A Comparison of the Variable J and Carbon-Isotopic Composition of Sugars Methods to Assess Mesophyll Conductance from the Leaf to the Canopy Scale in Drought-Stressed Cherry

 , , ,
, , ,  and
and
Abstract
:1. Introduction
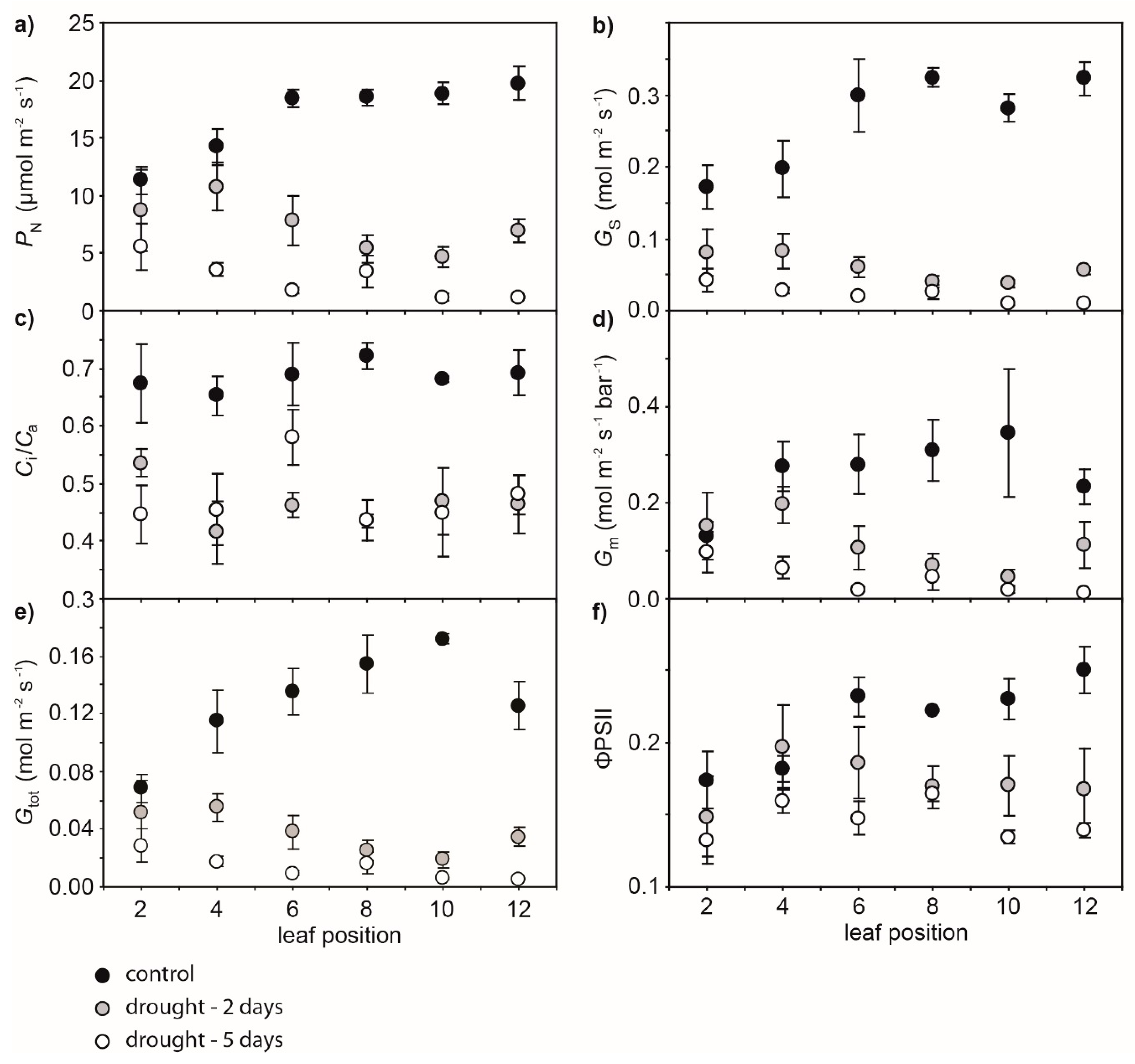
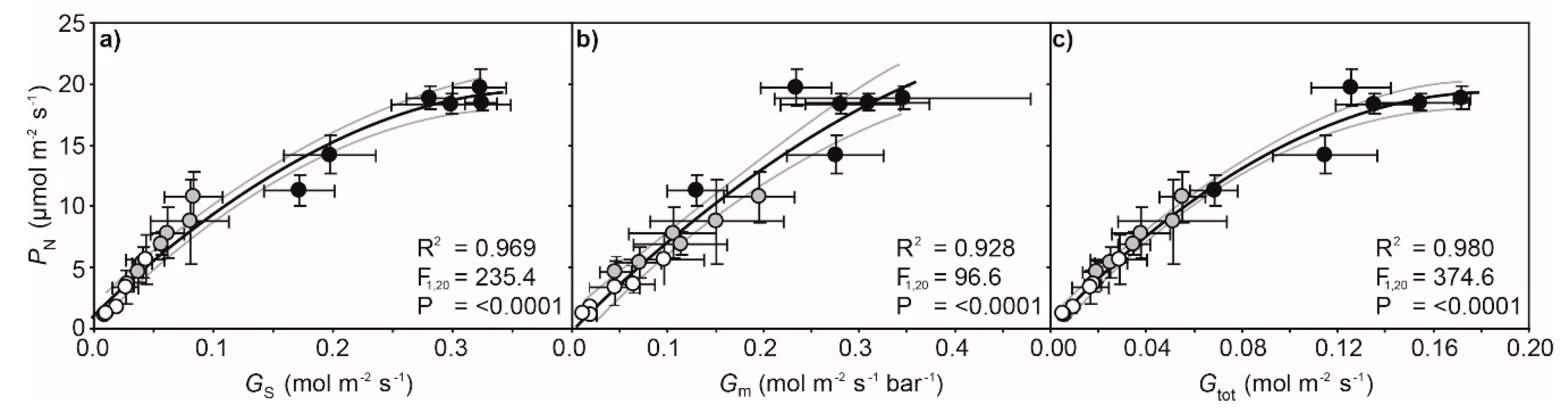
2. Results
3. Discussion
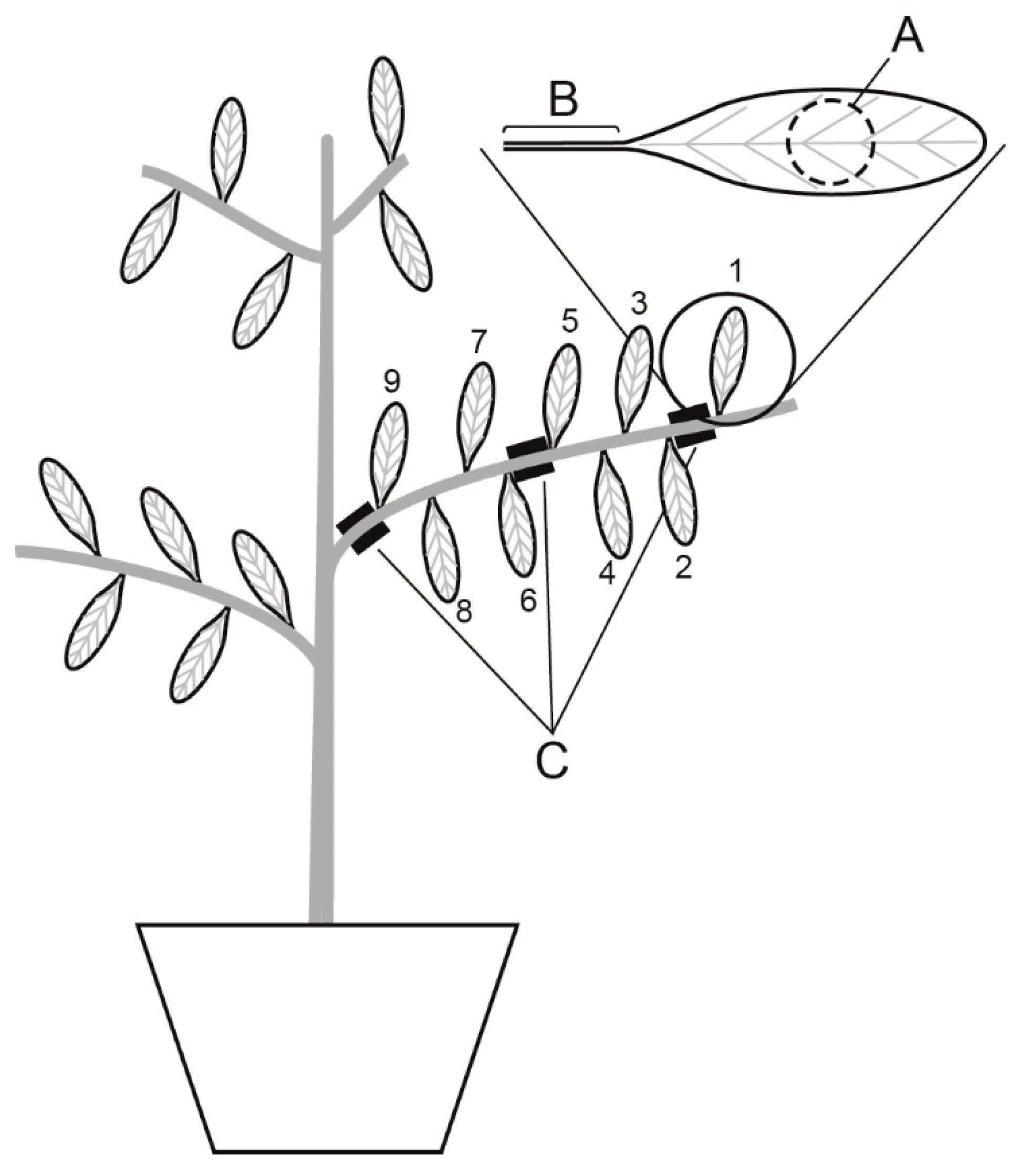
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Gas Exchange and Fluorescence Measurements
4.3. Leaf Sampling, Measurement of Leaf Water Potential, and Sap Collection
4.4. Analysis of Carbon Isotopic Composition and Calculation of Gm
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Harley, P.C.; Dimarco, G.; Sharkey, T.D. Estimation of mesophyll conductance to CO2 flux by three different methods. Plant Physiol. 1992, 98, 1437–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centritto, M.; Loreto, F.; Chartzoulakis, K. The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings. Plant Cell Environ. 2003, 26, 585–594. [Google Scholar] [CrossRef]
- Heath, O.V.S. Studies in stomatal behaviour. V. The role of carbon dioxide in the light response of stomata. J. Exp. Bot. 1950, 1, 29–62. [Google Scholar] [CrossRef]
- Heath, J. Stomata of trees growing in CO2-enriched air show reduced sensitivity to vapour pressure deficit and drought. Plant Cell Environ. 1998, 21, 1077–1088. [Google Scholar] [CrossRef]
- Haworth, M.; Cosentino, S.L.; Marino, G.; Brunetti, C.; Riggi, E.; Avola, G.; Loreto, F.; Centritto, M. Increased free abscisic acid during drought enhances stomatal sensitivity and modifies stomatal behaviour in fast growing giant reed (Arundo donax L.). Environ. Exp. Bot. 2018, 147, 116–124. [Google Scholar] [CrossRef]
- Aganchich, B.; Wahbi, S.; Loreto, F.; Centritto, M. Partial root zone drying: Regulation of photosynthetic limitations and antioxidant enzymatic activities in young olive (Olea europaea) saplings. Tree Physiol. 2009, 29, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Lauteri, M.; Haworth, M.; Serraj, R.; Monteverdi, M.C.; Centritto, M. Photosynthetic diffusional constraints affect yield in drought stressed rice cultivars during flowering. PLoS ONE 2014, 9, e109054. [Google Scholar] [CrossRef]
- Loreto, F.; Dimarco, G.; Tricoli, D.; Sharkey, T.D. Measurements of mesophyll conductance, photosynthetic electron Transp. and alternative electron sinks of field grown wheat leaves. Photosynth. Res. 1994, 41, 397–403. [Google Scholar] [CrossRef]
- Adachi, S.; Nakae, T.; Uchida, M.; Soda, K.; Takai, T.; Oi, T.; Yamamoto, T.; Ookawa, T.; Miyake, H.; Yano, M.; et al. The mesophyll anatomy enhancing CO2 diffusion is a key trait for improving rice photosynthesis. J. Exp. Bot. 2013, 64, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Kaldenhoff, R. Mechanisms underlying CO2 diffusion in leaves. Curr. Opin. Plant Biol. 2012, 15, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Niinemets, Ü.; Gallé, A.; Barbour, M.; Centritto, M.; Diaz-Espejo, A.; Douthe, C.; Galmés, J.; Ribas-Carbo, M.; Rodriguez, P.; et al. Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Moreno, M.J.; Gago, J.; Díaz-Vivancos, P.; Bernal, A.; Miedes, E.; Bresta, P.; Liakopoulos, G.; Fernie, A.R.; Hernández, J.A.; Flexas, J. The apoplastic antioxidant system and altered cell wall dynamics influence mesophyll conductance and the rate of photosynthesis. Plant J. 2019, 99, 1031–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flexas, J.; Ribas-Carbó, M.; Diaz-Espejo, A.; Galmēs, J.; Medrano, H. Mesophyll conductance to CO2: Current knowledge and future prospects. Plant Cell Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef]
- Lloyd, J.; Syvertsen, J.P.; Kriedemann, P.E.; Farquhar, G.D. Low conductances for CO2 diffusion from stomata to the sites of carboxylation in leaves of woody species. Plant Cell Environ. 1992, 15, 873–899. [Google Scholar] [CrossRef]
- Ethier, G.J.; Livingston, N.J. On the need to incorporate sensitivity to CO2 transfer conductance into the Farquhar–von Caemmerer–Berry leaf photosynthesis model. Plant Cell Environ. 2004, 27, 137–153. [Google Scholar] [CrossRef]
- Harley, P.C.; Loreto, F.; Dimarco, G.; Sharkey, T.D. Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Brugnoli, E.; Hubick, K.T.; Voncaemmerer, S.; Wong, S.C.; Farquhar, G.D. Correlation between the carbon isotope discrimination in leaf starch and sugars of C3 plants and the ratio of intercellular and atmospheric partial pressures of carbon dioxide. Plant Physiol. 1988, 88, 1418–1424. [Google Scholar] [CrossRef] [Green Version]
- Centritto, M.; Lauteri, M.; Monteverdi, M.C.; Serraj, R. Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. J. Exp. Bot. 2009, 60, 2325–2339. [Google Scholar] [CrossRef]
- Pons, T.L.; Flexas, J.; von Caemmerer, S.; Evans, J.R.; Genty, B.; Ribas-Carbo, M.; Brugnoli, E. Estimating mesophyll conductance to CO2: Methodology, potential errors, and recommendations. J. Exp. Bot. 2009, 60, 2217–2234. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.R. In Stand aside stomata, another actor deserves centre stage: The forgotten role of the internal conductance to CO2 transfer. In Proceedings of the 14th International Congress of Photosynthesis, Glasgow, Scotland, 22–27 July 2007; Oxford University Press: Glasgow, Scotland, 2007; pp. 1475–1487. [Google Scholar]
- Fini, A.; Loreto, F.; Tattini, M.; Giordano, C.; Ferrini, F.; Brunetti, C.; Centritto, M. Mesophyll conductance plays a central role in leaf functioning of Oleaceae species exposed to contrasting sunlight irradiance. Physiol. Plant 2016, 157, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Tomás, M.; Flexas, J.; Copolovici, L.; Galmés, J.; Hallik, L.; Medrano, H.; Ribas-Carbó, M.; Tosens, T.; Vislap, V.; Niinemets, Ü. Importance of leaf anatomy in determining mesophyll diffusion conductance to CO2 across species: Quantitative limitations and scaling up by models. J. Exp. Bot. 2013, 64, 2269–2281. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S.; Tognetti, R.; Minnocci, A.; Borghi, M.; Sebastiani, L. Variation in mesophyll anatomy and photosynthetic capacity during leaf development in a deciduous mesophyte fruit tree (Prunus persica) and an evergreen sclerophyllous Mediterranean shrub (Olea europaea). Trees-Struct. Funct. 2008, 22, 559–571. [Google Scholar] [CrossRef]
- Hanba, Y.T.; Shibasaka, M.; Hayashi, Y.; Hayakawa, T.; Kasamo, K.; Terashima, I.; Katsuhara, M. Overexpression of the barley aquaporin HvPIP2; 1 increases internal CO2 conductance and CO2 assimilation in the leaves of transgenic rice plants. Plant Cell Physiol. 2004, 45, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Killi, D.; Haworth, M. Diffusive and metabolic constraints to photosynthesis in quinoa during drought and salt stress. Plants 2017, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, G.; Haworth, M.; Wahbi, S.; Mahmood, T.; Zuomin, S.; Centritto, M. Abscisic acid induces rapid reductions in mesophyll conductance to carbon dioxide. PLoS ONE 2016, 11, e0148554. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Diaz-Espejo, A.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell Environ. 2007, 30, 1284–1298. [Google Scholar] [CrossRef]
- Dbara, S.; Haworth, M.; Emiliani, G.; Mimoun, M.B.; Gómez-Cadenas, A.; Centritto, M. Partial root-zone drying of olive (Olea europaea var.‘Chetoui’) induces reduced yield under field conditions. PLoS ONE 2016, 11, e0157089. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Pallardy, S.G.; Tu, K.; Law, B.E.; Wullschleger, S.D. Reliable estimation of biochemical parameters from C3 leaf photosynthesis–intercellular carbon dioxide response curves. Plant Cell Environ. 2010, 33, 1852–1874. [Google Scholar] [CrossRef]
- Tazoe, Y.; von Caemmerer, S.; Badger, M.R.; Evans, J.R. Light and CO2 do not affect the mesophyll conductance to CO2 diffusion in wheat leaves. J. Exp. Bot. 2009, 60, 2291–2301. [Google Scholar] [CrossRef] [Green Version]
- Douthe, C.; Dreyer, E.; Brendel, O.; Warren, C.R. Is mesophyll conductance to CO2 in leaves of three Eucalyptus species sensitive to short-term changes of irradiance under ambient as well as low O2? Funct. Plant Biol. 2012, 39, 435–448. [Google Scholar] [CrossRef]
- Tholen, D.; Ethier, G.; Genty, B.; Pepin, S.; Zhu, X.-G. Variable mesophyll conductance revisited: Theoretical background and experimental implications. Plant Cell Environ. 2012, 35, 2087–2103. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.E.; Pou, A.; Zwieniecki, M.A.; Holbrook, N.M. On measuring the response of mesophyll conductance to carbon dioxide with the variable J method. J. Exp. Bot. 2012, 63, 413–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunce, J.A. Use of the response of photosynthesis to oxygen to estimate mesophyll conductance to carbon dioxide in water-stressed soybean leaves. Plant Cell Environ. 2009, 32, 875–881. [Google Scholar] [CrossRef]
- Evans, J.R.; Sharkey, T.D.; Berry, J.A.; Farquhar, G.D. Carbon isotope discrimination measured concurrently with gas exchange to investigate CO2 diffusion in leaves of higher plants. Aust. J. Plant Physiol. 1986, 13, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Rodeghiero, M.; Niinemets, Ü.; Cescatti, A. Major diffusion leaks of clamp-on leaf cuvettes still unaccounted: How erroneous are the estimates of Farquhar et al. model parameters? Plant Cell Environ. 2007, 30, 1006–1022. [Google Scholar] [CrossRef]
- Haworth, M.; Marino, G.; Riggi, E.; Avola, G.; Brunetti, C.; Scordia, D.; Testa, G.; Gomes, M.T.G.; Loreto, F.; Cosentino, S.L.; et al. The effect of summer drought on the yield of Arundo donax is reduced by the retention of photosynthetic capacity and leaf growth later in the growing season. Ann. Bot. 2019, 124, 567–579. [Google Scholar] [CrossRef]
- Di Marco, G.; Manes, F.; Tricoli, D.; Vitale, E. Fluorescence parameters measured concurrently with net photosynthesis to investigate chloroplastic CO2 concentration in leaves of Quercus ilex L. J. Plant Physiol. 1990, 136, 538–543. [Google Scholar] [CrossRef]
- Flexas, J.; Díaz-Espejo, A.; Berry, J.; Cifre, J.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbó, M. Analysis of leakage in IRGA’s leaf chambers of open gas exchange systems: Quantification and its effects in photosynthesis parameterization. J. Exp. Bot. 2007, 58, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Loriaux, S.; Avenson, T.; Welles, J.; McDermitt, D.; Eckles, R.; Riensche, B.; Genty, B. Closing in on maximum yield of chlorophyll fluorescence using a single multiphase flash of sub-saturating intensity. Plant Cell Environ. 2013, 36, 1755–1770. [Google Scholar] [CrossRef]
- Farquhar, G.; Hubick, K.; Condon, A.; Richards, R. Carbon Isotope Fractionation and Plant Water-Use Efficiency. In Stable Isotopes in Ecological Research; Springer: New York, NY, USA, 1989; pp. 21–40. [Google Scholar]
- Vrábl, D.; Vašková, M.; Hronkova, M.; Flexas, J.; Šantrůček, J. Mesophyll conductance to CO2 transport estimated by two independent methods: Effect of variable CO2 concentration and abscisic acid. J. Exp. Bot. 2009, 60, 2315–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tazoe, Y.; Von Caemmerer, S.; Estavillo, G.M.; Evans, J.R. Using tunable diode laser spectroscopy to measure carbon isotope discrimination and mesophyll conductance to CO2 diffusion dynamically at different CO2 concentrations. Plant Cell Environ. 2011, 34, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Lauteri, M.; Scartazza, A.; Guido, M.C.; Brugnoli, E. Genetic variation in photosynthetic capacity, carbon isotope discrimination and mesophyll conductance in provenances of Castanea sativa adapted to different environments. Funct. Ecol. 1997, 11, 675–683. [Google Scholar] [CrossRef]
- Ubierna, N.; Marshall, J.D. Estimation of canopy average mesophyll conductance using delta C-13 of phloem contents. Plant Cell Environ. 2011, 34, 1521–1535. [Google Scholar] [CrossRef]
- Gentsch, L.; Hammerle, A.; Sturm, P.; Ogée, J.; Wingate, L.; Siegwolf, R.; Plüss, P.; Baur, T.; Buchmann, N.; Knohl, A. Carbon isotope discrimination during branch photosynthesis of Fagus sylvatica: A Bayesian modelling approach. Plant Cell Environ. 2014, 37, 1516–1535. [Google Scholar] [CrossRef] [Green Version]
- Scartazza, A.; Moscatello, S.; Matteucci, G.; Battistelli, A.; Brugnoli, E. Seasonal and inter-annual dynamics of growth, non-structural carbohydrates and C stable isotopes in a Mediterranean beech forest. Tree Physiol. 2013, 33, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Scartazza, A.; Moscatello, S.; Matteucci, G.; Battistelli, A.; Brugnoli, E. Combining stable isotope and carbohydrate analyses in phloem sap and fine roots to study seasonal changes of source–sink relationships in a Mediterranean beech forest. Tree Physiol. 2015, 35, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, R. The sink-source transition in leaves. Annu. Rev. Plant Biol. 1989, 40, 119–138. [Google Scholar] [CrossRef]
- Badeck, F.-W.; Tcherkez, G.; Nogués, S.; Piel, C.; Ghashghaie, J. Post-photosynthetic fractionation of stable carbon isotopes between plant organs—A widespread phenomenon. Rapid Commun. Mass Spectrom. 2005, 19, 1381–1391. [Google Scholar] [CrossRef]
- Haworth, M.; Catola, S.; Marino, G.; Brunetti, C.; Michelozzi, M.; Riggi, E.; Avola, G.; Cosentino, S.L.; Loreto, F.; Centritto, M. Moderate drought stress induces increased foliar dimethylsulphoniopropionate (DMSP) concentration and isoprene emission in two contrasting ecotypes of Arundo donax. Front. Plant Sci. 2017, 8, 1016. [Google Scholar] [CrossRef]
- Scartazza, A.; Lauteri, M.; Guido, M.C.; Brugnoli, E. Carbon isotope discrimination in leaf and stem sugars, water-use efficiency and mesophyll conductance during different developmental stages in rice subjected to drought. Aust. J. Plant Physiol. 1998, 25, 489–498. [Google Scholar] [CrossRef]
- Netting, A.G.; Theobald, J.C.; Dodd, I.C. Xylem sap collection and extraction methodologies to determine in vivo concentrations of ABA and its bound forms by gas chromatography-mass spectrometry (GC-MS). Plant Methods 2012, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jachetta, J.J.; Appleby, A.P.; Boersma, L. Use of the pressure vessel to measure concentrations of solutes in apoplastic and membrane-filtered symplastic sap in sunflower leaves. Plant Physiol. 1986, 82, 995–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bögelein, R.; Hassdenteufel, M.; Thomas, F.M.; Werner, W. Comparison of leaf gas exchange and stable isotope signature of water-soluble compounds along canopy gradients of co-occurring Douglas-fir and European beech. Plant Cell Environ. 2012, 35, 1245–1257. [Google Scholar] [CrossRef]
- Deslauriers, A.; Beaulieu, M.; Balducci, L.; Giovannelli, A.; Gagnon, M.J.; Rossi, S. Impact of warming and drought on carbon balance related to wood formation in black spruce. Ann. Bot. 2014, 114, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Oberhuber, W.; Swidrak, I.; Pirkebner, D.; Gruber, A. Temporal dynamics of nonstructural carbohydrates and xylem growth in Pinus sylvestris exposed to drought. Can. J. For. Res. 2011, 41, 1590–1597. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G.; Stoll, M.; Santos, T.; de Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef]
- Sun, P.; Wahbi, S.; Tsonev, T.; Haworth, M.; Liu, S.; Centritto, M. On the use of leaf spectral indices to assess water status and photosynthetic limitations in Olea europaea L. during water-stress and recovery. PLoS ONE 2014, 9, e105165. [Google Scholar] [CrossRef] [Green Version]
- Galmes, J.; Flexas, J.; Keys, A.J.; Cifre, J.; Mitchell, R.A.C.; Madgwick, P.J.; Haslam, R.P.; Medrano, H.; Parry, M.A.J. Rubisco specificity factor tends to be larger in plant species from drier habitats and in species with persistent leaves. Plant Cell Environ. 2005, 28, 571–579. [Google Scholar] [CrossRef]
- Kok, B. A critical consideration of the quantum yield of Chlorella photosynthesis. Enzymologia 1948, 13, 1–56. [Google Scholar]
- Laisk, A.; Loreto, F. Determining photosynthetic parameters from leaf CO2 exchange and chlorophyll fluorescence—ibulose-1,5-bisphosphate carboxylase oxygenase specificity factor, dark respiration in the light, excitation distribution between photosystems, alternative electron transport rate, and mesophyll diffusion resistance. Plant Physiol. 1996, 110, 903–912. [Google Scholar] [PubMed]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Haworth, M.; Marino, G.; Centritto, M. An introductory guide to gas exchange analysis of photosynthesis and its application to plant phenotyping and precision irrigation to enhance water use efficiency. J. Water Clim. Chang. 2018, 9, 786–808. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Von Caemmerer, S.; Evans, J.R. Determination of the average partial pressure of CO2 in chloroplasts from leaves of several C3 plants. Aust. J. Plant Physiol. 1991, 18, 287–305. [Google Scholar] [CrossRef]
- Dufrêne, E.; Pontailler, J.; Saugier, B. A branch bag technique for simultaneous CO2 enrichment and assimilation measurements on beech (Fagus sylvatica L.). Plant Cell Environ. 1993, 16, 1131–1138. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar δ13C (‰) | Gm C-Isotopic Sugars (mol m−2 s−1 bar−1) | Gm Variable J (mol m−2 s−1 bar−1) | ||||
|---|---|---|---|---|---|---|
| Control | Drought | Control | Drought | Control | Drought | |
| Leaf | −24.700 ± 0.178C | −23.355 ± 0.189B | 0.333 ± 0.045a | 0.039 ± 0.010d | 0.278 ± 0.038ab | 0.021 ± 0.008d |
| Leaf petiole sap | −23.166 ± 0.146B | −22.500 ± 0.132A | 0.228 ± 0.031b | 0.026 ± 0.006d | - | - |
| Branch sap | −24.427 ± 0.116C | −23.450 ± 0.199B | 0.287 ± 0.054ab | 0.125 ± 0.011c | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, G.; Haworth, M.; Scartazza, A.; Tognetti, R.; Centritto, M. A Comparison of the Variable J and Carbon-Isotopic Composition of Sugars Methods to Assess Mesophyll Conductance from the Leaf to the Canopy Scale in Drought-Stressed Cherry. Int. J. Mol. Sci. 2020, 21, 1222. https://doi.org/10.3390/ijms21041222
Marino G, Haworth M, Scartazza A, Tognetti R, Centritto M. A Comparison of the Variable J and Carbon-Isotopic Composition of Sugars Methods to Assess Mesophyll Conductance from the Leaf to the Canopy Scale in Drought-Stressed Cherry. International Journal of Molecular Sciences. 2020; 21(4):1222. https://doi.org/10.3390/ijms21041222
Chicago/Turabian StyleMarino, Giovanni, Matthew Haworth, Andrea Scartazza, Roberto Tognetti, and Mauro Centritto. 2020. "A Comparison of the Variable J and Carbon-Isotopic Composition of Sugars Methods to Assess Mesophyll Conductance from the Leaf to the Canopy Scale in Drought-Stressed Cherry" International Journal of Molecular Sciences 21, no. 4: 1222. https://doi.org/10.3390/ijms21041222