Erythroid Differentiation Regulator 1 Ameliorates Collagen-Induced Arthritis via Activation of Regulatory T Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
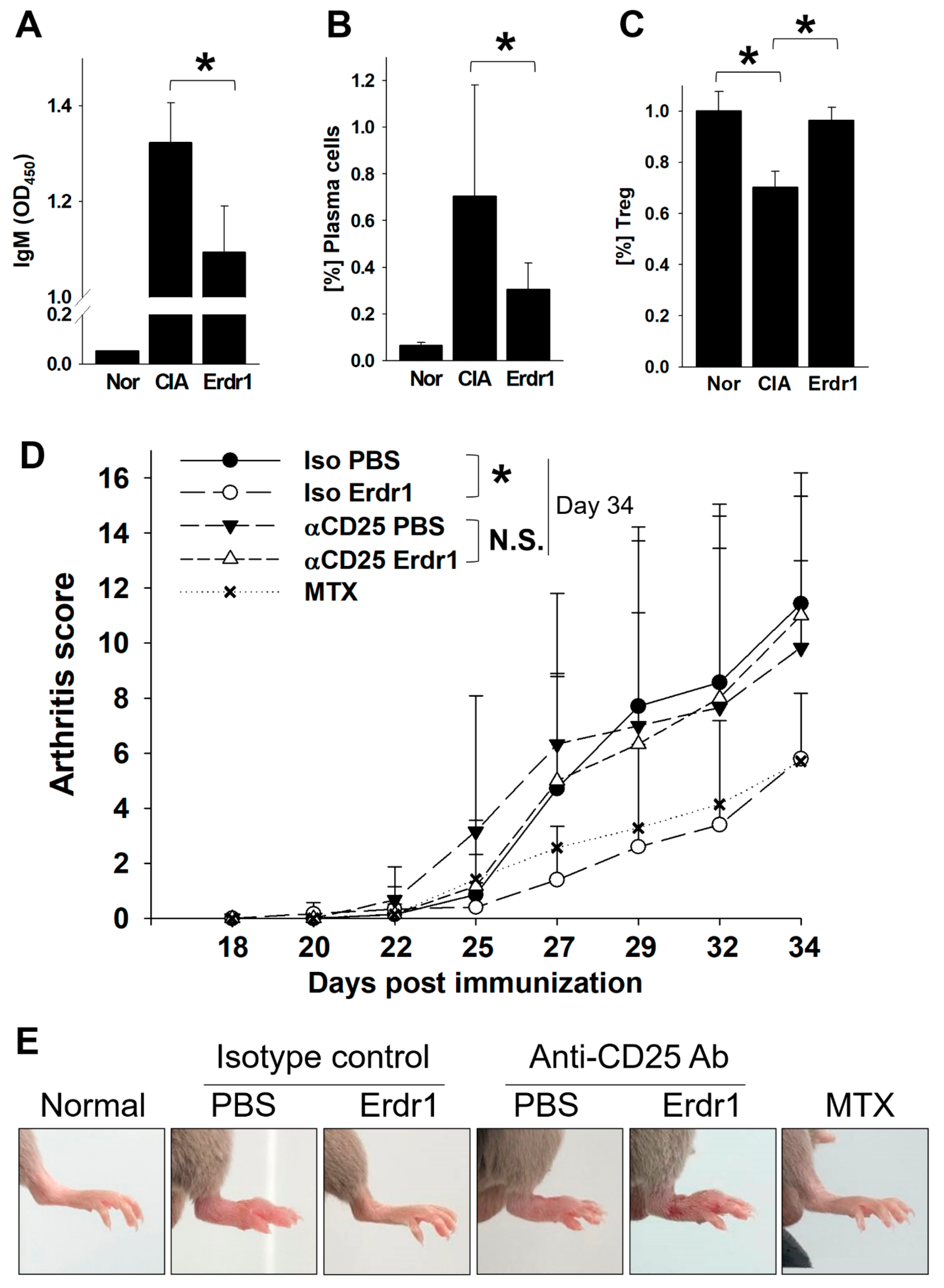
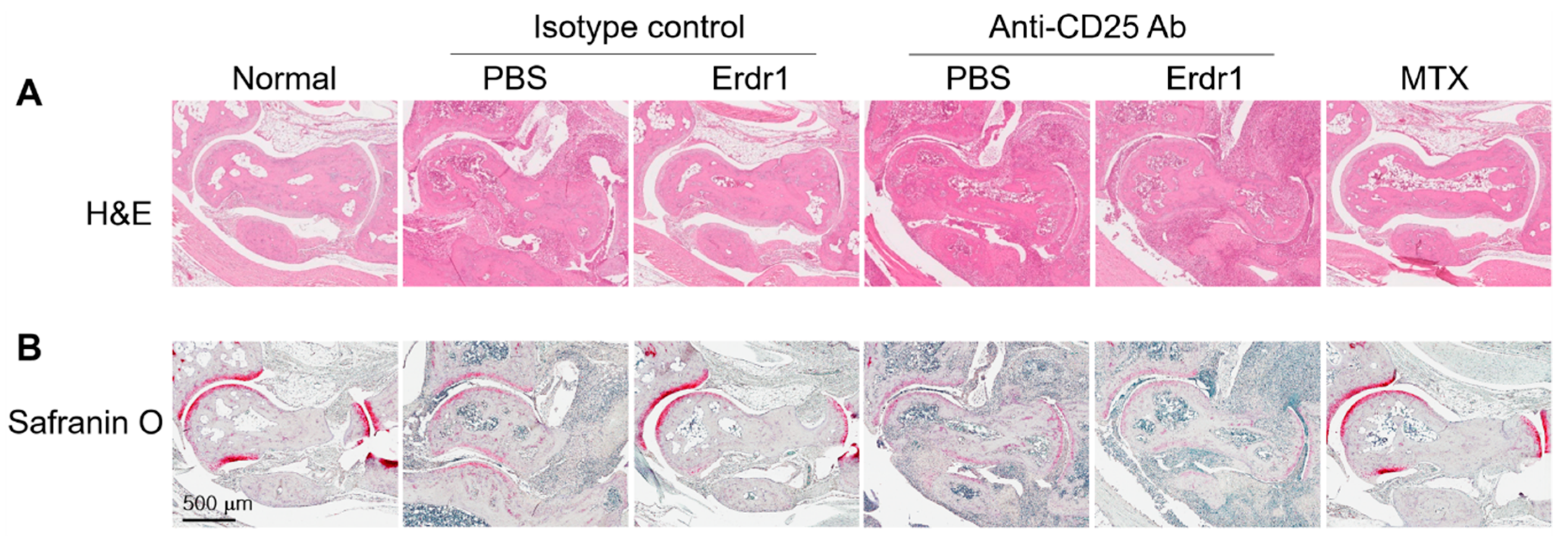
2.1. Erdr1 Ameliorates the Experimental Rheumatoid Arthritis via Treg Cells
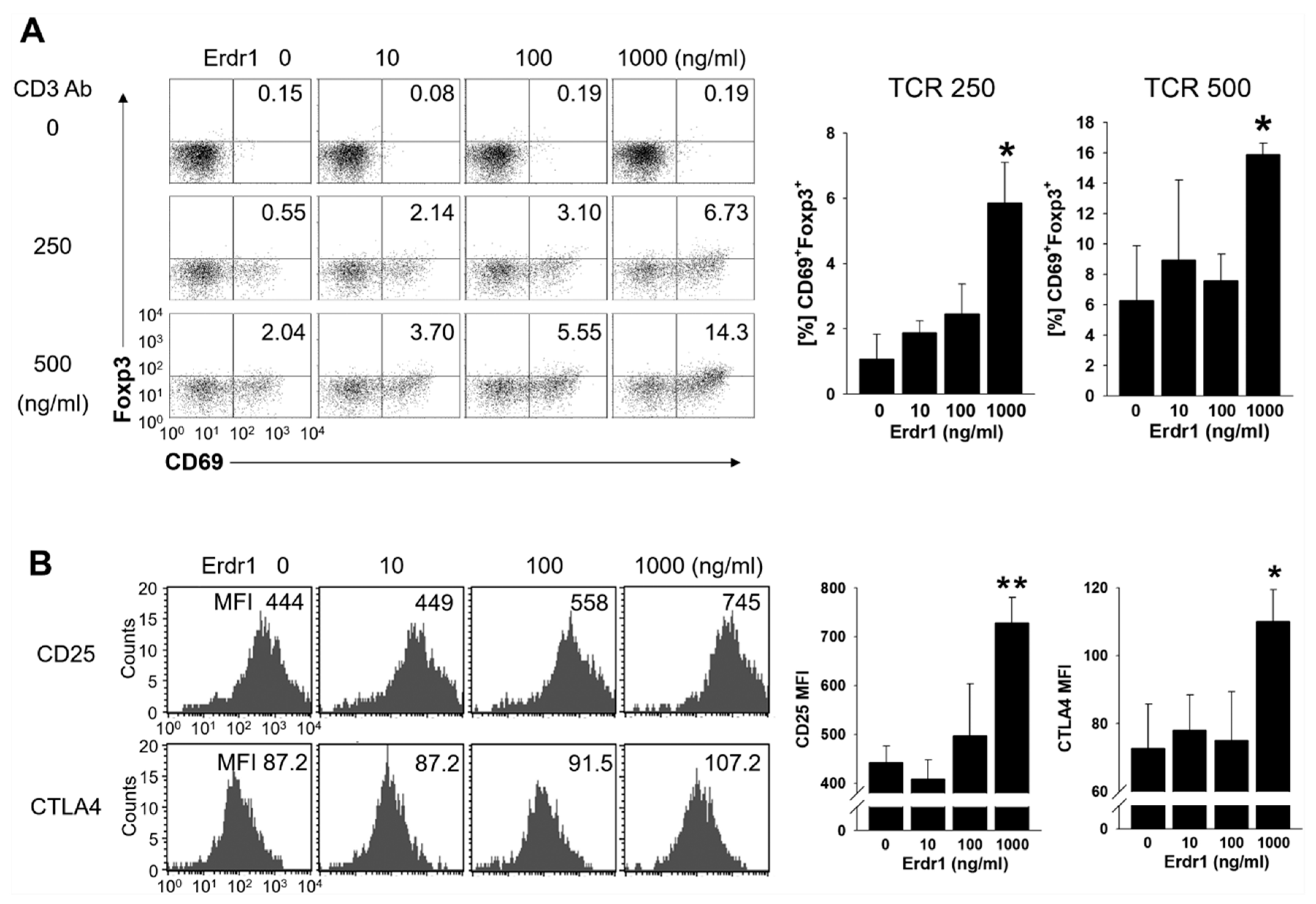
2.2. Erdr1 Induces Activation of Treg Cells
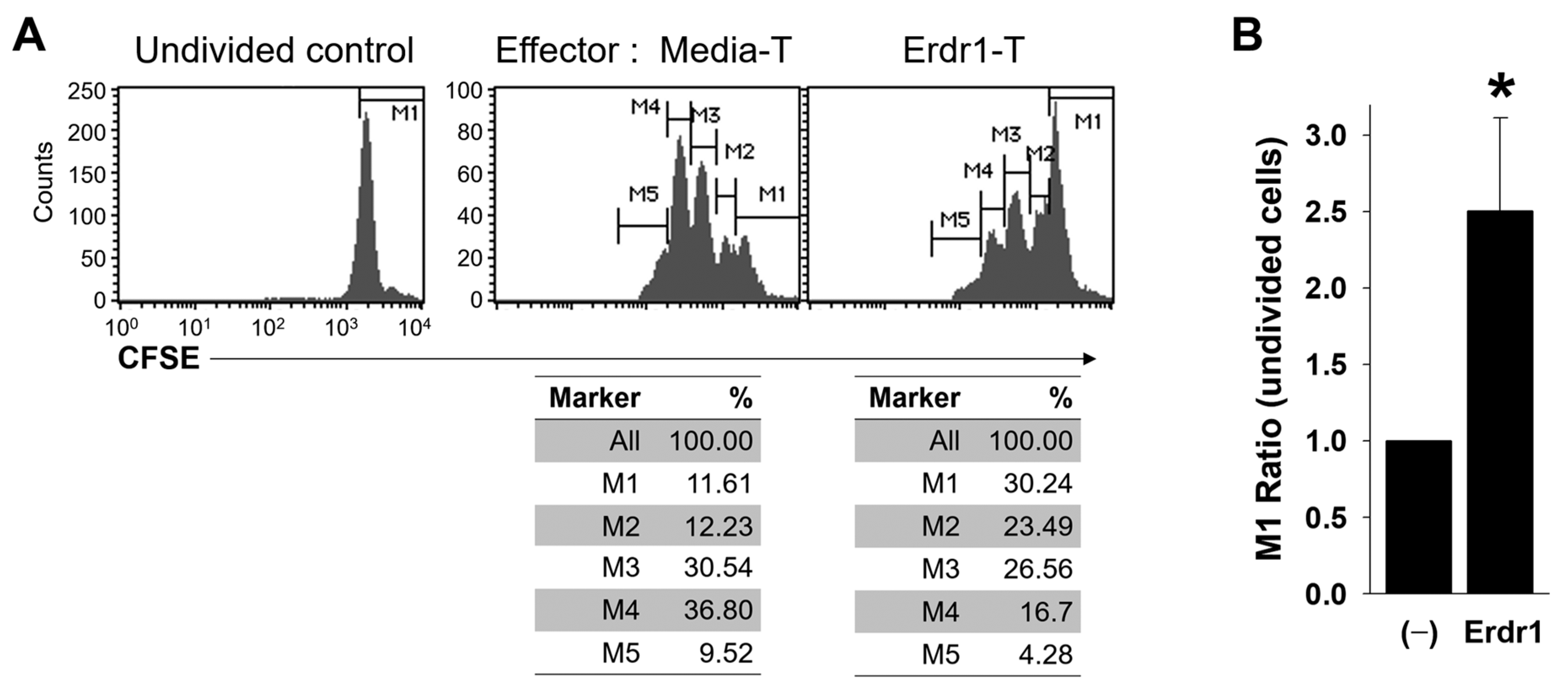
2.3. Erdr1-Activated Treg Cells are Functionally Suppressive
3. Discussion
4. Materials and Methods
4.1. Mice and Cells
4.2. Collagen-Induced Arthritis Model and Treg Depletion
4.3. Antibodies and Recombinant Erdr1
4.4. Tissue Stain
4.5. T Cell Stimulation and Suppression of T Cell Proliferation
4.6. Flow Cytometry
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CIA | Collagen-induced arthritis |
| Erdr1 | Erythroid differentiation regulator 1 |
| MFI | Mean fluorescence intensity |
| MTX | Methotrexate |
| TCR | T cell receptor |
| Treg | Regulatory T |
References
- Sakaguchi, S.; Wing, K.; Miyara, M. Regulatory T cells—a brief history and perspective. Eur. J. Immunol. 2007, 37, S116–S123. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Villar, M.; Hafler, D.A. Regulatory T cells in autoimmune disease. Nat. Immunol. 2018, 19, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Cortés, J.R.; Sánchez-Díaz, R.; Bovolenta, E.R.; Barreiro, E.; Lasarte, S.; Matesanz-Marín, R.; Toribio, M.L.; Sánchez-Madrid, F.; Martín, P. Maintenance of immune tolerance by Foxp3+ regulatory T cells requires CD69 expression. J. Autoimmun. 2014, 55, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Gu, J.; Ni, X.; Ding, Z.; Wang, Q.; Zhou, H.; Zheng, S.; Li, B.; Lu, L. CD25 signaling regulates the function and stability of peripheral Foxp3+ regulatory T cells derived from the spleen and lymph nodes of mice. Mol. Immunol. 2016, 76, 35–40. [Google Scholar] [CrossRef]
- Zhao, H.; Liao, X.; Kang, Y. Tregs: Where We Are and What Comes Next? Front. Immunol. 2017, 8, 1578. [Google Scholar] [CrossRef]
- Plitas, G.; Rudensky, A.Y. Regulatory T Cells: Differentiation and Function. Cancer Immunol. Res. 2016, 4, 721–725. [Google Scholar] [CrossRef] [Green Version]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 Control over Foxp3+ Regulatory T Cell Function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef]
- Dörmer, P.; Spitzer, E.; Möller, W. EDR is a stress-related survival factor from stroma and other tissues acting on early haematopoietic progenitors (E-Mix). Cytokine 2004, 27, 47–57. [Google Scholar] [CrossRef]
- Jung, M.K.; Park, Y.; Song, S.B.; Cheon, S.Y.; Park, S.; Houh, Y.; Ha, S.; Kim, H.J.; Park, J.M.; Kim, T.S.; et al. Erythroid Differentiation Regulator 1, an Interleukin 18-Regulated Gene, Acts as a Metastasis Suppressor in Melanoma. J. Investig. Dermatol. 2011, 131, 2096–2104. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.E.; Kim, S.; Park, S.; Houh, Y.; Yang, Y.; Park, S.B.; Kim, S.; Kim, D.; Hur, D.Y.; Kim, S.; et al. Therapeutic effect of erythroid differentiation regulator 1 (Erdr1) on collagen-induced arthritis in DBA/1J mouse. Oncotarget 2016, 7, 76354–76361. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Houh, Y.; Lee, J.; Cho, D.; Park, H.J. Downregulation of Erythroid Differentiation Regulator 1 (Erdr1) plays a critical role in psoriasis pathogenesis. Exp. Dermatol. 2016, 25, 570–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Kim, K.-E.; Jung, H.Y.; Jo, H.; Jeong, S.-W.; Lee, J.; Kim, C.H.; Kim, H.; Cho, D.; Park, H.J. Recombinant erythroid differentiation regulator 1 inhibits both inflammation and angiogenesis in a mouse model of rosacea. Exp. Dermatol. 2015, 24, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, S.; Jung, S.-J.; Park, S.; Kim, K.E.; Kim, T.S.; Park, H.J.; Cho, D. Erythroid differentiation regulator 1 strengthens TCR signaling in thymocytes by modulating calcium flux. Cell. Immunol. 2019, 336, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Casares, X.; Blanco, J.; Ambalavanan, P.; Yamanouchi, J.; Singha, S.; Fandos, C.; Tsai, S.; Wang, J.; Garabatos, N.; Izquierdo, C.; et al. Expanding antigen-specific regulatory networks to treat autoimmunity. Nature 2016, 530, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Gómez, M.; Sánchez-Madrid, F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005, 26, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lin, F.; Gao, Y.; Li, Z.; Zhang, J.; Xing, Y.; Deng, Z.; Yao, Z.; Tsun, A.; Li, B. FOXP3 and RORγt: Transcriptional regulation of Treg and Th17. Int. Immunopharmacol. 2011, 11, 536–542. [Google Scholar] [CrossRef]
- Barbi, J.; Pardoll, D.M.; Pan, F. Treg functional stability and its responsiveness to the microenvironment. Immunol. Rev. 2014, 259, 115–139. [Google Scholar] [CrossRef]
- Dayer, J.-M. Interleukin-18, rheumatoid arthritis, and tissue destruction. J. Clin. Investig. 1999, 104, 1337–1339. [Google Scholar] [CrossRef]
- Gracie, J.A.; Forsey, R.J.; Chan, W.L.; Gilmour, A.; Leung, B.P.; Greer, M.R.; Kennedy, K.; Carter, R.; Wei, X.-Q.; Xu, D.; et al. A proinflammatory role for IL-18 in rheumatoid arthritis. J. Clin. Investig. 1999, 104, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, A.; Samstein, R.M.; Treuting, P.; Liang, Y.; Pils, M.C.; Heinrich, J.-M.; Jack, R.S.; Wunderlich, F.T.; Brüning, J.C.; Müller, W.; et al. Interleukin-10 Signaling in Regulatory T Cells Is Required for Suppression of Th17 Cell-Mediated Inflammation. Immunity 2011, 34, 566–578. [Google Scholar] [CrossRef] [Green Version]
- Kelly, E.; Won, A.; Refaeli, Y.; Van Parijs, L. IL-2 and Related Cytokines Can Promote T Cell Survival by Activating AKT. J. Immunol. 2002, 168, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Soto, R.; Petersen, C.; Novis, C.L.; Kubinak, J.L.; Bell, R.; Stephens, W.Z.; Lane, T.E.; Fujinami, R.S.; Bosque, A.; O’Connell, R.M.; et al. Microbiota promotes systemic T-cell survival through suppression of an apoptotic factor. Proc. Natl. Acad. Sci. USA 2017, 114, 5497–5502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oral, H.B.; Kotenko, S.V.; Yilmaz, M.; Mani, O.; Zumkehr, J.; Blaser, K.; Akdis, C.A.; Akdis, M. Regulation of T cells and cytokines by the interleukin-10 (IL-10)-family cytokines IL-19, IL-20, IL-22, IL-24 andIL-26. Eur. J. Immunol. 2006, 36, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Tiemessen, M.M.; Kunzmann, S.; Schmidt-Weber, C.B.; Garssen, J.; Bruijnzeel-Koomen, C.A.; Knol, E.F.; van Hoffen, E. Transforming growth factor-beta inhibits human antigen-specific CD4+ T cell proliferation without modulating the cytokine response. Int. Immunol. 2003, 15, 1495–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, S.; Russo, V.; Dettori, B.; Palombi, C.; Baev, D.; Proietti, E.; Le Bon, A.; Belardelli, F.; Pace, L. Type I interferons induce peripheral T regulatory cell differentiation under tolerogenic conditions. Int. Immunol. 2020, dxaa058. [Google Scholar] [CrossRef]
- Bremser, A.; Brack, M.; Izcue, A. Higher Sensitivity of Foxp3+ Treg Compared to Foxp3- Conventional T Cells to TCR-Independent Signals for CD69 Induction. PLoS ONE 2015, 10, e0137393. [Google Scholar] [CrossRef]
- Narendra, S.C.; Chalise, J.P.; Biggs, S.; Kalinke, U.; Magnusson, M. Regulatory T-Cells Mediate IFN-α-Induced Resistance against Antigen-Induced Arthritis. Front. Immunol. 2018, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Jerala, N.K. The Role of Interferons in Inflammation and Inflammasome Activation. Front. Immunol. 2017, 8, 873. [Google Scholar] [CrossRef] [Green Version]
- Klocke, K.; Holmdahl, R.; Wing, K. CTLA-4 expressed by FOXP3+ regulatory T cells prevents inflammatory tissue attack and not T-cell priming in arthritis. Immunology 2017, 152, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Flores-Borja, F.; Jury, E.C.; Mauri, C.; Ehrenstein, M.R. Defects in CTLA-4 are associated with abnormal regulatory T cell function in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2008, 105, 19396–19401. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Shima, Y.; Wing, J.B.; Sakaguchi, S.; Ogata, A.; Kumanogoh, A. The Proportion of Regulatory T Cells in Patients with Rheumatoid Arthritis: A Meta-Analysis. PLoS ONE 2016, 11, e0162306. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.K.; Kim, K.E.; Ha, S.; Yang, Y.; Kim, D.; Kim, T.S.; Yoon, S.R.; Bang, S.I.; Cho, B.J.; Lee, W.J.; et al. Recombinant Erdr1 suppresses the migration and invasion ability of human gastric cancer cells, SNU-216, through the JNK pathway. Immunol. Lett. 2013, 150, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual Hematoxylin and Eosin Staining of Mouse Tissue Sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.S.; Lee, S.; Park, S.; Kim, K.E.; Park, H.J.; Cho, D. Erythroid Differentiation Regulator 1 Ameliorates Collagen-Induced Arthritis via Activation of Regulatory T Cells. Int. J. Mol. Sci. 2020, 21, 9555. https://doi.org/10.3390/ijms21249555
Kim MS, Lee S, Park S, Kim KE, Park HJ, Cho D. Erythroid Differentiation Regulator 1 Ameliorates Collagen-Induced Arthritis via Activation of Regulatory T Cells. International Journal of Molecular Sciences. 2020; 21(24):9555. https://doi.org/10.3390/ijms21249555
Chicago/Turabian StyleKim, Myun Soo, Sora Lee, Sunyoung Park, Kyung Eun Kim, Hyun Jeong Park, and Daeho Cho. 2020. "Erythroid Differentiation Regulator 1 Ameliorates Collagen-Induced Arthritis via Activation of Regulatory T Cells" International Journal of Molecular Sciences 21, no. 24: 9555. https://doi.org/10.3390/ijms21249555