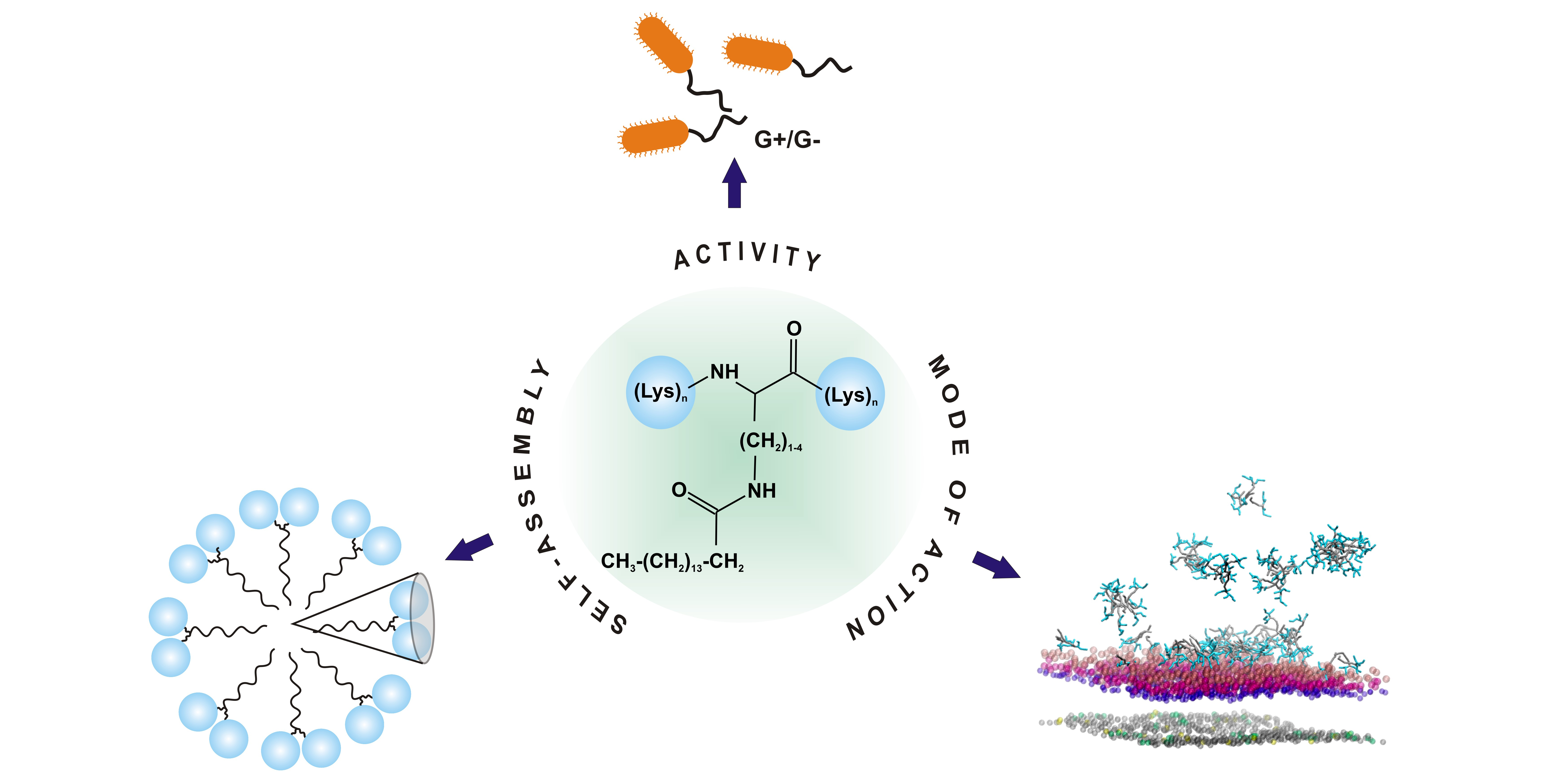
Double-Headed Cationic Lipopeptides: An Emerging Class of Antimicrobials

 , , ,
, , ,  , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Antimicrobial and Toxic Activities
2.2. Bacterial Viability
2.3. Self-Assembly Properties
2.4. Effect of Double-Headed Lipopeptides on the Lipid Hydrocarbon Chain Conformational State
2.5. Isothermal Titration Calorimetry (ITC) Studies of Lipopeptide-Membrane Interactions
2.6. Coarse-Grained Molecular Dynamic Simulations of Lipopeptide-Membrane Interactions
3. Materials and Methods
3.1. Reagents
3.2. Chemical Synthesis of Lipid-Peptide Conjugates
3.3. Antimicrobial Activity Assays
3.4. Hemolytic Activity
3.5. Cytotoxicity Towards Human Cells
3.6. Fluorescence Microscopy
3.7. Surface Tension Measurements
3.8. NMR Measurements
3.9. Liposome Preparation
3.10. Isothermal Titration Calorimetry Measurements
3.11. Fourier Transform Infrared (FTIR) Measurements
3.12. Molecular Dynamic Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CAC | Critical aggregation concentration |
| C16 | Palmitic acid |
| CDL2 | Cardiolipin 2 |
| CG MD | Coarse-grained molecular dynamics |
| DPPC | 1,2-Dipalmitoyl-sn-glycero-3-phosphocholine |
| DPPG | 1,2-Dipalmitoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] |
| LUV | Large unilamellar vesicle |
| MLV | Multilayer vesicle |
| POPC | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine |
| POPE | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylethanolamine |
| POPG | 1-Palmitoyl-2-oleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] |
| RaLPS | Ra mutant rough chemotype lipopolysaccharide |
References
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Menacho-Melgar, R.; Decker, J.S.; Hennigan, J.N.; Lynch, M.D. A review of lipidation in the development of advanced protein and peptide therapeutics. J. Control. Release 2018, 295, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, R.; Harris, P.W.; Williams, G.M.; Yang, S.-H.; Brimble, M.A. Peptide Lipidation—A Synthetic Strategy to Afford Peptide Based Therapeutics. In Peptides and Peptide-based Biomaterials and Their Biomedical Applications; Springer: Cham, Switzerland, 2017; pp. 185–227. [Google Scholar]
- Chu-Kung, A.F.; Nguyen, R.; Bozzelli, K.N.; Tirrell, M. Chain length dependence of antimicrobial peptide-fatty acid conjugate activity. J. Colloid Interface Sci. 2010, 345, 160–167. [Google Scholar] [CrossRef]
- Vaezi, Z.; Bortolotti, A.; Luca, V.; Perilli, G.; Mangoni, M.L.; Khosravi-Far, R.; Bobone, S.; Stella, L. Aggregation determines the selectivity of membrane-active anticancer and antimicrobial peptides: The case of killerFLIP. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183107. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Grossfield, A. Thermodynamics of micelle formation and membrane fusion modulate antimicrobial lipopeptide activity. Biophys. J. 2015, 109, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Sikorska, E.; Stachurski, O.; Neubauer, D.; Małuch, I.; Wyrzykowski, D.; Bauer, M.; Brzozowski, K.; Kamysz, W. Short arginine-rich lipopeptides: From self-assembly to antimicrobial activity. Biochim. Biophys. Acta Biomembr. 2018, 1860, 2242–2251. [Google Scholar] [CrossRef]
- Dawgul, M.; Greber, K.; Bartoszewska, S.; Baranska-Rybak, W.; Sawicki, W.; Kamysz, W. In vitro evaluation of cytotoxicity and permeation study on lysine-and arginine-based lipopeptides with proven antimicrobial activity. Molecules 2017, 22, 2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domalaon, R.; Brizuela, M.; Eisner, B.; Findlay, B.; Zhanel, G.G.; Schweizer, F. Dilipid ultrashort cationic lipopeptides as adjuvants for chloramphenicol and other conventional antibiotics against Gram-negative bacteria. Amino Acids 2019, 51, 383–393. [Google Scholar] [CrossRef]
- Azmi, F.; Elliott, A.G.; Marasini, N.; Ramu, S.; Ziora, Z.; Kavanagh, A.M.; Blaskovich, M.A.; Cooper, M.A.; Skwarczynski, M.; Toth, I. Short cationic lipopeptides as effective antibacterial agents: Design, physicochemical properties and biological evaluation. Bioorg. Med. Chem. 2016, 24, 2235–2241. [Google Scholar] [CrossRef]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Kamysz, W.; Silvestri, C.; Orlando, F.; Mocchegiani, F.; Vittoria, A.D.; Kamysz, E.; Saba, V.; et al. The lipopeptides Pal-Lys-Lys-NH(2) and Pal-Lys-Lys soaking alone and in combination with intraperitoneal vancomycin prevent vascular graft biofilm in a subcutaneous rat pouch model of staphylococcal infection. Peptides 2007, 28, 1299–1303. [Google Scholar] [CrossRef] [PubMed]
- Paduszynska, M.A.; Maciejewska, M.; Neubauer, D.; Golacki, K.; Szymukowicz, M.; Bauer, M.; Kamysz, W. Influence of Short Cationic Lipopeptides with Fatty Acids of Different Chain Lengths on Bacterial Biofilms Formed on Polystyrene and Hydrogel Surfaces. Pharmaceutics 2019, 11, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachurski, O.; Neubauer, D.; Małuch, I.; Wyrzykowski, D.; Bauer, M.; Bartoszewska, S.; Kamysz, W.; Sikorska, E. Effect of self-assembly on antimicrobial activity of double-chain short cationic lipopeptides. Bioorg. Med. Chem. 2019, 27, 115129. [Google Scholar] [CrossRef] [PubMed]
- Barchiesi, F.; Giacometti, A.; Cirioni, O.; Arzeni, D.; Silvestri, C.; Kamysz, W.; Abbruzzetti, A.; Riva, A.; Kamysz, E.; Scalise, G. In vitro activity of the synthetic lipopeptide Pal-Lys-Lys-NH2 alone and in combination with antifungal agents against clinical isolates of Cryptococcus neoformans. Peptides 2007, 28, 1509–1513. [Google Scholar] [CrossRef] [PubMed]
- Kamysz, W.; Silvestri, C.; Cirioni, O.; Giacometti, A.; Licci, A.; Della Vittoria, A.; Okroj, M.; Scalise, G. In vitro activities of the lipopeptides palmitoyl (Pal)-Lys-Lys-NH2 and Pal-Lys-Lys alone and in combination with antimicrobial agents against multiresistant Gram-positive cocci. Antimicrob. Agents Chemother. 2007, 51, 354–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greber, K.E.; Dawgul, M.; Kamysz, W.; Sawicki, W. Cationic net charge and counter ion type as antimicrobial activity determinant factors of short lipopeptides. Front. Microbiol. 2017, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Findlay, B.; Zhanel, G.G.; Schweizer, F. Investigating the antimicrobial peptide ‘window of activity’using cationic lipopeptides with hydrocarbon and fluorinated tails. Int. J. Antimicrob. Agents 2012, 40, 36–42. [Google Scholar] [CrossRef]
- Greber, K.E.; Dawgul, M.; Kamysz, W.; Sawicki, W.; Łukasiak, J. Biological and surface-active properties of double-chain cationic amino acid-based surfactants. Amino Acids 2014, 46, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Haldar, J.; Kondaiah, P.; Bhattacharya, S. Synthesis and antibacterial properties of novel hydrolyzable cationic amphiphiles. Incorporation of multiple head groups leads to impressive antibacterial activity. J. Med. Chem. 2005, 48, 3823–3831. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Samanta, S.K. Surfactants possessing multiple polar heads. A perspective on their unique aggregation behavior and applications. J. Phys. Chem. Lett. 2011, 2, 914–920. [Google Scholar] [CrossRef]
- Greber, K.E. Synthesis and Surface Activity of Cationic Amino Acid-Based Surfactants in Aqueous Solution. J. Surfactants Deterg. 2017, 20, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groves, P. Diffusion ordered spectroscopy (DOSY) as applied to polymers. Polym. Chem. 2017, 8, 6700–6708. [Google Scholar] [CrossRef]
- Schwieger, C.; Blume, A. Interaction of poly (L-arginine) with negatively charged DPPG membranes: Calorimetric and monolayer studies. Biomacromolecules 2009, 10, 2152–2161. [Google Scholar] [CrossRef] [PubMed]
- Schwieger, C.; Blume, A. Interaction of poly(L-lysines) with negatively charged membranes: An FT-IR and DSC study. Eur. Biophys. J. Biophy. 2007, 36, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, J.R.; Andresen, T.L. Thermodynamic profiling of peptide membrane interactions by isothermal titration calorimetry: A search for pores and micelles. Biophys. J. 2011, 101, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenk, M.R.; Seelig, J. Magainin 2 amide interaction with lipid membranes: Calorimetric detection of peptide binding and pore formation. Biochemistry 1998, 37, 3909–3916. [Google Scholar] [CrossRef]
- Wenk, M.R.; Seelig, J. Vesicle—Micelle Transformation of Phosphatidylcholine/Octyl-β-d-glucopyranoside Mixtures as Detected with Titration Calorimetry. J. Phys. Chem. 1997, 101, 5224–5231. [Google Scholar] [CrossRef]
- Hoernke, M.; Schwieger, C.; Kerth, A.; Blume, A. Binding of cationic pentapeptides with modified side chain lengths to negatively charged lipid membranes: Complex interplay of electrostatic and hydrophobic interactions. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1663–1672. [Google Scholar] [CrossRef] [Green Version]
- Sikorska, E.; Dawgul, M.; Greber, K.; Iłowska, E.; Pogorzelska, A.; Kamysz, W. Self-assembly and interactions of short antimicrobial cationic lipopeptides with membrane lipids: ITC, FTIR and molecular dynamics studies. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2625–2634. [Google Scholar] [CrossRef] [Green Version]
- Abraham, T.; Lewis, R.N.; Hodges, R.S.; McElhaney, R.N. Isothermal titration calorimetry studies of the binding of the antimicrobial peptide gramicidin S to phospholipid bilayer membranes. Biochemistry 2005, 44, 11279–11285. [Google Scholar] [CrossRef]
- Simon, S.A.; McIntosh, T.J. Peptide-Lipid Interactions, 1st ed.; Academic Press: Cambridge, MA, USA, 2002; Volume 52. [Google Scholar]
- Sinha, S.; Zheng, L.; Mu, Y.; Ng, W.J.; Bhattacharjya, S. Structure and Interactions of A Host Defense Antimicrobial Peptide Thanatin in Lipopolysaccharide Micelles Reveal Mechanism of Bacterial Cell Agglutination. Sci. Rep. 2017, 7, 17795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandenburg, K.; Garidel, P.; Fukuoka, S.; Howe, J.; Koch, M.H.; Gutsmann, T.; Andrä, J. Molecular basis for endotoxin neutralization by amphipathic peptides derived from the alpha-helical cationic core-region of NK-lysin. Biophys. Chem. 2010, 150, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Mohanram, H.; Bhattacharjya, S. Resurrecting inactive antimicrobial peptides from the lipopolysaccharide trap. Antimicrob. Agents. Chemother. 2014, 58, 1987–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshminarayanan, R.; Tan, W.X.; Aung, T.T.; Goh, E.T.; Muruganantham, N.; Li, J.; Chang, J.Y.; Dikshit, N.; Saraswathi, P.; Lim, R.R.; et al. Branched Peptide, B2088, Disrupts the Supramolecular Organization of Lipopolysaccharides and Sensitizes the Gram-negative Bacteria. Sci. Rep. 2016, 6, 25905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts: Approved Standards, 2nd ed.; CLSI Document M27-2A 2002; CLSI: Wayne, PA, USA, 2002. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests f or Bacteria That Grow Aerobically; Approved Standard, 9th ed.; CLSI document M07-A9; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Stejskal, E.; Tanner, J. Spin diffusion measurements: Spin echoes in the presence of a time-dependent field gradient. J. Chem. Phys. 1965, 42, 288–292. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Kutzner, C.; Van Der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Periole, X.; Marrink, S.J. The Martini coarse-grained force field. Methods Mol. Biol. 2013, 924, 533–565. [Google Scholar]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; de Vries, A.H. The MARTINI force field: Coarse grained model for biomolecular simulations. J. Phys. Chem. B 2007, 111, 7812–7824. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L.; Mackerell, A.D.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Dávila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M. CHARMM-GUI membrane builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chugunov, A.; Pyrkova, D.; Nolde, D.; Polyansky, A.; Pentkovsky, V.; Efremov, R. Lipid-II forms potential “landing terrain” for lantibiotics in simulated bacterial membrane. Sci. Rep. 2013, 3, 1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.C.; Bruininks, B.M.H.; Jefferies, D.; Cesar Telles de Souza, P.; Lee, J.; Patel, D.S.; Marrink, S.J.; Qi, Y.; Khalid, S.; Im, W. CHARMM-GUI Martini Maker for modeling and simulation of complex bacterial membranes with lipopolysaccharides. J. Comput. Chem. 2017, 38, 2354–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winger, M.; Trzesniak, D.; Baron, R.; van Gunsteren, W.F. On using a too large integration time step in molecular dynamics simulations of coarse-grained molecular models. Phys. Chem. Chem. Phys. 2009, 11, 1934–1941. [Google Scholar] [CrossRef] [Green Version]
- Allen, W.J.; Lemkul, J.A.; Bevan, D.R. GridMAT-MD: A grid-based membrane analysis tool for use with molecular dynamics. J. Comput. Chem. 2009, 30, 1952–1958. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | MIC [µg/mL] | ||||
|---|---|---|---|---|---|
| Gram-Positive Bacteria | Gram-Negative Bacteria | Fungus | |||
| S. aureus | S. epidermidis | P. aeruginosa | E. coli | C. albicans | |
| KK(KC16)KK-NH2 | 8 | 8 | 16 | 4 | 16 |
| KK(OC16)KK-NH2 | 8 | 4 | 32 | 8 | 32 |
| KK(DabC16)KK-NH2 | 4 | 8 | 32 | 8 | 32 |
| KK(DapC16)KK-NH2 | 4 | 8 | 32 | 8 | 256 |
| K(KC16)K-NH2 | 4 | 8 | 16 | 8 | 8 |
| K(OC16)K-NH2 | 8 | 8 | 8 | 8 | 8 |
| K(DabC16)K-NH2 | 8 | 8 | 16 | 8 | 16 |
| K(DapC16)K-NH2 | 8 | 4 | 8 | 8 | 16 |
| Peptide | IC50 [µg/mL] | |||||||
|---|---|---|---|---|---|---|---|---|
| 46BR.1N (24h) | HaCaT (24h) | A375 (48h) | A549 (48h) | HCT-116 (48h) | HT-29 (48h) | LNCaP (48h) | MCF-7 (48h) | |
| KK(KC16)KK-NH2 | 7.56 ± 1.31 | 11.92 ± 1.57 | >25 | >25 | 15.98 ± 0.16 | 18.98 ± 0.38 | >25 | >25 |
| KK(OC16)KK-NH2 | 8.21 ± 1.12 | 16.65 ± 2.29 | 14.90 ± 0.75 | 19.36 ± 0.58 | ||||
| KK(DabC16)KK-NH2 | 9.45 ± 2.13 | 16.95 ± 1.43 | 16.74 ± 0.33 | 20.75 ± 0.42 | ||||
| KK(DapC16)KK-NH2 | 12.77 ± 1.32 | 49.31 ± 16.6 | 23.05 ± 0.92 | 24.68 ± 0.49 | ||||
| K(KC16)K-NH2 | 9.36 ± 2.31 | 9.08 ± 2.28 | 20.34 ± 1.47 | 20.72 ± 0.62 | 21.97 ± 1.10 | 22.71 ± 0.79 | ||
| K(OC16)K-NH2 | 9.02 ± 0.69 | 6.25 ± 3.08 | 21.60 ± 0.76 | 18.07 ± 0.54 | 20.51 ± 0.41 | 15.47 ± 1.04 | ||
| K(DabC16)K-NH2 | 9.76 ± 0.70 | 7.96 ± 2.49 | 11.99 ± 0.38 | 16.79 ± 0.34 | 21.56 ± 0.22 | 14.3 ± 0.45 | 16.28 ± 1.02 | |
| K(DapC16)K-NH2 | 8.68 ± 1.00 | 7.25 ± 2.18 | 11.63 ± 0.26 | 14.90 ± 0.30 | 20.24 ± 0.81 | 14.0 ± 0.41 | 20.11 ± 2.51 | |
| Peptide | CAC | γCAC | |
|---|---|---|---|
| mM | mg/mL | mN/m | |
| K(KC16)K-NH2 | 5.4 | 3.5 | 43.0 |
| K(OC16)K-NH2 | 5.9 | 3.7 | 39.2 |
| K(DabC16)K-NH2 | 6.6 | 4.0 | 39.8 |
| K(DapC16)K-NH2 | 7.2 | 4.3 | 38.5 |
| Peptide | n | pKITC | ΔH° kcal mol−1 | TΔS° kcal mol−1 | ΔG° kcal mol−1 |
|---|---|---|---|---|---|
| POPG | |||||
| KK(DapC16)KK-NH2 | 3.46 ± 0.06 | 5.66 ± 0.12 | −0.68 ± 0.01 | 9.41 ± 0.17 | −10.09 ± 0.17 |
| K(KC16)K-NH2 | 1.13 ± 0.03 | 5.66 ± 0.11 | −1.24 ± 0.04 | 8.85 ± 0.20 | −10.10 ± 0.15 |
| K(OC16)K-NH2 | 1.59 ± 0.03 | 5.79 ± 0.12 | −1.35 ± 0.03 | 8.92 ± 0.16 | −10.27 ± 0.16 |
| K(DabC16)K-NH2 | 0.73 ± 0.06 | 8.09 ± 0.37 | −1.22 ± 0.06 | 9.41 ± 0.17 | −10.09 ± 0.17 |
| 1.57 ± 0.07 | 6.59 ± 0.26 | −0.68 ± 0.04 | 10.68 ± 0.36 | −11.36 ± 0.36 | |
| K(DapC16)K-NH2 | 0.43 ± 0.03 | 8.93 ± 0.53 | −1.20 ± 0.03 | 13.34 ± 0.72 | −14.55 ± 0.72 |
| 0.94 ± 0.03 | 6.64 ± 0.25 | −0.73 ± 0.03 | 10.69 ± 0.34 | −11.42 ± 0.34 | |
| LPS E. coli 055:B5 | |||||
| KK(KC16)KK-NH2 | 0.59 ± 0.04 | 7.67 ± 0.29 | −1.96 ± 0.50 | 10.987 ± 0.64 | −12.83 ± 0.40 |
| 0.67 ± 0.06 | 6.38 ± 0.14 | −13.16 ± 1.42 | −2.09 ± 1.43 | −11.07 ± 0.19 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Małuch, I.; Stachurski, O.; Kosikowska-Adamus, P.; Makowska, M.; Bauer, M.; Wyrzykowski, D.; Hać, A.; Kamysz, W.; Deptuła, M.; Pikuła, M.; et al. Double-Headed Cationic Lipopeptides: An Emerging Class of Antimicrobials. Int. J. Mol. Sci. 2020, 21, 8944. https://doi.org/10.3390/ijms21238944
Małuch I, Stachurski O, Kosikowska-Adamus P, Makowska M, Bauer M, Wyrzykowski D, Hać A, Kamysz W, Deptuła M, Pikuła M, et al. Double-Headed Cationic Lipopeptides: An Emerging Class of Antimicrobials. International Journal of Molecular Sciences. 2020; 21(23):8944. https://doi.org/10.3390/ijms21238944
Chicago/Turabian StyleMałuch, Izabela, Oktawian Stachurski, Paulina Kosikowska-Adamus, Marta Makowska, Marta Bauer, Dariusz Wyrzykowski, Aleksandra Hać, Wojciech Kamysz, Milena Deptuła, Michał Pikuła, and et al. 2020. "Double-Headed Cationic Lipopeptides: An Emerging Class of Antimicrobials" International Journal of Molecular Sciences 21, no. 23: 8944. https://doi.org/10.3390/ijms21238944