Natural Salicylates and Their Roles in Human Health



Abstract
:1. Introduction
2. Pharmacological Roles of Natural Salicylates with Targets
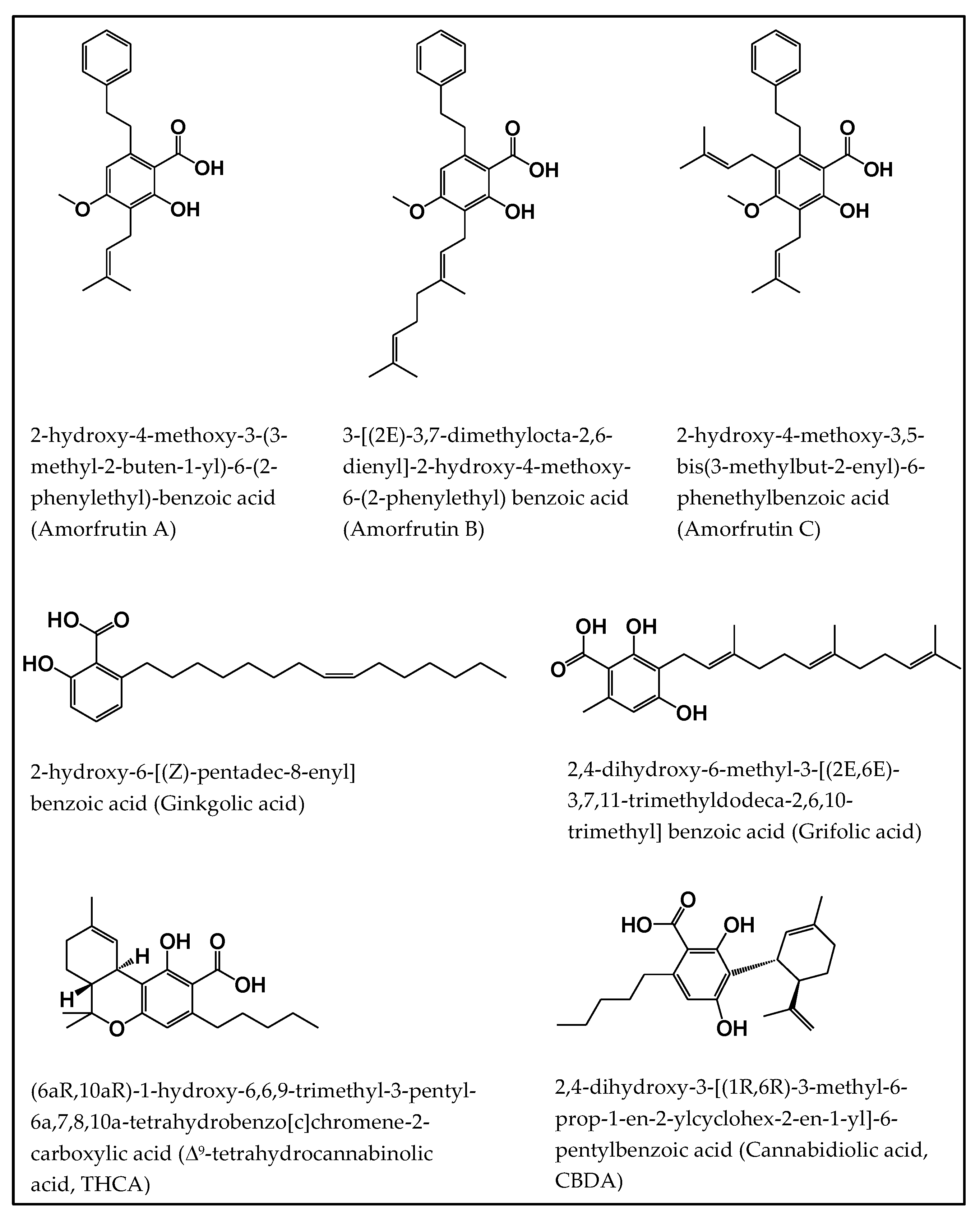
2.1. Amorfrutins
2.1.1. Antidiabetic Property
2.1.2. Anticancer Activity
2.1.3. Anti-Inflammatory Effect
2.2. Ginkgolic Acids
2.2.1. Anticancer Activity
2.2.2. Neuroprotective Activity
2.2.3. Antiviral Activity
2.3. Grifolic Acids
Antitumor Activity
2.4. Tetrahydrocannabinolic Acid (THCA) and Cannabidiolic Acid (CBDA)
2.4.1. Immunomodulatory Effect
2.4.2. Anti-Inflammatory Role
2.4.3. Neuroprotective Role
2.4.4. Anticancer Activity
2.4.5. Antiemetic Effect and Anticonvulsant Effect
3. Conclusions
Funding
Conflicts of Interest
References
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedner, T.; Everts, B. The early clinical history of salicylates in rheumatology and pain. Clin. Rheumatol. 1998, 17, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Fecka, I. Qualitative and quantitative determination of hydrolysable tannins and other polyphenols in herbal products from meadowsweet and dog rose. Phytochem. Anal. 2009, 20, 177–190. [Google Scholar] [CrossRef]
- Pino, J.A.; Mesa, J.; Muñoz, Y.; Martí, M.P.; Marbot, R. Volatile Components from Mango (Mangifera indica L.) Cultivars. J. Agric. Food Chem. 2005, 53, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Towers, G.; Tse, A.; Maass, W. Phenolic acids and phenolic glycosides of Gaultheria species. Phytochem. 1966, 5, 677–681. [Google Scholar] [CrossRef]
- Yilmaz, N.; Yayli, N.; Misir, G.; Karaoglu, S. Chemical composition and antimicrobial activities of the essential oils of Viburnum opulus, Viburnum lantana and Viburnum oriental. Asian J. Chem. 2008, 20, 3324–3330. [Google Scholar]
- Klessig, D.F.; Tian, M.; Choi, H.W. Multiple Targets of Salicylic Acid and Its Derivatives in Plants and Animals. Front. Immunol. 2016, 7, 206. [Google Scholar] [CrossRef] [Green Version]
- Anthony, J.C.; Breitner, J.C.S.; Zandi, P.P.; Meyer, M.R.; Jurasova, I.; Norton, M.C.; Stone, S.V. Reduced prevalence of AD in users of NSAIDs and H2 receptor antagonists: The Cache County Study. Neurology 2000, 54, 2066–2071. [Google Scholar] [CrossRef]
- Thun, M.J.; Jacobs, E.J.; Patrono, C. The role of aspirin in cancer prevention. Nat. Rev. Clin. Oncol. 2012, 9, 259–267. [Google Scholar] [CrossRef]
- Shirakawa, K.; Wang, L.; Man, N.; Maksimoska, J.; Sorum, A.W.; Lim, H.W.; Lee, I.S.; Shimazu, T.; Newman, J.C.; Schröder, S.; et al. Salicylate, diflunisal and their metabolites inhibit CBP/p300 and exhibit anticancer activity. eLife 2016, 5, e11156. [Google Scholar] [CrossRef]
- Kopp, E.; Ghosh, S. Inhibition of NF-κB by Sodium Salicylate and Aspirin. Science 1994, 265, 956–959. [Google Scholar] [CrossRef] [PubMed]
- Ostrowska, H. Inhibition of human platelet cathepsin A by non-steroidal anti-inflammatory drugs—in vitro study. Pol. J. Pharmacol. 1996, 48, 113–116. [Google Scholar] [PubMed]
- Stevenson, M.A.; Zhao, M.J.; Asea, A.; Coleman, C.N.; Calderwood, S.K. Salicylic Acid and Aspirin Inhibit the Activity of RSK2 Kinase and Repress RSK2-Dependent Transcription of Cyclic AMP Response Element Binding Protein- and NF-κB-Responsive Genes. J. Immunol. 1999, 163, 5608. [Google Scholar] [PubMed]
- Dachineni, R.; Ai, G.; Kumar, D.R.; Sadhu, S.S.; Tummala, H.; Bhat, G.J. Cyclin A2 and CDK2 as Novel Targets of Aspirin and Salicylic Acid: A Potential Role in Cancer Prevention. Mol. Cancer Res. 2016, 14, 241–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, O.H.; Bukhave, K.; Elmgreen, J.; Ahnfelt-Rønne, I. Inhibition of 5-lipoxygenase pathway of arachidonic acid metabolism in human neutrophils by sulfasalazine and 5-aminosalicylic acid. Dig. Dis. Sci. 1987, 32, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Tian, M.; Manohar, M.; Harraz, M.M.; Park, S.W.; Schroeder, F.C.; Snyder, S.H.; Klessig, D.F. Human GAPDH Is a Target of Aspirin’s Primary Metabolite Salicylic Acid and Its Derivatives. PLoS ONE 2015, 10, e0143447. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F.; Niederlehner, A.; Carramanzana, N.; Anton, P. Sulfasalazine inhibits the binding of TNFα to its receptor. Immunopharmacology 1990, 20, 217–224. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Saunders, M.; Barnes, P.J.; Newton, R.; Belvisi, M.G. Sodium salicylate inhibits cyclo-oxygenase-2 activity independently of transcription factor (nuclear factor kappaB) activation: Role of arachidonic acid. Mol. Pharmacol. 1997, 51, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.W.; Tian, M.; Song, F.; Venereau, E.; Preti, A.; Park, S.W.; Hamilton, K.; Swapna, G.V.T.; Manohar, M.; Moreau, M.; et al. Aspirin’s Active Metabolite Salicylic Acid Targets High Mobility Group Box 1 to Modulate Inflammatory Responses. Mol. Med. 2015, 21, 526–535. [Google Scholar] [CrossRef]
- Choi, H.W.; Wang, L.; Powell, A.F.; Strickler, S.R.; Wang, D.; Dempsey, D.A.; Schroeder, F.C.; Klessig, D.F. A genome-wide screen for human salicylic acid (SA)-binding proteins reveals targets through which SA may influence development of various diseases. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Vane, J.R. Inhibition of Prostaglandin Synthesis as a Mechanism of Action for Aspirin-like Drugs. Nat. New Biol. 1971, 231, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Akarasereenont, P.; Thiemermann, C.; Flower, R.J.; Vane, J.R. Selectivity of nonsteroidal antiinflammatory drugs as inhibitors of constitutive and inducible cyclooxygenase. Proc. Natl. Acad. Sci. USA 1993, 90, 11693–11697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.J.; Yamamoto, Y.; Gaynor, R.B. The anti-inflammatory agents aspirin and salicylate inhibit the activity of I(kappa)B kinase-beta. Nature 1998, 396, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Min, S.W.; Chen, X.; Tracy, T.E.; Li, Y.; Zhou, Y.; Wang, C.; Shirakawa, K.; Minami, S.S.; Defensor, E.; Mok, S.A.; et al. Critical role of acetylation in tau-mediated neurodegeneration and cognitive deficits. Nat. Med. 2015, 21, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Mitscher, L.A.; Park, Y.H.; AlShamma, A.; Hudson, P.; Haas, T. Amorfrutin A and B, bibenzyl antimicrobial agents from Amorpha fruticosa. Phytochemistry 1981, 20, 781–785. [Google Scholar] [CrossRef]
- Isah, T. Rethinking Ginkgo biloba L.: Medicinal uses and conservation. Pharmacogn. Rev. 2015, 9, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Fujita, T.; Kuwahara, S.; Ogura, Y. Synthesis of amorfrutins B and D from amorfrutin A ethyl ester. Tetrahedron Lett. 2020, 61, 151477. [Google Scholar] [CrossRef]
- Weidner, C.; Rousseau, M.; Micikas, R.J.; Fischer, C.; Plauth, A.; Wowro, S.J.; Siems, K.; Hetterling, G.; Kliem, M.; Schroeder, F.C.; et al. Amorfrutin C Induces Apoptosis and Inhibits Proliferation in Colon Cancer Cells through Targeting Mitochondria. J. Nat. Prod. 2016, 79, 2–12. [Google Scholar] [CrossRef]
- He, X.G.; Bernart, M.W.; Nolan, G.S.; Lin, L.Z.; Lindenmaier, M.P. High-Performance Liquid Chromatography--Electrospray Ionization-Mass Spectrometry Study of Ginkgolic Acid in the leaves and Fruits of the Ginkgo Tree (Ginkgo biloba). J. Chromatogr. Sci. 2000, 38, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.F.; Jiang, F.; Liang, X.Y.; Wei, L.-L.; Zhao, Y.Y.; Ma, Q.; Hu, Y.S.; Su, X. Grifolic acid causes osteosarcoma cell death in vitro and in tumor-bearing mice. Biomed. Pharmacother. 2018, 103, 1035–1042. [Google Scholar] [CrossRef]
- Ye, M.; Liu, J.K.; Lu, Z.X.; Zhao, Y.; Liu, S.F.; Li, L.; Tan, M.; Weng, X.X.; Li, W.; Cao, Y. Grifolin, a potential antitumor natural product from the mushroomAlbatrellus confluens, inhibits tumor cell growth by inducing apoptosis in vitro. FEBS Lett. 2005, 579, 3437–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Pang, R.P.; Shen, J.N.; Huang, G.; Wang, J.; Zhou, J.G. Grifolin induces apoptosis via inhibition of PI3K/AKT signalling pathway in human osteosarcoma cells. Apoptosis 2007, 12, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yang, L.; Xiao, L.; Xia, X.; Dong, X.; Zhong, J.; Liu, Y.; Li, N.; Chen, L.; Hongde, L.; et al. Grifolin directly targets ERK1/2 to epigenetically suppress cancer cell metastasis. Oncotarget 2015, 6, 42704–42716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, X.; Yan, H.; Sun, H.; Dongol, S.; Wang, Y.; Lv, Q.; Jiang, J. Grifolin induces autophagic cell death by inhibiting the Akt/mTOR/S6K pathway in human ovarian cancer cells. Oncol. Rep. 2016, 36, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Li, Y. Grifolin exhibits anti-cancer activity by inhibiting the development and invasion of gastric tumor cells. Oncotarget 2017, 8, 21454–21460. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.F.; Zhang, L.; Yan, A.; Chen, D.; Xie, R.; Liu, Y.; Liang, X.; Zhao, Y.; Wei, L.; Yu, J.; et al. Grifolic acid induces GH3 adenoma cell death by inhibiting ATP production through a GPR120-independent mechanism. BMC Pharmacol. Toxicol. 2018, 19, 26. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, V.; Pellati, F.; Steinbach, M.; Maran, D.; Benvenuti, S. Development of a new extraction technique and HPLC method for the analysis of non-psychoactive cannabinoids in fibre-type Cannabis sativa L. (hemp). J. Pharm. Biomed. Anal. 2017, 143, 228–236. [Google Scholar] [CrossRef]
- Pellati, F.; Brighenti, V.; Sperlea, J.; Marchetti, L.; Bertelli, D.; Benvenuti, S. New Methods for the Comprehensive Analysis of Bioactive Compounds in Cannabis sativa L. (hemp). Molecules 2018, 23, 2639. [Google Scholar] [CrossRef] [Green Version]
- Weidner, C.; De Groot, J.C.; Prasad, A.; Freiwald, A.; Quedenau, C.; Kliem, M.; Witzke, A.; Kodelja, V.; Han, C.; Giegold, S.; et al. Amorfrutins are potent antidiabetic dietary natural products. Proc. Natl. Acad. Sci. USA 2012, 109, 7257–7262. [Google Scholar] [CrossRef] [Green Version]
- Weidner, C.; Wowro, S.J.; Freiwald, A.; Kawamoto, K.; Witzke, A.; Kliem, M.; Siems, K.; Müller-Kuhrt, L.; Schroeder, F.C.; Sauer, S. Amorfrutin B is an efficient natural peroxisome proliferator-activated receptor gamma (PPARγ) agonist with potent glucose-lowering properties. Diabetology 2013, 56, 1802–1812. [Google Scholar] [CrossRef] [PubMed]
- Fuhr, L.; Rousseau, M.; Plauth, A.; Schroeder, F.C.; Sauer, S. Amorfrutins Are Natural PPARγ Agonists with Potent Anti-inflammatory Properties. J. Nat. Prod. 2015, 78, 1160–1164. [Google Scholar] [CrossRef] [PubMed]
- Meierhofer, D.; Weidner, C.; Hartmann, L.; Mayr, J.A.; Han, C.T.; Schroeder, F.C.; Sauer, S. Protein Sets Define Disease States and PredictIn VivoEffects of Drug Treatment. Mol. Cell. Proteom. 2013, 12, 1965–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Na, N.; Sheng, H.; Feng, B.; Wang, H.; Zhu, P.; Zhang, W.; Zhang, M.; Deng, Z. Ginkgolic acid inhibits the growth of renal cell carcinoma cells via inactivation of the EGFR signaling pathway. Exp. Ther. Med. 2020, 19, 2949–2956. [Google Scholar] [CrossRef] [Green Version]
- Hamdoun, S.; Efferth, T. Ginkgolic acids inhibit migration in breast cancer cells by inhibition of NEMO sumoylation and NF-κB activity. Oncotarget 2017, 8, 35103–35115. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.H.; Ko, J.H.; Lee, J.H.; Kim, C.; Lee, H.; Nam, D.; Lee, J.; Lee, S.G.; Yang, W.M.; Um, J.Y.; et al. Ginkgolic Acid Inhibits Invasion and Migration and TGF-β-Induced EMT of Lung Cancer Cells through PI3K/Akt/mTOR Inactivation. J. Cell. Physiol. 2016, 232, 346–354. [Google Scholar] [CrossRef]
- Ma, J.; Duan, W.; Han, S.; Lei, J.; Xu, Q.; Chen, X.; Jiang, Z.; Nan, L.; Li, J.; Chen, K.; et al. Ginkgolic acid suppresses the development of pancreatic cancer by inhibiting pathways driving lipogenesis. Oncotarget 2015, 6, 20993–21003. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Zheng, J.; Jin, X.; Wei, G.; Wang, G.; Sun, X.; Li, X. Ginkgolic acid inhibits the invasiveness of colon cancer cells through AMPK activation. Oncol. Lett. 2017, 14, 5831–5838. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.H.; Lee, J.H.; Kim, C.; Ko, J.H.; Ryu, S.H.; Lee, S.G.; Yang, W.M.; Um, J.Y.; Chinnathambi, A.; Alharbi, S.A.; et al. Ginkgolic Acid C 17:1, Derived from Ginkgo biloba Leaves, Suppresses Constitutive and Inducible STAT3 Activation through Induction of PTEN and SHP-1 Tyrosine Phosphatase. Molecules 2017, 22, 276. [Google Scholar] [CrossRef]
- Zhou, C.; Li, X.; Du, W.; Feng, Y.; Kong, X.; Li, Y.; Xiao, L.; Zhang, P. Antitumor Effects of Ginkgolic Acid in Human Cancer Cell Occur via Cell Cycle Arrest and Decrease the Bcl-2/Bax Ratio to Induce Apoptosis. Chemotherapy 2010, 56, 393–402. [Google Scholar] [CrossRef]
- Qi, Q.; Xue, Y.; Lv, J.; Sun, D.; Du, J.; Cai, S.; Li, Y.; Gu, T.; Wang, M. Ginkgolic acids induce HepG2 cell death via a combination of apoptosis, autophagy and the mitochondrial pathway. Oncol. Lett. 2018, 15, 6400–6408. [Google Scholar] [CrossRef] [PubMed]
- Mango, D.; Weisz, F.; Nisticò, R. Ginkgolic Acid Protects against Aβ-Induced Synaptic Dysfunction in the Hippocampus. Front. Pharmacol. 2016, 7, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, I.; Ito, A.; Hirai, G.; Nishimura, S.; Kawasaki, H.; Saitoh, H.; Kimura, K.I.; Sodeoka, M.; Yoshida, M. Ginkgolic Acid Inhibits Protein SUMOylation by Blocking Formation of the E1-SUMO Intermediate. Chem. Biol. 2009, 16, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lü, J.M.; Yan, S.; Jamaluddin, S.; Weakley, S.M.; Liang, Z.; Siwak, E.B.; Yao, Q.; Chen, C. Ginkgolic acid inhibits HIV protease activity and HIV infection in vitro. Med. Sci. Monit. 2012, 18, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borenstein, R.; Hanson, B.A.; Markosyan, R.M.; Gallo, E.S.; Narasipura, S.D.; Bhutta, M.; Shechter, O.; Lurain, N.S.; Cohen, F.S.; Al-Harthi, L.; et al. Ginkgolic acid inhibits fusion of enveloped viruses. Sci. Rep. 2020, 10, 4746. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.C.M.; Korthout, H.A.; Van Meeteren-Kreikamp, A.; Ehlert, K.A.; Wang, M.; Van Der Greef, J.; Rodenburg, R.J.T.; Witkamp, R.F. Unheated Cannabis sativa extracts and its major compound THC-acid have potential immuno-modulating properties not mediated by CB1 and CB2 receptor coupled pathways. Int. Immunopharmacol. 2006, 6, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Ruhaak, L.R.; Felth, J.; Karlsson, P.C.; Rafter, J.J.; Verpoorte, R.; Bohlin, L. Evaluation of the Cyclooxygenase Inhibiting Effects of Six Major Cannabinoids Isolated from Cannabis sativa. Biol. Pharm. Bull. 2011, 34, 774–778. [Google Scholar] [CrossRef] [Green Version]
- Nallathambi, R.; Mazuz, M.; Ion, A.; Selvaraj, G.; Weininger, S.; Fridlender, M.; Nasser, A.; Sagee, O.; Kumari, P.; Nemichenizer, D.; et al. Anti-Inflammatory Activity in Colon Models Is Derived from Δ9-Tetrahydrocannabinolic Acid That Interacts with Additional Compounds in Cannabis Extracts. Cannabis Cannabinoid Res. 2017, 2, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Moldzio, R.; Pacher, T.; Krewenka, C.; Kranner, B.; Novak, J.; Duvigneau, J.C.; Rausch, W.D. Effects of cannabinoids Δ(9)-tetrahydrocannabinol, Δ(9)-tetrahydrocannabinolic acid and cannabidiol in MPP+ affected murine mesencephalic cultures. Phytomedicine 2012, 19, 819–824. [Google Scholar] [CrossRef]
- Nadal, X.; Del Río, C.; Casano, S.; Palomares, B.; Ferreiro-Vera, C.; Navarrete, C.; Sánchez-Carnerero, C.; Cantarero, I.; Bellido, M.L.; Meyer, S.; et al. Tetrahydrocannabinolic acid is a potent PPARγ agonist with neuroprotective activity. Br. J. Pharmacol. 2017, 174, 4263–4276. [Google Scholar] [CrossRef] [Green Version]
- Ligresti, A.; Moriello, A.S.; Starowicz, K.; Matias, I.; Pisanti, S.; De Petrocellis, L.; Laezza, C.; Portella, G.; Bifulco, M.; Di Marzo, V. Antitumor Activity of Plant Cannabinoids with Emphasis on the Effect of Cannabidiol on Human Breast Carcinoma. J. Pharmacol. Exp. Ther. 2006, 318, 1375–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Petrocellis, L.; Ligresti, A.; Schiano Moriello, A.; Iappelli, M.; Verde, R.; Stott, C.G.; Cristino, L.; Orlando, P.; Di Marzo, V. Non-THC cannabinoids inhibit prostate carcinoma growth in vitro and in vivo: Pro-apoptotic effects and underlying mechanisms. Br. J. Pharmacol. 2013, 168, 79–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirao-Suzuki, M.; Takeda, S.; Koga, T.; Takiguchi, M.; Toda, A. Cannabidiolic acid dampens the expression of cyclooxygenase-2 in MDA-MB-231 breast cancer cells: Possible implication of the peroxisome proliferator-activated receptor β/δ abrogation. J. Toxicol. Sci. 2020, 45, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Okazaki, H.; Ikeda, E.; Abe, S.; Yoshioka, Y.; Watanabe, K.; Aramaki, H. Down-regulation of cyclooxygenase-2 (COX-2) by cannabidiolic acid in human breast cancer cells. J. Toxicol. Sci. 2014, 39, 711–716. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Okajima, S.; Miyoshi, H.; Yoshida, K.; Okamoto, Y.; Okada, T.; Amamoto, T.; Watanabe, K.; Omiecinski, C.J.; Aramaki, H. Cannabidiolic acid, a major cannabinoid in fiber-type cannabis, is an inhibitor of MDA-MB-231 breast cancer cell migration. Toxicol. Lett. 2012, 214, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Takeda, S.; Okazaki, H.; Watanabe, K.; Takiguchi, M.; Aramaki, H. Cannabidiolic Acid-Mediated Interference with AP-1 Transcriptional Activity in MDA-MB-231 Breast Cancer Cells. Nat. Prod. Commun. 2017, 12, 759–761. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Vellani, V.; Schiano-Moriello, A.; Marini, P.; Magherini, P.C.; Orlando, P.; Di Marzo, V. Plant-Derived Cannabinoids Modulate the Activity of Transient Receptor Potential Channels of Ankyrin Type-1 and Melastatin Type-8. J. Pharmacol. Exp. Ther. 2008, 325, 1007–1015. [Google Scholar] [CrossRef]
- Takeda, S.; Misawa, K.; Yamamoto, I.; Watanabe, K. Cannabidiolic Acid as a Selective Cyclooxygenase-2 Inhibitory Component in Cannabis. Drug Metab. Dispos. 2008, 36, 1917–1921. [Google Scholar] [CrossRef] [Green Version]
- Rock, E.M.; Limebeer, C.L.; Parker, L.A. Effect of cannabidiolic acid and ∆9-tetrahydrocannabinol on carrageenan-induced hyperalgesia and edema in a rodent model of inflammatory pain. Psychopharmacology 2018, 235, 3259–3271. [Google Scholar] [CrossRef]
- Rock, E.M.; Bolognini, D.; Limebeer, C.L.; Cascio, M.G.; Anavigoffer, S.; Fletcher, P.J.; Mechoulam, R.; Pertwee, R.G.; Parker, L.A. Cannabidiol, a non-psychotropic component of cannabis, attenuates vomiting and nausea-like behaviour via indirect agonism of 5-HT1A somatodendritic autoreceptors in the dorsal raphe nucleus. Br. J. Pharmacol. 2012, 165, 2620–2634. [Google Scholar] [CrossRef] [Green Version]
- Bolognini, D.; Rock, E.M.; Cluny, N.L.; Cascio, M.G.; Limebeer, C.L.; Duncan, M.; Stott, C.G.; Javid, F.A.; Parker, L.A.; Pertwee, R.G. Cannabidiolic acid prevents vomiting in Suncus murinus and nausea-induced behaviour in rats by enhancing 5-HT1A receptor activation. Br. J. Pharmacol. 2013, 168, 1456–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, L.L.; Low, I.K.; Banister, S.D.; McGregor, I.S.; Arnold, J.C. Pharmacokinetics of Phytocannabinoid Acids and Anticonvulsant Effect of Cannabidiolic Acid in a Mouse Model of Dravet Syndrome. J. Nat. Prod. 2019, 82, 3047–3055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehrke, M.; Lazar, M.A. The many faces of PPARgamma. Cell 2005, 123, 993–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [CrossRef] [Green Version]
- Sciacovelli, M.; Schmidt, C.; Maher, E.R.; Frezza, C. Metabolic Drivers in Hereditary Cancer Syndromes. Annu. Rev. Cancer Biol. 2020, 4, 77–97. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, M.P. Drug resistance: Challenges to effective therapy. Curr. Cancer Drug Targets 2011, 11, 613–623. [Google Scholar] [CrossRef]
- Li, J.W.H.; Vederas, J.C. Drug Discovery and Natural Products: End of an Era or an Endless Frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products As Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Ricote, M.; Glass, C.K. PPARs and molecular mechanisms of transrepression. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2007, 1771, 926–935. [Google Scholar] [CrossRef] [Green Version]
- Pascual, G.; Fong, A.L.; Ogawa, S.; Gamliel, A.; Li, A.C.; Perissi, V.; Rose, D.W.; Willson, T.M.; Rosenfeld, M.G.; Glass, C.K. A SUMOylation-dependent pathway mediates transrepression of inflammatory response genes by PPAR-gamma. Nature 2005, 437, 759–763. [Google Scholar] [CrossRef]
- Hou, Y.; Moreau, F.; Chadee, K. PPARγ is an E3 ligase that induces the degradation of NFκB/p65. Nat. Commun. 2012, 3, 1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.C.; Du, W.; Wen, Z.; Li, J.Y.; Zhang, P. Effects of natural plant ginkgolic acids on the apoptosis of human Hep-2 cancer cells. Sichuan Da Xue Xue Bao. Yi Xue Ban (J. Sichuan Univ. Med. Sci. Ed.) 2009, 40, 459–461. [Google Scholar]
- Gerstmeier, J.; Seegers, J.; Witt, F.; Waltenberger, B.; Temml, V.; Rollinger, J.; Stuppneret, H.; Koeberle, A.; Schuster, D.; Werz, O. Ginkgolic Acid is a Multi-Target Inhibitor of Key Enzymes in Pro-Inflammatory Lipid Mediator Biosynthesis. Front. Pharmacol. 2019, 10, 797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Z.; Wu, C.; Fan, G.; Tang, Z.; Cao, F. The antibacterial activity and mechanism of ginkgolic acid C15:1. BMC Biotechnol. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.; Firestone, G.L.; Bjeldanes, L.F. Bcl-2 family-mediated apoptotic effects of 3,3′-diindolylmethane (DIM) in human breast cancer cells. Biochem. Pharmacol. 2002, 63, 1085–1097. [Google Scholar] [CrossRef]
- Wesche-Soldato, D.E.; Swan, R.Z.; Chung, C.S.; Ayala, A. The apoptotic pathway as a therapeutic target in sepsis. Curr. Drug Targets 2007, 8, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Mackey, T.J.; Borkowski, A.; Amin, P.; Jacobs, S.C.; Kyprianou, N. bcl-2/bax ratio as a predictive marker for therapeutic response to radiotherapy in patients with prostate cancer. Urology 1998, 52, 1085–1090. [Google Scholar] [CrossRef]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Orfanos, C.E.; Geilen, C.C.; Wieder, T.; Sturm, I.; Daniel, P.T. The Bax/Bcl-2 Ratio Determines the Susceptibility of Human Melanoma Cells to CD95/Fas-Mediated Apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, C.M.; League, A.A.; Clark, W.S.; Lawson, D.; Derose, P.B.; Cohen, C. Bcl-2: Bax and Bcl-2: Bcl-x ratios by image cytometric quantitation of immunohistochemical expression in ovarian carcinoma: Correlation with prognosis. Cytometry 2000, 42, 61–66. [Google Scholar] [CrossRef]
- Catlett-Falcone, R.; Landowski, T.H.; Oshiro, M.M.; Turkson, J.; Levitzki, A.; Savino, R.; Ciliberto, G.; Moscinski, L.; Fernández-Luna, J.L.; Nuñez, G.; et al. Constitutive Activation of Stat3 Signaling Confers Resistance to Apoptosis in Human U266 Myeloma Cells. Immunology 1999, 10, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Bharti, A.C.; Donato, N.; Aggarwal, B.B. Curcumin (Diferuloylmethane) Inhibits Constitutive and IL-6-Inducible STAT3 Phosphorylation in Human Multiple Myeloma Cells. J. Immunol. 2003, 171, 3863–3871. [Google Scholar] [CrossRef]
- Wang, X.; Crowe, P.J.; Goldstein, D.; Yang, J.L. STAT3 inhibition, a novel approach to enhancing targeted therapy in human cancers (review). Int. J. Oncol. 2012, 41, 1181–1191. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodge, D.R.; Hurt, E.M.; Farrar, W.L. The role of IL-6 and STAT3 in inflammation and cancer. Eur. J. Cancer 2005, 41, 2502–2512. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Steinberg, B.M. PTEN is a negative regulator of STAT3 activation in human papillomavirus-infected cells. J. Gen. Virol. 2002, 83, 1651–1658. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Amin, H.M.; Franko, B.; Frantz, C.; Shi, X.; Lai, R. Loss of SHP1 enhances JAK3/STAT3 signaling and decreases proteosome degradation of JAK3 and NPM-ALK in ALK+ anaplastic large-cell lymphoma. Blood 2006, 108, 2796–2803. [Google Scholar] [CrossRef] [PubMed]
- Dorval, V.; Fraser, P.E. SUMO on the road to neurodegeneration. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1773, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, P.M.; Hayashi, K.M.; de Zubicaray, G.; Janke, A.L.; Rose, S.E.; Semple, J.; Herman, D.; Hong, M.S.; Dittmer, S.S.; Doddrell, D.M.; et al. Dynamics of gray matter loss in Alzheimer’s disease. J. Neurosci. 2003, 23, 994–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendt, T. Synaptic degeneration in Alzheimer’s disease. Acta Neuropathol. 2009, 118, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Smith, M.; Holodniy, M.; O Kahn, J. HIV-1 protease inhibitors. A review for clinicians. JAMA 1997, 277, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Formato, M.; Crescente, G.; Scognamiglio, M.; Fiorentino, A.; Pecoraro, M.T.; Piccolella, S.; Catauro, M.; Pacifico, S. (–)-Cannabidiolic Acid, a Still Overlooked Bioactive Compound: An Introductory Review and Preliminary Research. Molecules 2020, 25, 2638. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.W.; Lane, B.; Newton, C.A.; Friedman, H. The cannabinoid system and cytokine network. Proc. Soc. Exp. Biol. Med. 2000, 225, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bidaut-Russell, M.; DeVane, W.A.; Howlett, A.C. Cannabinoid Receptors and Modulation of Cyclic AMP Accumulation in the Rat Brain. J. Neurochem. 1990, 55, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Herring, A.C.; Kaminski, N.E. Cannabinol-mediated inhibition of nuclear factor-kappaB, cAMP response element-binding protein, and interleukin-2 secretion by activated thymocytes. J. Pharmacol. Exp. Ther. 1999, 291, 1156–1163. [Google Scholar]
- Bonhaus, D.W.; Chang, L.K.; Kwan, J.; Martin, G.R. Dual activation and inhibition of adenylyl cyclase by cannabinoid receptor agonists: Evidence for agonist-specific trafficking of intracellular responses. J. Pharmacol. Exp. Ther. 1998, 287, 884–888. [Google Scholar] [PubMed]
- Klein, T.W.; Newton, C.A.; Friedman, H. Cannabinoids and the immune system. Pain Res. Manag. 2001, 6, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Grotenhermen, F. Pharmacokinetics and Pharmacodynamics of Cannabinoids. Clin. Pharmacokinet. 2003, 42, 327–360. [Google Scholar] [CrossRef]
- Wiley, J.L.; Martin, B.R. Cannabinoid pharmacology: Implications for additional cannabinoid receptor subtypes. Chem. Phys. Lipids 2002, 121, 57–63. [Google Scholar] [CrossRef]
- Stone, N.L.; Murphy, A.J.; England, T.J.; O’Sullivan, S.E. A systematic review of minor phytocannabinoids with promising neuroprotective potential. Br. J. Pharmacol. 2020, 177, 4330–4352. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural Products for the Treatment of Type 2 Diabetes Mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, C.S.; Conroy, M.; Heuvel, B.D.V.; Park, S.H. Cannabidiol Drugs Clinical Trial Outcomes and Adverse Effects. Front. Pharmacol. 2020, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.; Robertson, A.A.B.; Cooper, M.A. Natural product and natural product derived drugs in clinical trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Salicylates & Structure | Proteins | References |
|---|---|---|
| Salicylic acid | Ferrochelatase (FECH) | [16] |
 | Cyclin-dependent kinase 2 (CDK2) | [14] |
| α-Enolase (ENO1) | [20] | |
| Pyruvate kinase isozyme M2 (PKM2) | [20] | |
| Acetylsalicylic acid (Aspirin) | Cyclooxygenase-1 (COX-1) | [21,22] |
 | Cyclooxygenase-2 (COX-2) | [18,21,22] |
| Nuclear factor-kappaB (NF-κB) | [11] | |
| Cathepsin A (CTSA) | [12] | |
| Inhibitor of nuclear factorkappa-B kinase subunit beta (Iκκ-β) | [23] | |
| Ribosomal S6 kinase 2 (RSK2) | [13] | |
| Cyclin-dependent kinase 2 (CDK2) | [14] | |
| 5-aminosalicylic acid (Mesalazine) | Arachidonate 5-lipoxygenase (ALOX5) | [15] |
 | Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) | [16] |
| 2-(2-hydroxybenzoyl) oxybenzoic acid (Salsalate) | Acetyltransferase p300 (P300) | [24] |
 | ||
| Acetyl 3-aminoethyl salicylic acid (Ac3AESA) | Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) | [16] |
 | High mobility group box 1 (HMGB1) | [19] |
| 2-hydroxy-5-[(E)-2-{4-[(pyridin-2-yl) sulfamoyl] phenyl} diazen-1-yl] benzoic acid (Sulfasalazine) | Tumor necrosis factor alpha (TNFα) | [17] |
 | Arachidonate 5-lipoxygenase (ALOX5) | [15] |
| Sodium salicylate | Cyclooxygenase-2 (COX-2) | [18,21,22] |
 | Nuclear factor-kappaB(NF-κB) | [11] |
| Inhibitor of nuclear factorkappa-B kinase subunit beta (Iκκ-β) | [23] |
| Natural Salicylates | Source | Biological Activity | Mechanism of Action | Ref |
|---|---|---|---|---|
| Amorfrutins | A. fruticosa L. (bastard indigo), G. foetida Desf. (licorice) | Antidiabetic | Inhibition of insulin resistance formation, dyslipidemia, liver steatosis, and induction of PPARγ ser273 phosphorylation by HFD | [40,41,43] |
| Anticancer | Formation of reactive oxygen species, DNA fragmentation, caspase activation, phosphatidylserine externalization, and PARP cleavage induced apoptosis in colorectal cancer cells | [28] | ||
| Anti-inflammatory | Lowering the expression of various inflammatory genes such as COX-2, GRO-α, IL-8, and MIP-3α in TNF-α- stimulated colon cells | [42] | ||
| Ginkgolic acids | G. biloba L. | Anticancer | Inhibition of the proliferation of renal cell carcinoma (RCC) cell lines 786-O and A498 by inactivating epidermal growth factor receptor (EGFR) signaling pathway with the downregulation of p-Akt and p-Erk expression | [44] |
| Inhibition of IκBα degradation and reduction of the activity of NF-κB in MCF-7 and MDA-MB 231 breast cancer cells | [45] | |||
| Inhibition of the PI3K/Akt/mTOR signaling pathway of lung cancer cells | [46] | |||
| Reduction of the cell viability and prevention of the de novo lipogenesis of pancreatic cancer cells | [47] | |||
| Reduction of proliferation, migration, and invasion of SW480 colon cancer cells. Stimulation of AMPK activation and decreasing the expression of invasion-associated proteins, such as (MMP)-2, (CXCR4) and uPA | [48] | |||
| Suppression of STAT3 phosphorylation and reduction of cell proliferation in multiple myeloma cancer cells. Upregulation of the expression of PTEN and SHP-1 | [49] | |||
| Inhibition of the growth of human tongue squamous carcinoma cells. Reduction of the Bcl-2/Bax ratio and stimulation of caspase-3 activity | [50] | |||
| Inhibition the cell proliferation and induction of cell death through a combination of apoptosis, autophagy, and the mitochondrial pathway in human hepatoblastoma HepG2 cells | [51] | |||
| Neuroprotective | Enhancement of long-term potentiation (LTP) in the hippocampus, restoring the Aβ-mediated paired-pulse ratio (PPR) alteration and rescuing the Aβ-mediated change in excitatory neurotransmission in mice model. | [52] | ||
| Inhibition of the formation of E1-SUMO intermediate in an in situ cell-based SUMOylation assay | [53] | |||
| Antiviral | Inhibition of HIV protease activity of HIV-1SF162 infection in human peripheral blood mononuclear cells | [54] | ||
| The inhibitory effect on the fusion of a variety of enveloped viruses, including ZIKV, HSV-1, HCMV HIV, EBOV, IAV and EBV, and a nonenveloped human adenovirus | [55] | |||
| Grifolic acid | A. confluens | Antitumor | Reduction of cell viability by blocking NADH production and inhibiting MMP and ATP production on both GH3 cells and osteosarcoma cell | [30,36] |
| Tetrahydro-cannabinolic acid (THCA) | C. sativa | Immunomodulatory | Targeting PC-PLC enzymatic activity in phospholipids metabolism and inhibition of tumor TNF-a release from LPS-activated U937 macrophages and peripheral blood macrophages | [56] |
| Anti-inflammatory | Inhibition COX1 and COX2 expression, Reduction of prostaglandin production also inhibition MMP9 expression in colon cancer cell | [57,58] | ||
| Neuroprotective | Reduction of dopaminergic neurons degenerative effect of MPP+, increasing cell survival in mice mesencephalic cultures | [59] | ||
| Reduction of degenerative effects of 3-NPA, through a PPARγ-dependent pathway in N2a cell and in vivo mice model | [60] | |||
| Antineoplastic | Inhibition of cell proliferation in the breast cancer cell and prostate carcinoma cell | [61,62] | ||
| Cannabidiolic acid (CBDA) | C. sativa | Anticancer | Inhibition of the transcriptional activities of both activator protein I (AP-I) and peroxisome proliferator-activated receptor PPARβ/δ in breast cancer cell | [63] |
| Inhibition of human breast cancer cell metastasis by suppressing COX2 and proto-oncogene c-Fos expression and upregulation of the expression of SHARP1 | [64,65,66] | |||
| Anti-inflammatory | Stimulation of vanilloid 1 and ankyrin 1 transient receptor potential (TRP) channels (TRPV1 and TRPA1, respectively), and antagonize a receptor, the Transient Receptor Potential Cation Channel Subfamily M Member 8 (TRPM8) in HEK 293 cells | [67] | ||
| Selective inhibition of COX2. Reduction of inflammation when intraperitoneally administered in a rodent model of carrageenan-induced acute inflammation in the rat hind paw. | [68,69] | |||
| Antiemetic | Induction 5-HT1A receptors activity in vivo in mice | [70,71] | ||
| Anticonvulsant | Increasing the temperature threshold at which the Scn1aRX/+ mice had a generalized tonic-clonic seizure. | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeasmin, F.; Choi, H.W. Natural Salicylates and Their Roles in Human Health. Int. J. Mol. Sci. 2020, 21, 9049. https://doi.org/10.3390/ijms21239049
Yeasmin F, Choi HW. Natural Salicylates and Their Roles in Human Health. International Journal of Molecular Sciences. 2020; 21(23):9049. https://doi.org/10.3390/ijms21239049
Chicago/Turabian StyleYeasmin, Fatema, and Hyong Woo Choi. 2020. "Natural Salicylates and Their Roles in Human Health" International Journal of Molecular Sciences 21, no. 23: 9049. https://doi.org/10.3390/ijms21239049