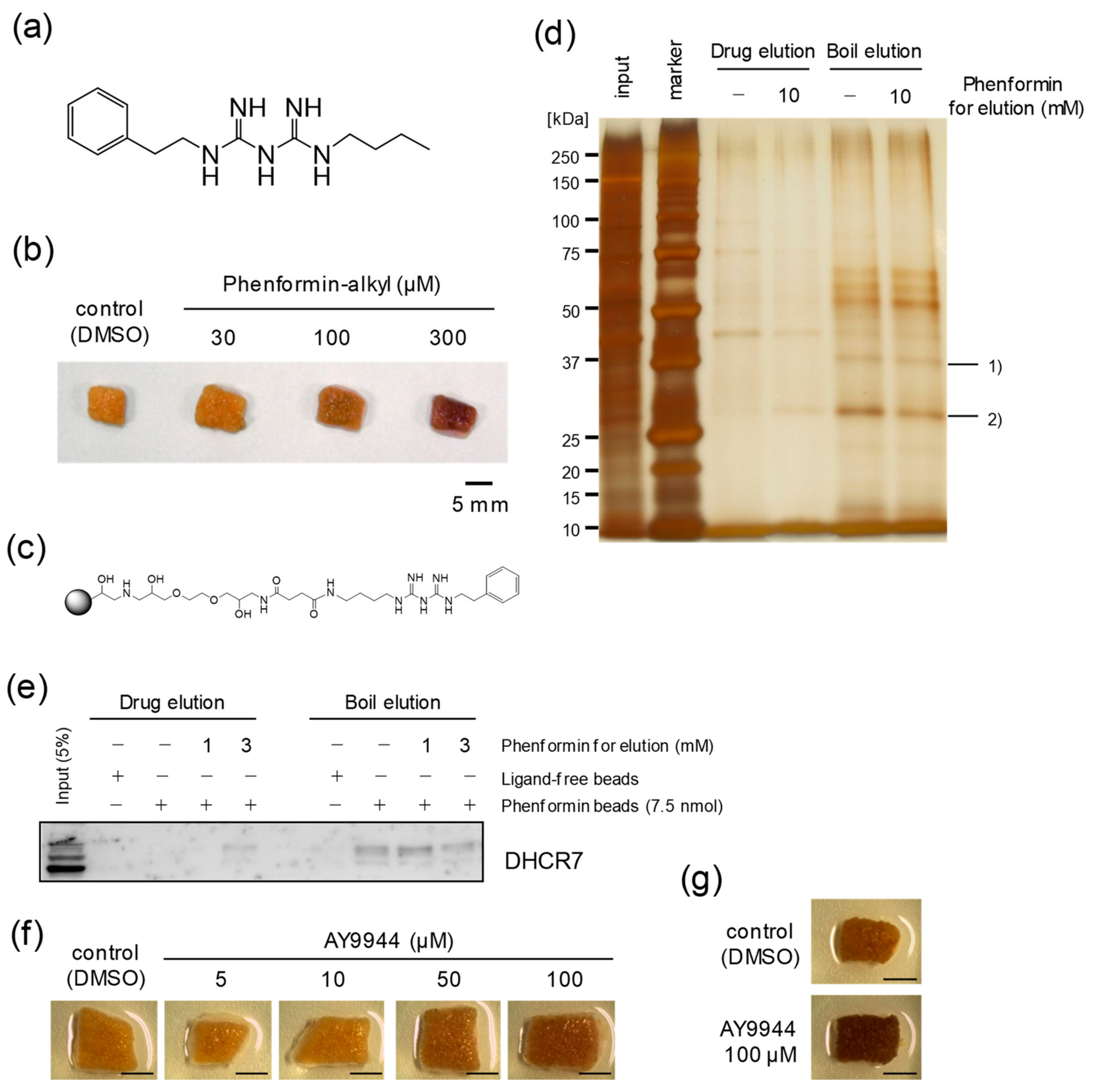
2.3. Identification of DHCR7 as a Phenformin-Binding Protein in Epidermal Cells
2.4. Phenformin as well as AY9944 Suppresses Autophagic Activity, Leading to the Accumulation of Melanosomes in Epidermal Cells
2.5. Induction of Cutaneous Darkening by Berberine Emphasizes the Involvement of Cholesterol Biosynthesis in Skin Color Development
2.6. Berberine Affects Not only Melanosome Degradation in Epidermal Cells but also Melanin Synthesis in Melanocytes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Human Skin
4.3. Cell Culture
4.4. Measurement of DOPA Oxidase Activity
4.5. Measurement of Melanin Content in NHEMs
4.6. Melanosome Incorporation into NHEKs
4.7. Western Blot Analysis
4.8. Preparation of Phenformin-Immobilized FG-Beads
4.9. Identification of Phenformin-Binding Proteins (Chemical Pull-down Assay)
4.10. Human Skin Culture
4.11. Fontana-Masson Staining
4.12. Immunofluorescence Microscopic Analysis
4.13. Measurement of Intracellular Cholesterol Contents
4.14. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | African-American |
| AMPK | AMP-activated protein kinase |
| BPE | Bovine pituitary extract |
| DAPI | 4’6-diamidino-2-phenylindole |
| DDW | Double distilled water |
| DHCR7 | 7-dehydrocholesterol reductase |
| DMF | N, N’-dimethylformamide |
| DMSO | Dimethyl sulfoxide |
| DOPA | Dihydroxyphenylalanine |
| EGF | Epidermal growth factor |
| FBS | Fetal bovine serum |
| HMG-CoA | 3-hydroxy-3-methylglutaryl coenzyme A |
| NHEK | Normal human epidermal keratinocyte |
| NHEM | Normal human epidermal melanocyte |
| OTC | Over-the-counter |
| PBS | Phosphate-buffered saline |
| PMSF | Phenylmethylsulfonyl fluoride |
| SAR | Structure-activity relationship |
| SR-BI | Scavenger receptor class B type 1 |
| TRP1 | Tyrosinase-related protein 1 |
| TYR | Tyrosinase |
References
- Sturm, R.A. Molecular genetics of human pigmentation diversity. Hum. Mol. Genet. 2009, 18, R9–R17. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, W.C.; Holstein, T.J. General biology of mammalian pigmentation. In The Pigmentary System; Nordlund, J.J., Boissy, R.E., Hearing, V.J., King, R.A., Oetting, W.S., Ortonne, J.-P., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 63–90. [Google Scholar]
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef] [PubMed]
- Halder, R.M.; Nootheti, P.K. Ethnic skin disorders overview. J. Am. Acad. Dermatol. 2003, 48, S143–S148. [Google Scholar] [CrossRef] [PubMed]
- Swinney, D.C.; Anthony, J. How were new medicines discovered? Nat. Rev. Drug Discov. 2011, 10, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Murase, D.; Hachiya, A.; Takano, K.; Hicks, R.; Visscher, M.O.; Kitahara, T.; Hase, T.; Takema, Y.; Yoshimori, T. Autophagy Has a Significant Role in Determining Skin Color by Regulating Melanosome Degradation in Keratinocytes. J. Investig. Dermatol. 2013, 133, 2416–2424. [Google Scholar] [CrossRef] [Green Version]
- Murase, D.; Hachiya, A.; Fullenkamp, R.; Beck, A.; Moriwaki, S.; Hase, T.; Takema, Y.; Manga, P. Variation in Hsp70-1A Expression Contributes to Skin Color Diversity. J. Investig. Dermatol. 2016, 136, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Alaimo, P.J.; Shogren-Knaak, M.A.; Shokat, K.M. Chemical genetic approaches for the elucidation of signaling pathways. Curr. Opin. Chem. Biol. 2001, 5, 360–367. [Google Scholar] [CrossRef]
- Zheng, X.F.; Chan, T.F. Chemical genomics in the global study of protein functions. Drug Discov. Today. 2002, 7, 197–205. [Google Scholar] [CrossRef]
- Hart, C.P. Finding the target after screening the phenotype. Drug Discov. Today. 2005, 10, 513–519. [Google Scholar] [CrossRef]
- Tashiro, E.; Imoto, M. Target identification of bioactive compounds. Bioorg. Med. Chem. 2012, 20, 1910–1921. [Google Scholar] [CrossRef]
- Kawatani, M.; Osada, H. Affinity-based identification for bioactive small molecules. Med Chem. Commun. 2014, 5, 277–287. [Google Scholar] [CrossRef]
- Lehraiki, A.; Abbe, P.; Cerezo, M.; Rouaud, F.; Regazzetti, C.; Chignon-Sicard, B.; Passeron, T.; Bertolotto, C.; Ballotti, R.; Rocchi, S. Inhibition of Melanogenesis by the Antidiabetic Metformin. J. Investig. Dermatol. 2014, 134, 2589–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterfield, W.J.; Whichelow, M.J. The hypoglycemic action of phenformin. Effect of phenformin on glucose metabolism in peripheral tissues. Diabetes 1962, 11, 281–286. [Google Scholar] [PubMed]
- Searle, G.L.; Gulli, R. The mechanism of the acute hypoglycemic action of phenformin (DBI). Metabolism 1980, 29, 630–635. [Google Scholar] [CrossRef]
- Zhang, L.; He, H.; Balschi, J.A. Metformin and phenformin activate AMP-activated protein kinase in the heart by increasing cytosolic AMP concentration. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H457–H466. [Google Scholar] [CrossRef] [Green Version]
- Winder, A.J.; Harris, H. New assays for the tyrosinase hydroxylase and dopa oxidase activities of tyrosinase. Eur. J. Biochem. 1991, 198, 317–326. [Google Scholar] [CrossRef]
- Del Marmol, V.; Beermann, F. Tyrosinase and related proteins in mammalian pigmentation. FEBS Lett. 1996, 381, 165–168. [Google Scholar] [CrossRef]
- Sakamoto, S.; Kabe, Y.; Hatakeyama, M.; Yamaguchi, Y.; Handa, H. Development and application of high-performance affinity beads: toward chemical biology and drug discovery. Chem. Rec. 2009, 9, 66–85. [Google Scholar] [CrossRef]
- Tint, G.S.; Batta, A.K.; Frieden, R.; Chen, T.S.; Irons, M.; Elias, E.R.; Salen, G. Defective Cholesterol Biosynthesis Associated with the Smith-Lemli-Opitz Syndrome. New Engl. J. Med. 1994, 330, 107–113. [Google Scholar] [CrossRef]
- Clémençon, B.; Babot, M.; Trézégue, V. The mitochondrial ADP/ATP carrier (SLC25 family): pathological implications of its dysfunction. Mol. Asp. Med. 2013, 34, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, F.; Monné, M. Discoveries, metabolic roles and diseases of mitochondrial carriers: A review. Biochim. Biophys Acta. 2016, 1863, 2362–2378. [Google Scholar] [CrossRef] [PubMed]
- Rosand, J.; Friedberg, J.W.; Yang, J.M. Fatal phenformin-associated lactic acidosis. Ann. Intern. Med. 1997, 127, 170. [Google Scholar] [CrossRef] [PubMed]
- Kraml, M.; Bagli, J.F.; Dvornik, D. Inhibition of the conversion of 7-dehydrocholesterol to cholesterol by AY-9944. Biochem. Biophys Res. Commun. 1964, 15, 455–457. [Google Scholar] [CrossRef]
- Dempsey, M.E. Inhibition of lipid biosynthesis. Ann. N. Y. Acad. Sci. 1968, 148, 631–646. [Google Scholar] [CrossRef]
- Lucero, H.A.; Robbins, P.W. Lipid rafts-protein association and the regulation of protein activity. Arch Biochem. Biophys. 2004, 426, 208–224. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T. How to interpret LC3 immunoblotting. Autophagy. 2007, 3, 542–545. [Google Scholar] [CrossRef]
- Wu, N.; Sarna, L.K.; Siow, Y.L.; Karmin, O. Regulation of hepatic cholesterol biosynthesis by berberine during hyperhomocysteinemia. Am. J. Physiol. Regul. Integr. Comp Physiol. 2011, 300, R635–R643. [Google Scholar] [CrossRef]
- Pollak, M. Metformin and other biguanides in oncology: Advancing the research agenda. J Cancer Prev. Res. 2010, 3, 1060–1065. [Google Scholar] [CrossRef] [Green Version]
- Rena, G.; Pearson, E.R.; Sakamoto, K. Molecular mechanism of action of metformin: old or new insights? Diabetologia 2013, 56, 1898–1906. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Issandou, M.; Guillard, R.; Boullay, A.B.; Linhart, V.; Lopez-Perez, E. Up-regulation of low-density lipoprotein receptor in human hepatocytes is induced by sequestration of free cholesterol in the endosomal/lysosomal compartment. Biochem. Pharm. 2004, 67, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Tsuruoka, H.; Khovidhunkit, W.; Brown, B.E.; Fluhr, J.W.; Elias, P.M.; Feingold, K.R. Scavenger receptor class B type I is expressed in cultured keratinocytes and epidermis. Regulation in response to changes in cholesterol homeostasis and barrier requirements. J. Biol. Chem. 2002, 277, 2916–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinslechner, K.; Schütz, B.; Pistek, M.; Rapolter, P.; Weitzenböck, H.P.; Hundsberger, H.; Mikulits, W.; Grillari, R.; Röhrl, C.; Hengstschläger, M.; et al. Loss of SR-BI Down-Regulates MITF and Suppresses Extracellular Vesicle Release in Human Melanoma. Int. J. Mol. Sci. 2019, 20, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, A.; Kuroda, M.; Tanzawa, K. Competitive inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase by ML-236A and ML-236B fungal metabolites, having hypocholesterolemic activity. FEBS Lett. 1976, 72, 323–326. [Google Scholar] [CrossRef] [Green Version]
- Galus, R.; Niderla, J.; Sladowski, D.; Sajjad, E.; Włodarski, K.; Jóźwiak, J. Fluvastatin increases tyrosinase synthesis induced by alpha-melanocyte-stimulating hormone in B16F10 melanoma cells. Pharm. Rep. 2010, 62, 164–169. [Google Scholar] [CrossRef]
- Galus, R.; Włodarski, K.; Malejczyk, J.; Jóźwiak, J. Fluvastatin influences hair color in C57BL/6 mice. Int. J. Mol. Sci. 2013, 14, 14333–14345. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, J.; Jung, E.; Cho, J.Y.; Park, D. Artemisinic acid inhibits melanogenesis through downregulation of C/EBP α-dependent expression of HMG-CoA reductase gene. Food Chem. Toxicol. 2013, 51, 225–230. [Google Scholar] [CrossRef]
- Lee, H.J.; Jo, S.Y.; Hwang, J.S.; Chan, S.E. Mevastatin suppresses melanogenesis by lowering the levels of cyclic adenosine monophosphate and cholesterol. Exp Derm. 2016, 25, 820–822. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef]
- Abu, T.M.; Pramod, K.; Ansari, S.H.; Ali, J. Current remedies for vitiligo. Autoimmun. Rev. 2010, 9, 516–520. [Google Scholar]
- Ali, S.A.; Naaz, I.; Choudhary, R.K. Berberine-induced pigment dispersion in Bufo melanostictus melanophores by stimulation of beta-2 adrenergic receptors. J. Recept. Signal Transduct. Res. 2014, 34, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Yoshida, M.; Ito, M.; Akiyama, K.; Kim, J.H.; Yoon, T.J.; Lee, J.H.; Matsui, M.S.; Ichihashi, M.; et al. Keratinocytes in culture accumulate phagocytosed melanosomes in the perinuclear area. Pigment Cell Melanoma Res. 2010, 23, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, A.; Sriwiriyanont, P.; Kaiho, E.; Kitahara, T.; Takema, Y.; Tsuboi, R. An in vivo mouse model of human skin substitute containing spontaneously sorted melanocytes demonstrates physiological changes after UVB irradiation. J. Invest Derm. 2005, 125, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takano, K.; Hachiya, A.; Murase, D.; Kawasaki, A.; Uda, H.; Kasamatsu, S.; Sugai, Y.; Takahashi, Y.; Hase, T.; Ohuchi, A.; et al. The Surprising Effect of Phenformin on Cutaneous Darkening and Characterization of Its Underlying Mechanism by a Forward Chemical Genetics Approach. Int. J. Mol. Sci. 2020, 21, 1451. https://doi.org/10.3390/ijms21041451
Takano K, Hachiya A, Murase D, Kawasaki A, Uda H, Kasamatsu S, Sugai Y, Takahashi Y, Hase T, Ohuchi A, et al. The Surprising Effect of Phenformin on Cutaneous Darkening and Characterization of Its Underlying Mechanism by a Forward Chemical Genetics Approach. International Journal of Molecular Sciences. 2020; 21(4):1451. https://doi.org/10.3390/ijms21041451
Chicago/Turabian StyleTakano, Kei, Akira Hachiya, Daiki Murase, Akiko Kawasaki, Hirokazu Uda, Shinya Kasamatsu, Yoshiya Sugai, Yoshito Takahashi, Tadashi Hase, Atsushi Ohuchi, and et al. 2020. "The Surprising Effect of Phenformin on Cutaneous Darkening and Characterization of Its Underlying Mechanism by a Forward Chemical Genetics Approach" International Journal of Molecular Sciences 21, no. 4: 1451. https://doi.org/10.3390/ijms21041451