Identification and Physicochemical Characterization of a New Allergen from Ascaris lumbricoides

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
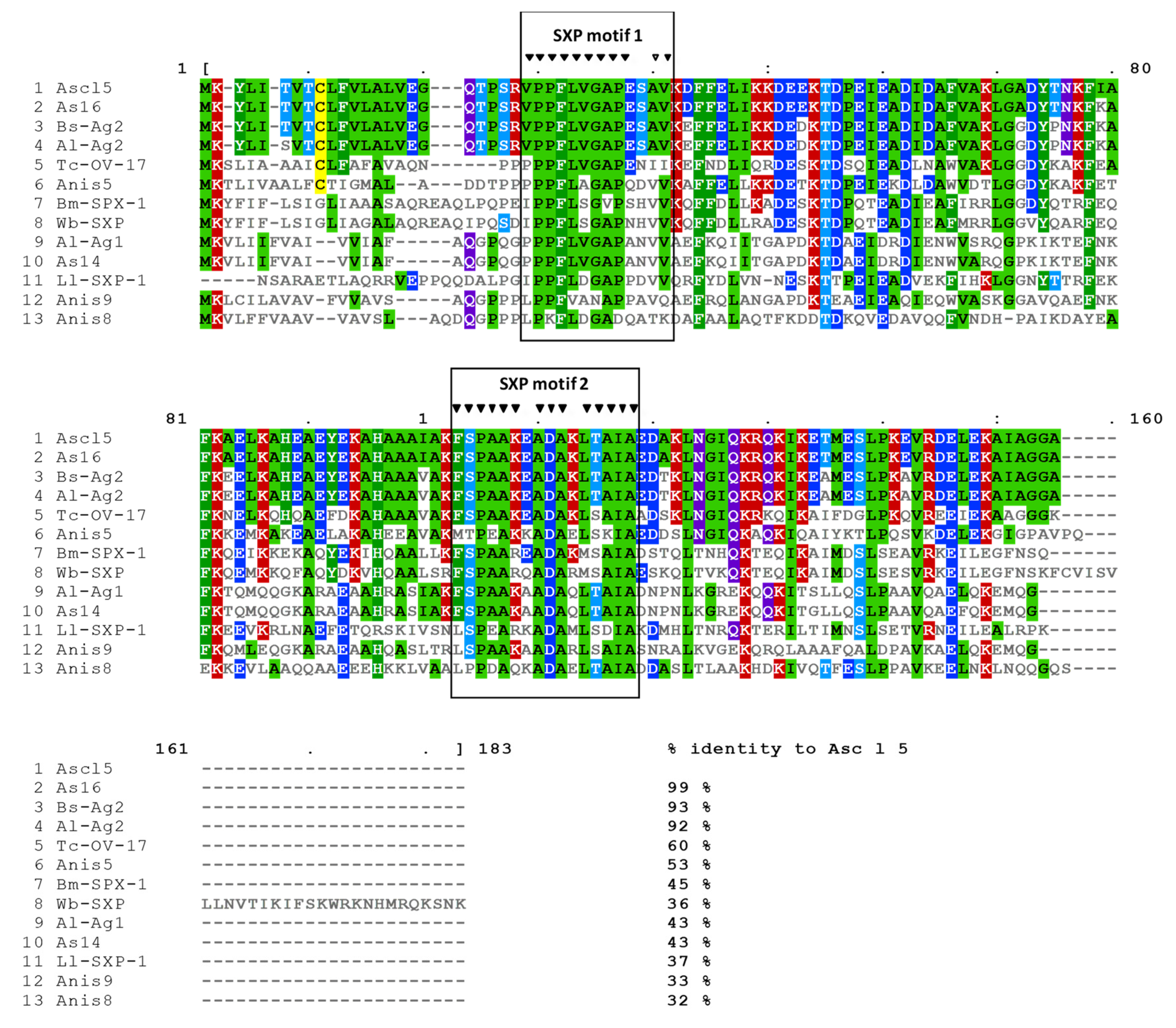
2.1. Asc l 5 Belongs to the SXP/RAL-2 Protein Family
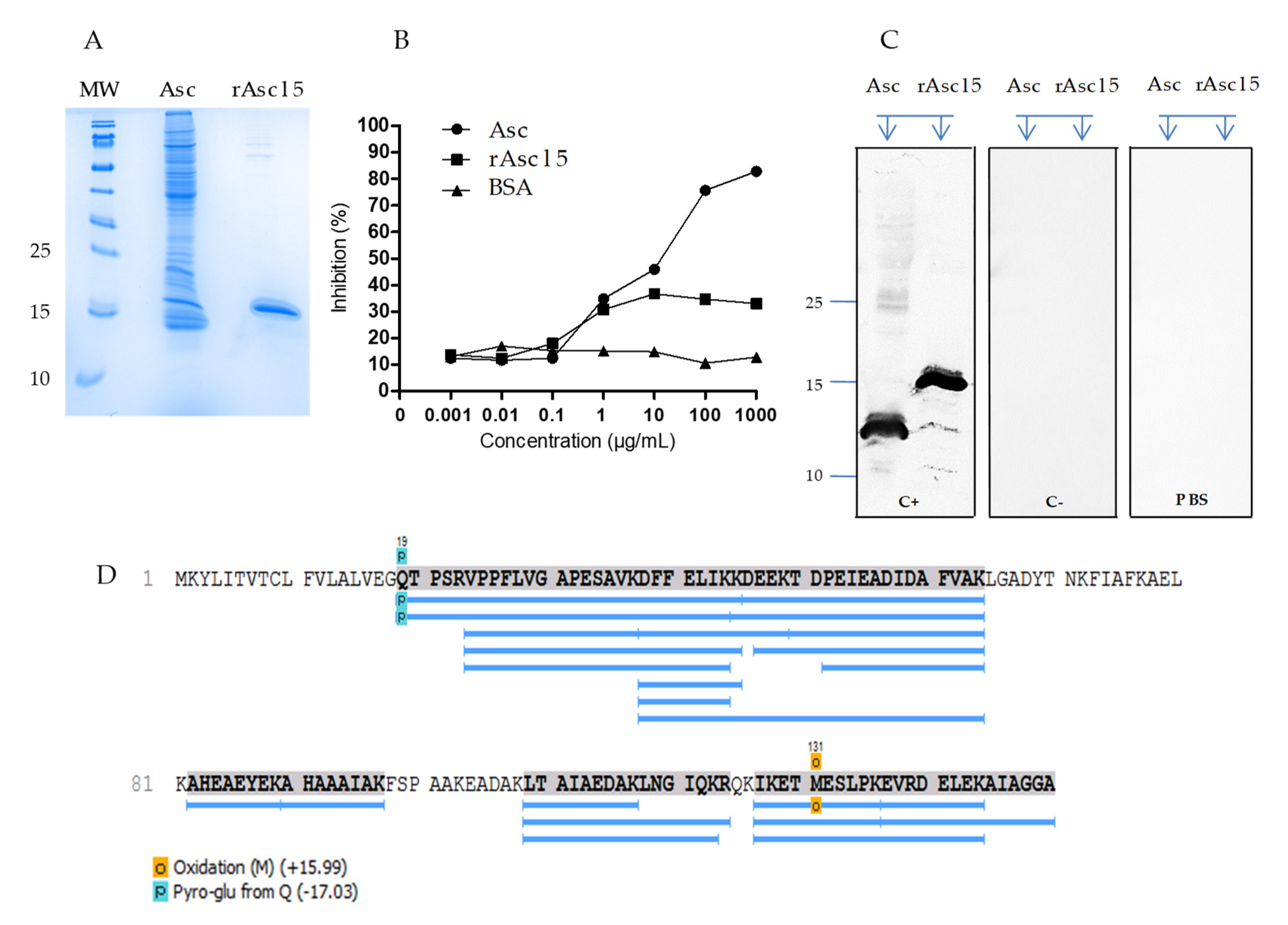
2.2. Natural Asc l 5 in the A. lumbricoides Extract
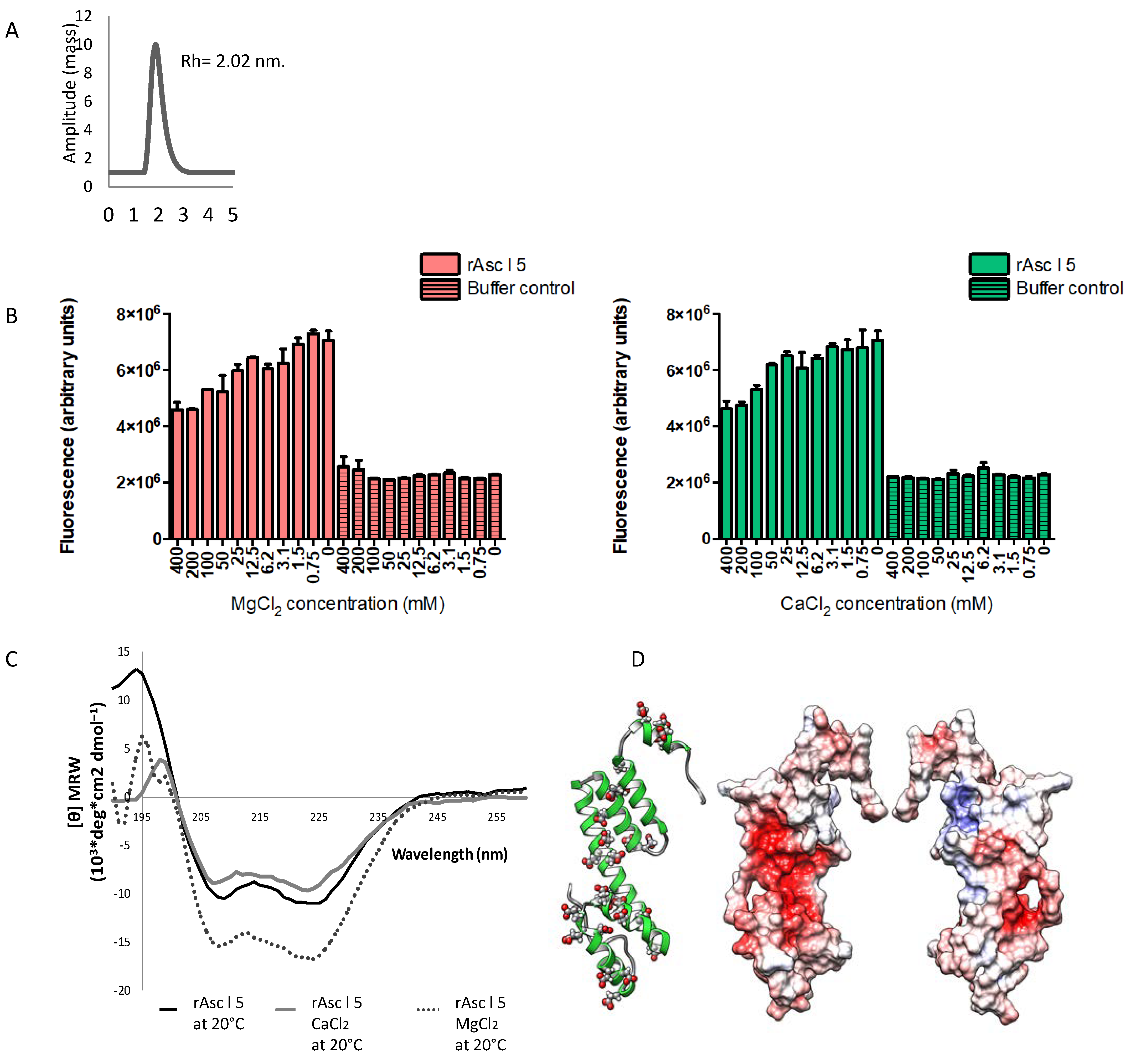
2.3. rAsc l 5 Is a Monomeric Protein
2.4. Calcium and Magnesium Induce Structural Changes in rAsc l 5
2.5. Three-Dimensional Structure Model of Asc l 5 Comprises Six Alpha Helices with Negatively Charged Patches and Predicted Calcium and Magnesium-Binding Sites
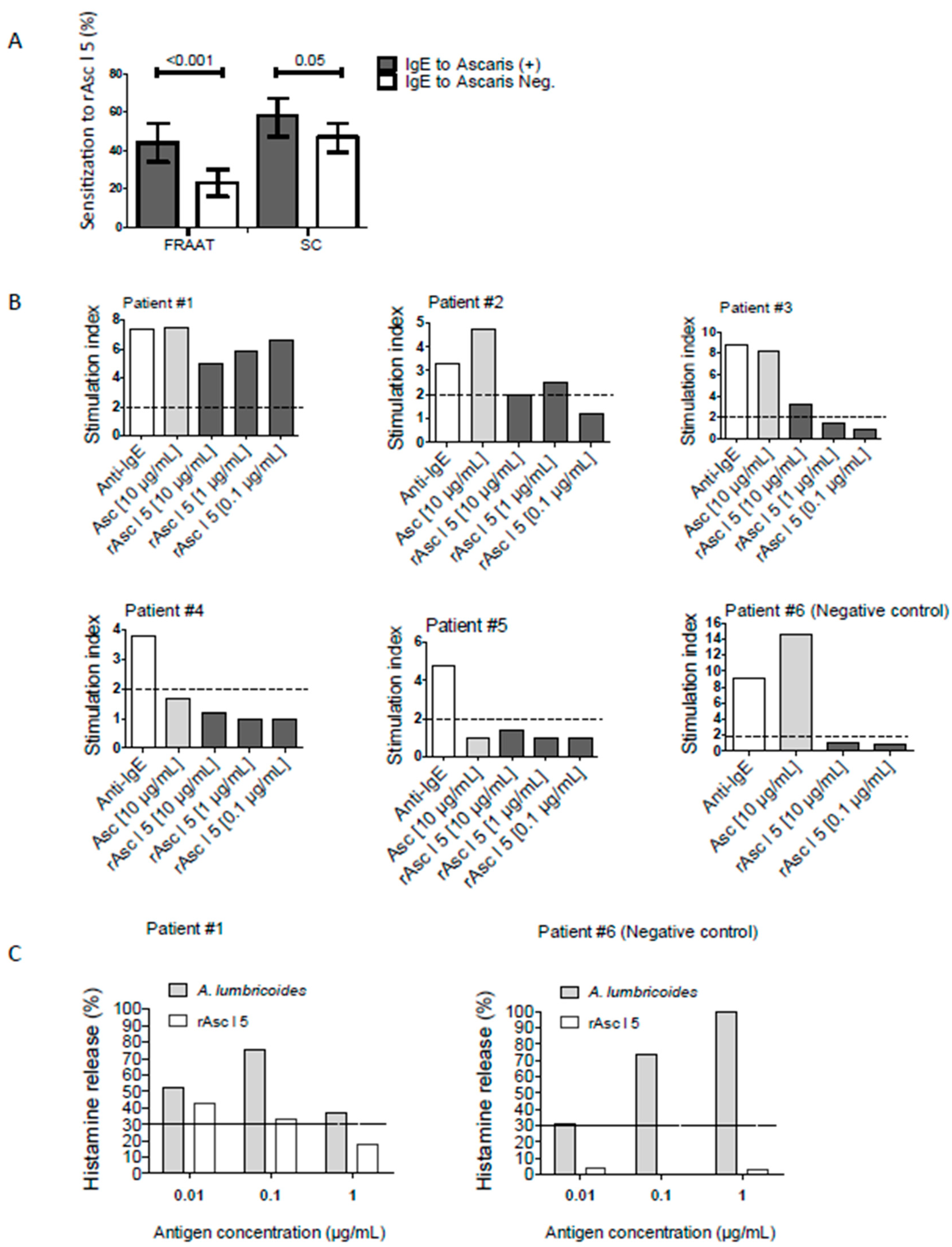
2.6. rAsc l 5 Binds IgE, Activates Basophils and Induces Histamine Release
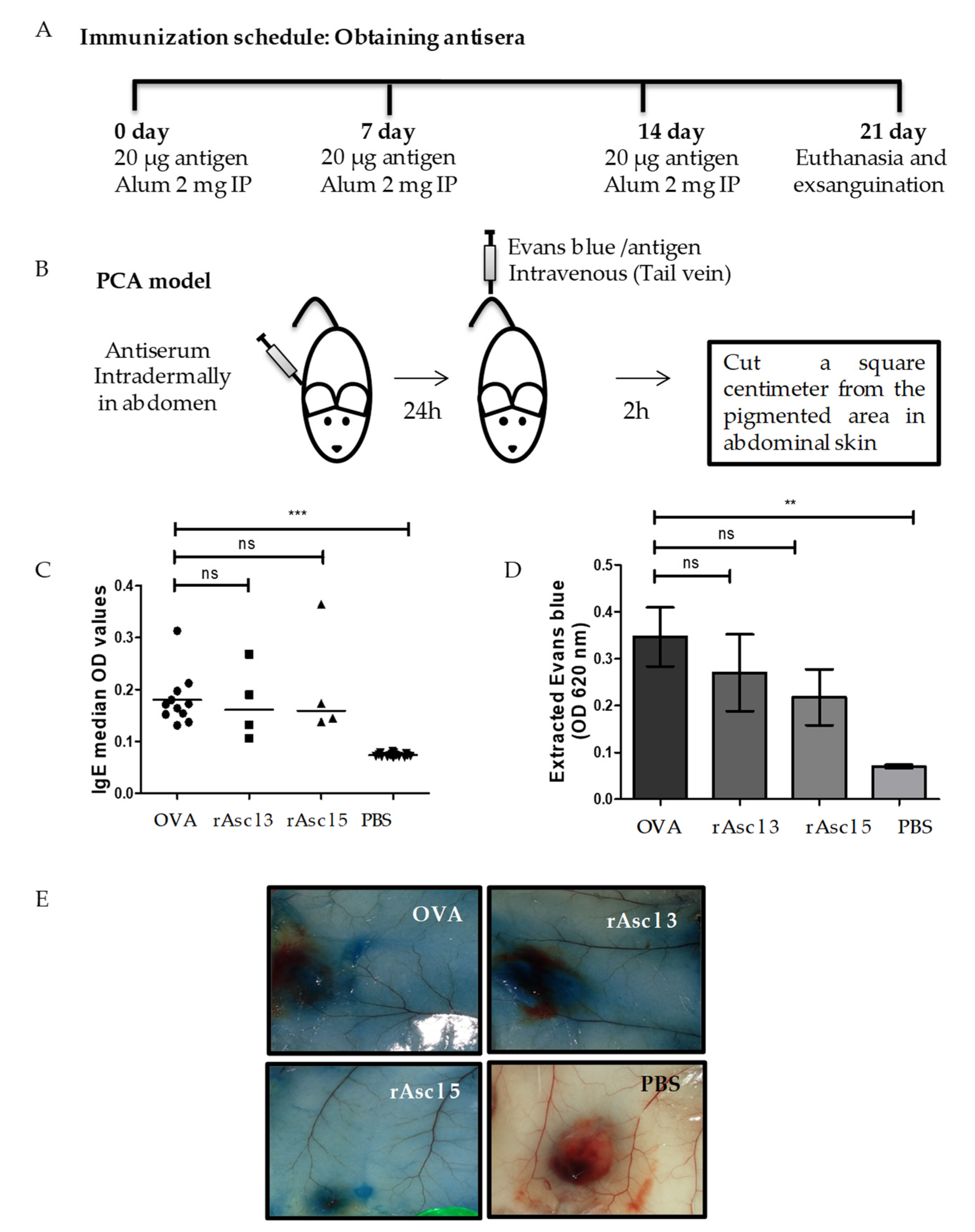
2.7. rAsc l 5 Induces Specific IgE Production in Immunized Mice and A Positive Passive Cutaneous Anaphylaxis (PCA) Reaction
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Ethical Statement
4.3. IgE Immunoscreening of A. lumbricoides cDNA Library
4.4. Isolation, Sequencing, and Nucleotide Analysis of IgE-Binding Clones
4.5. Cloning, Expression, and Purification of Recombinant Asc l 5
4.6. A. lumbricoides Extract Preparation, Enzyme-Linked Immunosorbent Assay (ELISA) Inhibition and Western Blot (WB)
4.7. Peptide Analysis by Nano-Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS)
4.8. Size Exclusion Chromatography, Amino Acid Composition, Aggregation Behavior and Fourier Transform Infrared (FTIR) Spectrosocpy Experiments
4.9. Calcium and Magnesium-Binding Experiments and Circular Dichroism (CD) Spectroscopy
4.10. Modeling of Three-Dimensional Structure of Asc l 5 and Ca2+ and Mg2+ Binding Sites
4.11. Specific IgE
4.12. Basophil Activation Test
4.13. Histamine Release
4.14. Production of Antisera against rAsc l 5, Antibody Determinations and PCA Model
4.15. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IPTG | isopropyl β-D-1-thiogalactopyranoside |
| PBS | phosphate buffered saline |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| nano-LC-MS/MS | Nanoscale liquid chromatography coupled to tandem mass spectrometry |
| CD | Circular dichroism |
| DLS | Dynamic light scattering |
| ELISA | Enzyme-linked immunosorbent assay |
| SPT | skin prick test |
| E. coli | Escherichia coli |
| LB | Luria Bertani |
| DUF | domain of unknown function |
| HDM | House dust mite |
| IUIS | International Union of Immunological Societies |
References
- Hagel, I.; Cabrera, M.; Hurtado, M.A.; Sanchez, P.; Puccio, F.; Di Prisco, M.C.; Palenque, M. Infection by Ascaris lumbricoides and bronchial hyper reactivity: An outstanding association in Venezuelan school children from endemic areas. Acta Trop. 2007, 103, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Moncayo, A.L.; Vaca, M.; Oviedo, G.; Workman, L.J.; Chico, M.E.; Platts-Mills, T.A.; Rodrigues, L.C.; Barreto, M.L.; Cooper, P.J. Effects of geohelminth infection and age on the associations between allergen-specific IgE, skin test reactivity and wheeze: A case-control study. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2013, 43, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caraballo, L. The tropics, helminth infections and hygiene hypotheses. Expert Rev. Clin. Immunol. 2018, 14, 99–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weatherhead, J.E.; Porter, P.; Coffey, A.; Haydel, D.; Versteeg, L.; Zhan, B.; Gazzinelli Guimaraes, A.C.; Fujiwara, R.; Jaramillo, A.M.; Bottazzi, M.E.; et al. Ascaris Larval Infection and Lung Invasion Directly Induce Severe Allergic Airway Disease in Mice. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, N.; Caraballo, L. IgE cross-reactivity between Ascaris lumbricoides and mite allergens: Possible influences on allergic sensitization and asthma. Parasite Immunol. 2011, 33, 309–321. [Google Scholar] [CrossRef]
- Acevedo, N.; Sanchez, J.; Erler, A.; Mercado, D.; Briza, P.; Kennedy, M.; Fernandez, A.; Gutierrez, M.; Chua, K.Y.; Cheong, N.; et al. IgE cross-reactivity between Ascaris and domestic mite allergens: The role of tropomyosin and the nematode polyprotein ABA-1. Allergy 2009, 64, 1635–1643. [Google Scholar] [CrossRef]
- Caraballo, L.; Acevedo, N.; Zakzuk, J. Ascariasis as a model to study the helminth/allergy relationships. Parasite Immunol. 2019, 41, e12595. [Google Scholar] [CrossRef]
- Acevedo, N.; Mohr, J.; Zakzuk, J.; Samonig, M.; Briza, P.; Erler, A.; Pomes, A.; Huber, C.G.; Ferreira, F.; Caraballo, L. Proteomic and immunochemical characterization of glutathione transferase as a new allergen of the nematode Ascaris lumbricoides. PLoS ONE 2013, 8, e78353. [Google Scholar] [CrossRef]
- Acevedo, N.; Erler, A.; Briza, P.; Puccio, F.; Ferreira, F.; Caraballo, L. Allergenicity of Ascaris lumbricoides tropomyosin and IgE sensitization among asthmatic patients in a tropical environment. Int. Arch. Allergy Immunol. 2011, 154, 195–206. [Google Scholar] [CrossRef]
- Caraballo, L.; Coronado, S. Parasite allergens. Mol. Immunol. 2018, 100, 113–119. [Google Scholar] [CrossRef]
- Caraballo, L.; Acevedo, N. New Allergens of Relevance in Tropical Regions: The Impact of Ascaris lumbricoides Infections. World Allergy Organ. J. 2011, 4, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakzuk, J.; Acevedo, N.; Cifuentes, L.; Bornacelly, A.; Sanchez, J.; Ahumada, V.; Ring, J.; Ollert, M.; Caraballo, L. Early life IgE responses in children living in the tropics: A prospective analysis. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatric Allergy Immunol. 2013, 24, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Caraballo, L. Mite allergens. Expert Rev. Clin. Immunol. 2017, 13, 297–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Mayoral, M.F.; Trevino, M.A.; Perez-Pinar, T.; Caballero, M.L.; Knaute, T.; Umpierrez, A.; Bruix, M.; Rodriguez-Perez, R. Relationships between IgE/IgG4 epitopes, structure and function in Anisakis simplex Ani s 5, a member of the SXP/RAL-2 protein family. PLoS Negl. Trop. Dis. 2014, 8, e2735. [Google Scholar] [CrossRef]
- Stigler, J.; Rief, M. Calcium-dependent folding of single calmodulin molecules. Proc. Natl. Acad. Sci. USA 2012, 109, 17814–17819. [Google Scholar] [CrossRef] [Green Version]
- Gabellieri, E.; Strambini, G.B. ANS fluorescence detects widespread perturbations of protein tertiary structure in ice. Biophys. J. 2006, 90, 3239–3245. [Google Scholar] [CrossRef] [Green Version]
- Kirberger, M.; Yang, J.J. Calcium-Binding Protein Site Types. In Encyclopedia of Metalloproteins; Kretsinger, R.H., Uversky, V.N., Permyakov, E.A., Eds.; Springer: New York, NY, USA, 2013; pp. 511–521. [Google Scholar] [CrossRef]
- Zheng, H.; Cooper, D.R.; Porebski, P.J.; Shabalin, I.G.; Handing, K.B.; Minor, W. CheckMyMetal: A macromolecular metal-binding validation tool. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Caraballo, L.; Zakzuk, J.; Lee, B.W.; Acevedo, N.; Soh, J.Y.; Sanchez-Borges, M.; Hossny, E.; Garcia, E.; Rosario, N.; Ansotegui, I.; et al. Particularities of allergy in the Tropics. World Allergy Organ. J. 2016, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Ahumada, V.; Garcia, E.; Dennis, R.; Rojas, M.X.; Rondon, M.A.; Perez, A.; Penaranda, A.; Barragan, A.M.; Jimenez, S.; Kennedy, M.W.; et al. IgE responses to Ascaris and mite tropomyosins are risk factors for asthma. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2015, 45, 1189–1200. [Google Scholar] [CrossRef]
- Zakzuk, J.; Mercado, D.; Bornacelly, A.; Sanchez, J.; Ahumada, V.; Acevedo, N.; Caraballo, L. Hygienic conditions influence sensitization to Blomia tropicalis allergenic components: Results from the FRAAT birth cohort. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2019, 30, 172–178. [Google Scholar] [CrossRef]
- Buendia, E.; Zakzuk, J.; Mercado, D.; Alvarez, A.; Caraballo, L. The IgE response to Ascaris molecular components is associated with clinical indicators of asthma severity. World Allergy Organ. J. 2015, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, N.; Suzuki, K.; Kasuga-Aoki, H.; Matsumoto, Y.; Arakawa, T.; Ishiwata, K.; Isobe, T. Intranasal immunization with recombinant Ascaris suum 14-kilodalton antigen coupled with cholera toxin B subunit induces protective immunity to A. suum infection in mice. Infect. Immun. 2001, 69, 7285–7292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Versteeg, L.; Liu, Z.; Keegan, B.; Gazzinelli-Guimaraes, A.C.; Fujiwara, R.T.; Briggs, N.; Jones, K.M.; Strych, U.; Beaumier, C.M.; et al. Yeast-expressed recombinant As16 protects mice against Ascaris suum infection through induction of a Th2-skewed immune response. PLoS Negl. Trop. Dis. 2017, 11, e0005769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, R.T.; Zhan, B.; Mendez, S.; Loukas, A.; Bueno, L.L.; Wang, Y.; Plieskatt, J.; Oksov, Y.; Lustigman, S.; Bottazzi, M.E.; et al. Reduction of worm fecundity and canine host blood loss mediates protection against hookworm infection elicited by vaccination with recombinant Ac-16. Clin. Vaccine Immunol. 2007, 14, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.V.; Eswaran, M.; Ravi, V.; Gnanasekhar, B.; Narayanan, R.B.; Kaliraj, P.; Jayaraman, K.; Marson, A.; Raghavan, N.; Scott, A.L. The Wuchereria bancrofti orthologue of Brugia malayi SXP1 and the diagnosis of bancroftian filariasis. Mol. Biochem. Parasitol. 2000, 107, 71–80. [Google Scholar] [CrossRef]
- Klion, A.D.; Vijaykumar, A.; Oei, T.; Martin, B.; Nutman, T.B. Serum immunoglobulin G4 antibodies to the recombinant antigen, Ll-SXP-1, are highly specific for Loa loa infection. J. Infect. Dis. 2003, 187, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Ishizaki, S.; Shimakura, K.; Nagashima, Y.; Shiomi, K. Molecular cloning and expression of two new allergens from Anisakis simplex. Parasitol. Res. 2007, 100, 1233–1241. [Google Scholar] [CrossRef]
- Rodriguez-Perez, R.; Moneo, I.; Rodriguez-Mahillo, A.; Caballero, M.L. Cloning and expression of Ani s 9, a new Anisakis simplex allergen. Mol. Biochem. Parasitol. 2008, 159, 92–97. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Shimakura, K.; Ishizaki, S.; Nagashima, Y.; Shiomi, K. Purification and cDNA cloning of a new heat-stable allergen from Anisakis simplex. Mol. Biochem. Parasitol. 2007, 155, 138–145. [Google Scholar] [CrossRef]
- Finn, B.E.; Forsen, S. The evolving model of calmodulin structure, function and activation. Structure 1995, 3, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Yamniuk, A.P.; Vogel, H.J. Calmodulin’s flexibility allows for promiscuity in its interactions with target proteins and peptides. Mol. Biotechnol. 2004, 27, 33–57. [Google Scholar] [CrossRef]
- Yap, K.L.; Kim, J.; Truong, K.; Sherman, M.; Yuan, T.; Ikura, M. Calmodulin target database. J. Struct. Funct. Genom. 2000, 1, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.K.; Einsle, O. Role of Calcium in Secondary Structure Stabilization during Maturation of Nitrous Oxide Reductase. Biochemistry 2016, 55, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Dubiela, P.; Aina, R.; Polak, D.; Geiselhart, S.; Humeniuk, P.; Bohle, B.; Alessandri, S.; Del Conte, R.; Cantini, F.; Borowski, T.; et al. Enhanced Pru p 3 IgE-binding activity by selective free fatty acid-interaction. J. Allergy Clin. Immunol. 2017, 140, 1728–1731.e1710. [Google Scholar] [CrossRef] [Green Version]
- Dubiela, P.; Del Conte, R.; Cantini, F.; Borowski, T.; Aina, R.; Radauer, C.; Bublin, M.; Hoffmann-Sommergruber, K.; Alessandri, S. Impact of lipid binding on the tertiary structure and allergenic potential of Jug r 3, the non-specific lipid transfer protein from walnut. Sci. Rep. 2019, 9, 2007. [Google Scholar] [CrossRef]
- Engel, E.; Richter, K.; Obermeyer, G.; Briza, P.; Kungl, A.J.; Simon, B.; Auer, M.; Ebner, C.; Rheinberger, H.J.; Breitenbach, M.; et al. Immunological and biological properties of Bet v 4, a novel birch pollen allergen with two EF-hand calcium-binding domains. J. Biol. Chem. 1997, 272, 28630–28637. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.T.; Costa, V.A.; Pereira, T.G.; Sales, I.R.; Silva, S.F.; Maciel, M.A.; Malagueño, E.; Souza, V.M. Severity of atopic dermatitis and Ascaris lumbricoides infection: An evaluation of CCR4+ and CXCR3+ helper T cell frequency. Rev. Soc. Bras. Med. Trop. 2012, 45, 761–763. [Google Scholar] [CrossRef] [Green Version]
- Zakzuk, J.; Casadiego, S.; Mercado, A.; Alvis-Guzman, N.; Caraballo, L. Ascaris lumbricoides infection induces both, reduction and increase of asthma symptoms in a rural community. Acta Trop. 2018, 187, 1–4. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Suzuki, S.; Nozoye, T.; Yamakawa, T.; Takashima, Y.; Arakawa, T.; Tsuji, N.; Takaiwa, F.; Hayashi, Y. Oral immunogenicity and protective efficacy in mice of transgenic rice plants producing a vaccine candidate antigen (As16) of Ascaris suum fused with cholera toxin B subunit. Transgenic Res. 2009, 18, 185–192. [Google Scholar] [CrossRef]
- Tsuji, N.; Miyoshi, T.; Islam, M.K.; Isobe, T.; Yoshihara, S.; Arakawa, T.; Matsumoto, Y.; Yokomizo, Y. Recombinant Ascaris 16-Kilodalton protein-induced protection against Ascaris suum larval migration after intranasal vaccination in pigs. J. Infect. Dis. 2004, 190, 1812–1820. [Google Scholar] [CrossRef] [Green Version]
- Diemert, D.J.; Pinto, A.G.; Freire, J.; Jariwala, A.; Santiago, H.; Hamilton, R.G.; Periago, M.V.; Loukas, A.; Tribolet, L.; Mulvenna, J.; et al. Generalized urticaria induced by the Na-ASP-2 hookworm vaccine: Implications for the development of vaccines against helminths. J. Allergy Clin. Immunol. 2012, 130, 169–176 e166. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, N.; Sanchez, J.; Zakzuk, J.; Bornacelly, A.; Quiroz, C.; Alvarez, A.; Puello, M.; Mendoza, K.; Martinez, D.; Mercado, D.; et al. Particular characteristics of allergic symptoms in tropical environments: Follow up to 24 months in the FRAAT birth cohort study. BMC Pulm. Med. 2012, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Dennis, R.J.; Caraballo, L.; Garcia, E.; Rojas, M.X.; Rondon, M.A.; Perez, A.; Aristizabal, G.; Penaranda, A.; Barragan, A.M.; Ahumada, V.; et al. Prevalence of asthma and other allergic conditions in Colombia 2009-2010: A cross-sectional study. BMC Pulm. Med. 2012, 12, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronado, S.; Barrios, L.; Zakzuk, J.; Regino, R.; Ahumada, V.; Franco, L.; Ocampo, Y.; Caraballo, L. A recombinant cystatin from Ascaris lumbricoides attenuates inflammation of DSS-induced colitis. Parasite Immunol. 2017, 39. [Google Scholar] [CrossRef]
- Wolf, M.; Twaroch, T.E.; Huber, S.; Reithofer, M.; Steiner, M.; Aglas, L.; Hauser, M.; Aloisi, I.; Asam, C.; Hofer, H.; et al. Amb a 1 isoforms: Unequal siblings with distinct immunological features. Allergy 2017, 72, 1874–1882. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [Green Version]
- Laimer, J.; Hofer, H.; Fritz, M.; Wegenkittl, S.; Lackner, P. MAESTRO--multi agent stability prediction upon point mutations. BMC Bioinform. 2015, 16, 116. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. A Publ. Protein Soc. 2018, 27, 112–128. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Breindel, C.; Saraswat, D.; Cullen, P.J.; Edgerton, M. Candida albicans Sap6 amyloid regions function in cellular aggregation and zinc binding, and contribute to zinc acquisition. Sci. Rep. 2017, 7, 2908. [Google Scholar] [CrossRef]
- Lin, Y.F.; Cheng, C.W.; Shih, C.S.; Hwang, J.K.; Yu, C.S.; Lu, C.H. MIB: Metal Ion-Binding Site Prediction and Docking Server. J. Chem. Inf. Modeling 2016, 56, 2287–2291. [Google Scholar] [CrossRef] [Green Version]
- MacGlashan, D.W., Jr. Basophil activation testing. J. Allergy Clin. Immunol. 2013, 132, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.; Munera, M.; Cantillo, J.F.; Wortmann, J.; Zakzuk, J.; Keller, W.; Caraballo, L.; Puerta, L. An Engineered Hybrid Protein from Dermatophagoides pteronyssinus Allergens Shows Hypoallergenicity. Int. J. Mol. Sci. 2019, 20, 3025. [Google Scholar] [CrossRef] [Green Version]
- Zakzuk, J.; Benedetti, I.; Fernandez-Caldas, E.; Caraballo, L. The influence of chitin on the immune response to the house dust mite allergen Blo T 12. Int. Arch. Allergy Immunol. 2014, 163, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Sun, N.; Li, Y.; Jia, X. A BALB/c mouse model for assessing the potential allergenicity of proteins: Comparison of allergen dose, sensitization frequency, timepoint and sex. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 62, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, S.B.; Reish, R.; Fernandes, J.; Gaudry, P.; Dai, G.; Reese, G. Enhancement of murine IgE antibody detection by IgG removal. J. Immunol. Methods 2004, 284, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.; Killoran, K.E.; Mitre, E. Measuring local anaphylaxis in mice. J. Vis. Exp. 2014, 14, e52005. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Asthmatics, n (%) | Non Asthmatics, n (%) | p Value |
|---|---|---|---|
| n = 50 | n = 248 | ||
| Asc l 5 sensitization 1 | 29 (58.0%) | 126 (50.8%) | 0.353 |
| IgE to Asc l 5, OD 2 | 0.154 (0.114–0.253) | 0.146 (0.115–0.233) | 0.388 |
| Ascaris spp. sensitization 1 | 26 (50%) | 104 (42%) | 0.190 |
| IgE to Ascaris spp 2 | 0.444 (0.036–1.475) | 0.251 (0.07–1.134) | 0.821 |
| n = 48 | n = 242 | ||
| A. lumbricoides by stool examination 1 | 27 (56.3%) | 168 (69.4%) | 0.076 |
| A. lumbricoides epg 2 | 546 (0–2047) | 689 (0-2340) | 0.197 |
| n = 49 | n = 236 | ||
| Positive SPT to Ascaris 1 | 13 (26.5%) | 30 (12.7%) | 0.014 |
| Patients | ||||||
|---|---|---|---|---|---|---|
| Data | #1 | #2 | #3 | #4 | #5 | #6 |
| Code | ST371 | ST581 | ASC217 | ST383 | ST438 | ASC040 ɫ |
| Diagnosis | Asthma | Parasited | Asthma | Rhinitis | Asthma | Asthma |
| IgE to rAsc l 5 1 | 0.29 | 0.51 | 0.29 | 0.38 | 1.20 | 0.12 |
| IgE to Ascaris 2 | 16.2 | 8.19 | 3.64 | 0.30 | 0.03 | 9.02 |
| IgE to Blomia 2 | 22.80 | 0.67 | 0.48 | 2.01 | 0.10 | 1.33 |
| IgE to Dp. 2 | 2.20 | 0.10 | 0.31 | 7.04 | 0.02 | 1.16 |
| Total IgE 2 | 642.6 | 727.1 | 588.3 | 478.7 | 228.5 | 975.8 |
| Gender | F | F | M | F | F | F |
| Age | 50 | 61 | 35 | 36 | 3 | 28 |
| Stimulation index (SI) of CD203 based-basophil activation test | ||||||
| Anti-IgE | 7.4 | 3.3 | 8.7 | 3.8 | 4.8 | 9.0 |
| rAsc l 5 10 µg/mL | 5.0 | 2.0 | 3.2 | 1.2 | 1.4 | 1.0 |
| rAsc l 5 1 µg/mL | 5.9 | 2.5 | 1.4 | 1.0 | 1.0 | nd |
| rAsc l 5 0.1 µg/mL | 6.6 | 1.2 | 0.9 | 1.0 | 1.0 | 0.9 |
| Ascaris 10 µg/mL | 7.5 | 4.7 | 8.1 | 1.7 | 1.0 | 14.6 |
| Percentage of histamine release (%) | ||||||
| Ascaris 0.01 µg/mL | 52.2 | nd | nd | nd | nd | 31.0 |
| Ascaris 0.1 µg/mL | 75.2 | nd | nd | nd | nd | 73.0 |
| Ascaris 1 µg/mL | 37.0 | nd | nd | nd | nd | 100 |
| rAsc l 5 0.01 µg/mL | 42.3 | nd | nd | nd | nd | 4 |
| rAsc l 5 0.1 µg/mL | 33.2 | nd | nd | nd | nd | 0.4 |
| rAsc l 5 1 µg/mL | 18.0 | nd | nd | nd | nd | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahumada, V.; Manotas, M.; Zakzuk, J.; Aglas, L.; Coronado, S.; Briza, P.; Lackner, P.; Regino, R.; Araujo, G.; Ferreira, F.; et al. Identification and Physicochemical Characterization of a New Allergen from Ascaris lumbricoides. Int. J. Mol. Sci. 2020, 21, 9761. https://doi.org/10.3390/ijms21249761
Ahumada V, Manotas M, Zakzuk J, Aglas L, Coronado S, Briza P, Lackner P, Regino R, Araujo G, Ferreira F, et al. Identification and Physicochemical Characterization of a New Allergen from Ascaris lumbricoides. International Journal of Molecular Sciences. 2020; 21(24):9761. https://doi.org/10.3390/ijms21249761
Chicago/Turabian StyleAhumada, Velky, María Manotas, Josefina Zakzuk, Lorenz Aglas, Sandra Coronado, Peter Briza, Peter Lackner, Ronald Regino, Galber Araujo, Fatima Ferreira, and et al. 2020. "Identification and Physicochemical Characterization of a New Allergen from Ascaris lumbricoides" International Journal of Molecular Sciences 21, no. 24: 9761. https://doi.org/10.3390/ijms21249761