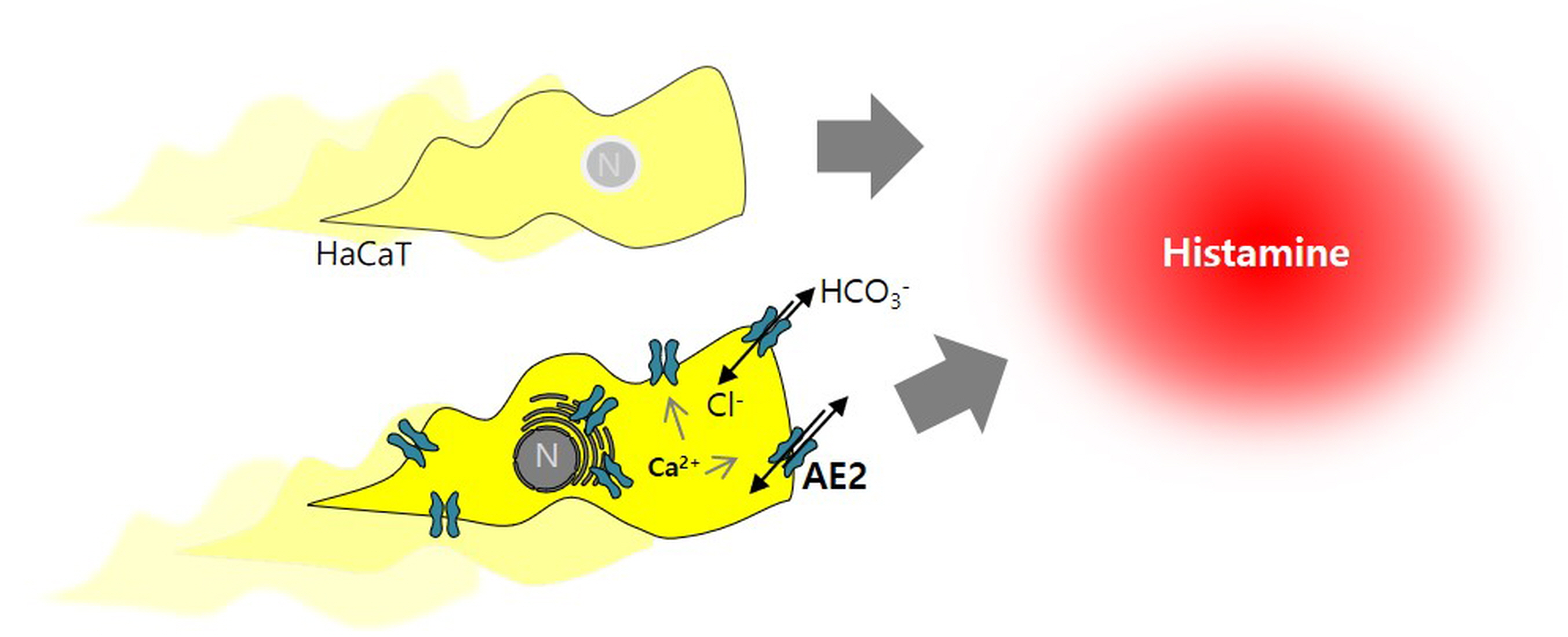
Intracellular Ca2+-Mediated AE2 Is Involved in the Vectorial Movement of HaCaT Keratinocyte


Abstract
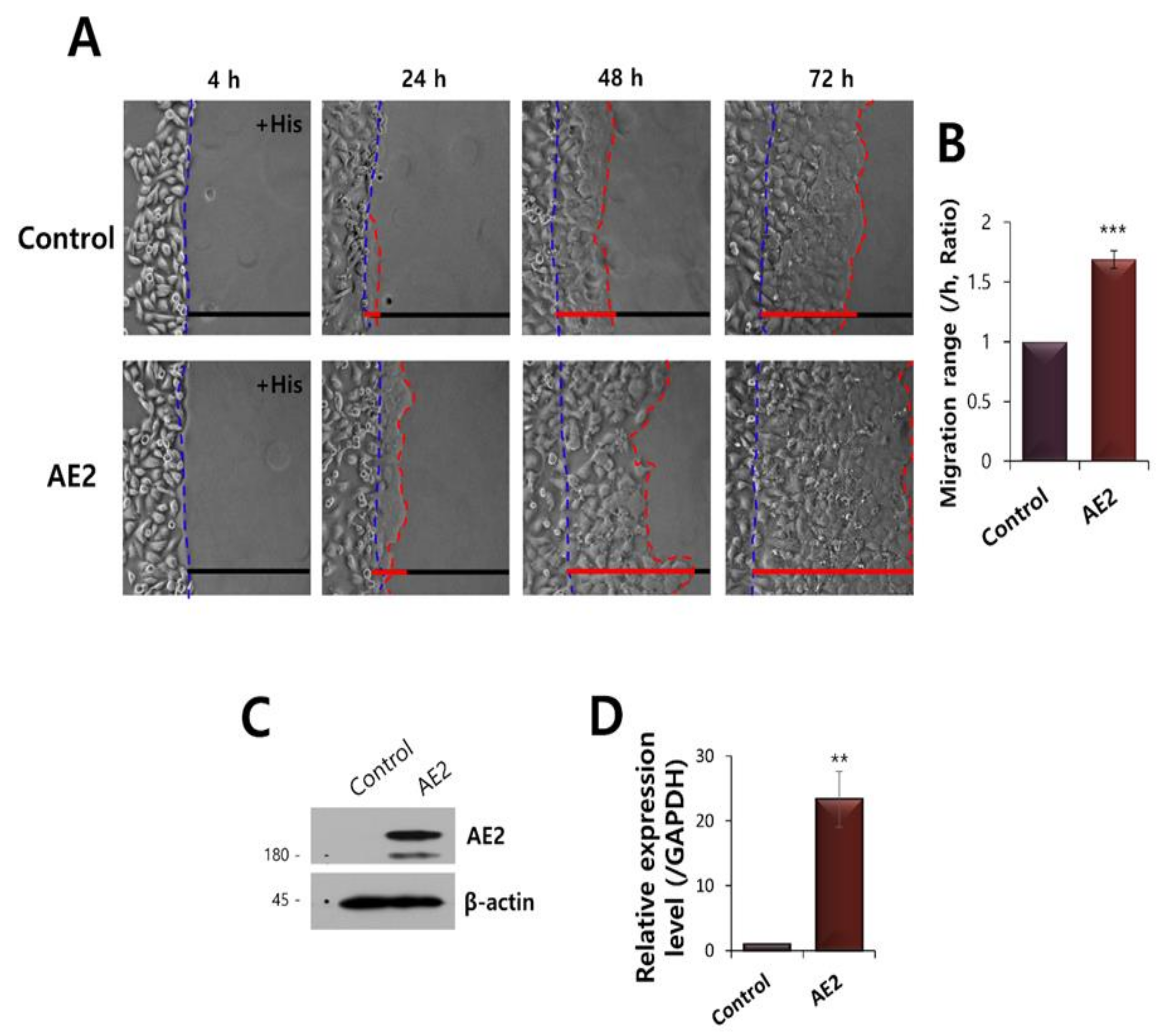
:
1. Introduction
2. Results
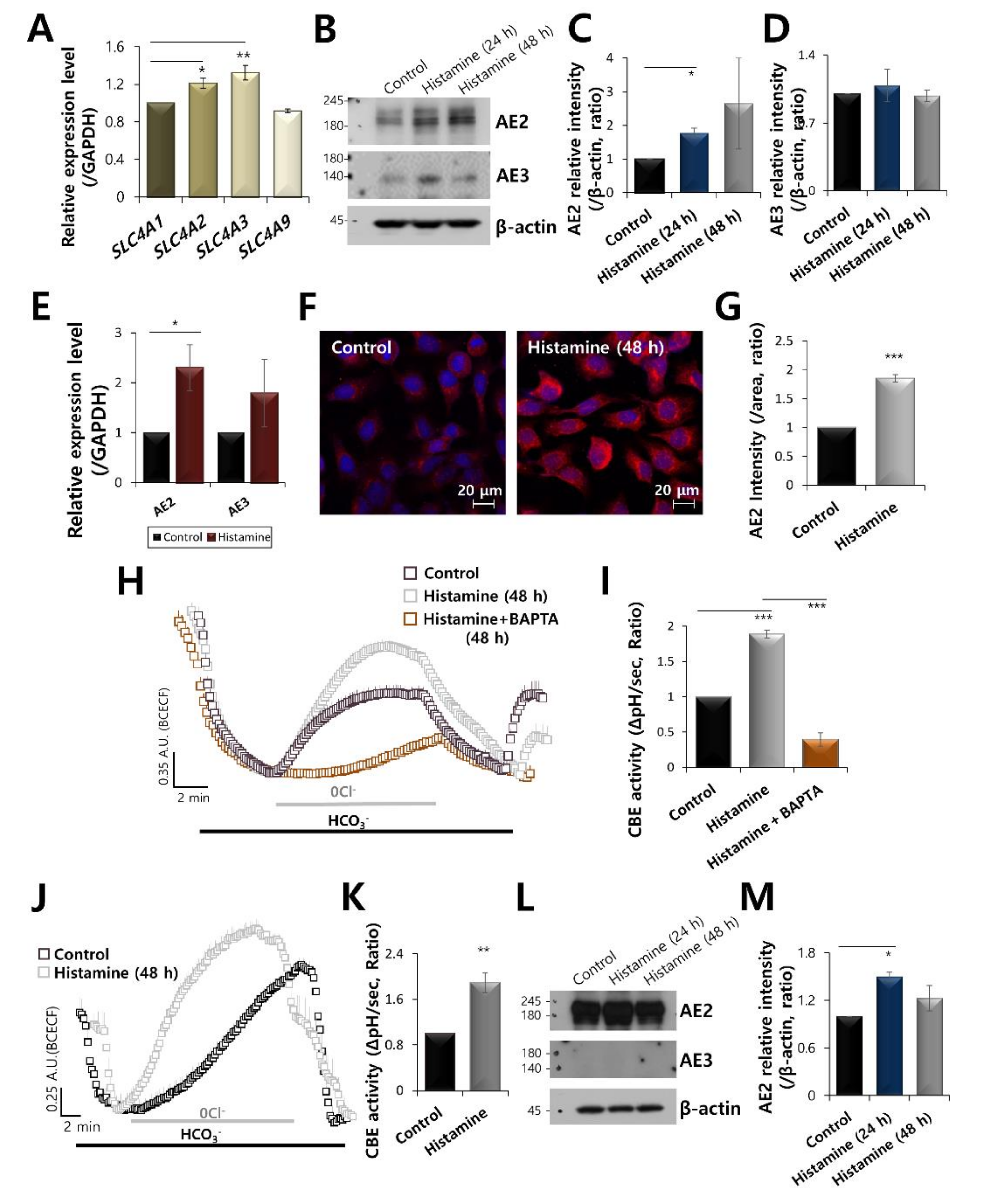
2.1. AE2 Is Activated by Stimulation of Histamine in Keratinocytes
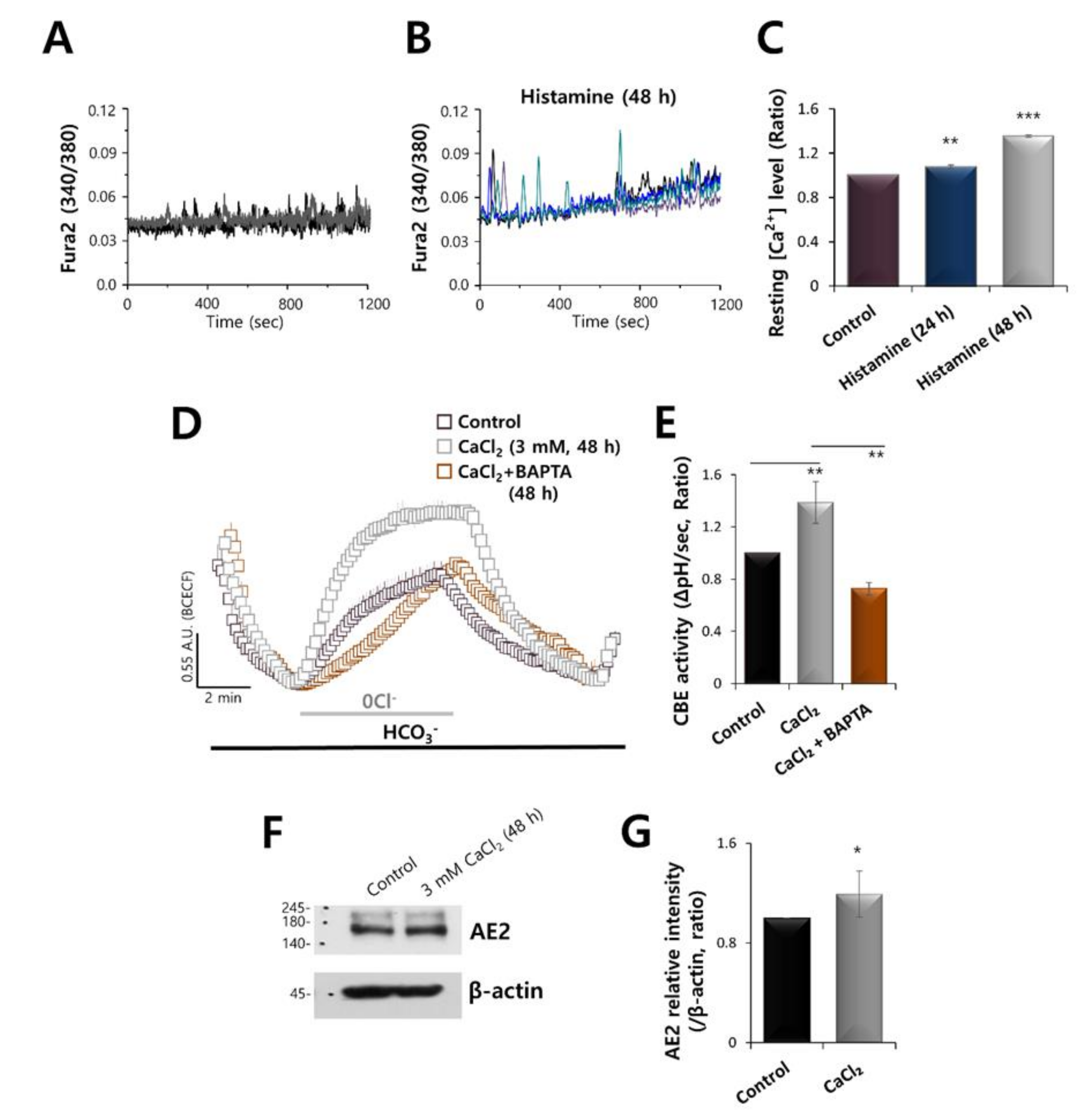
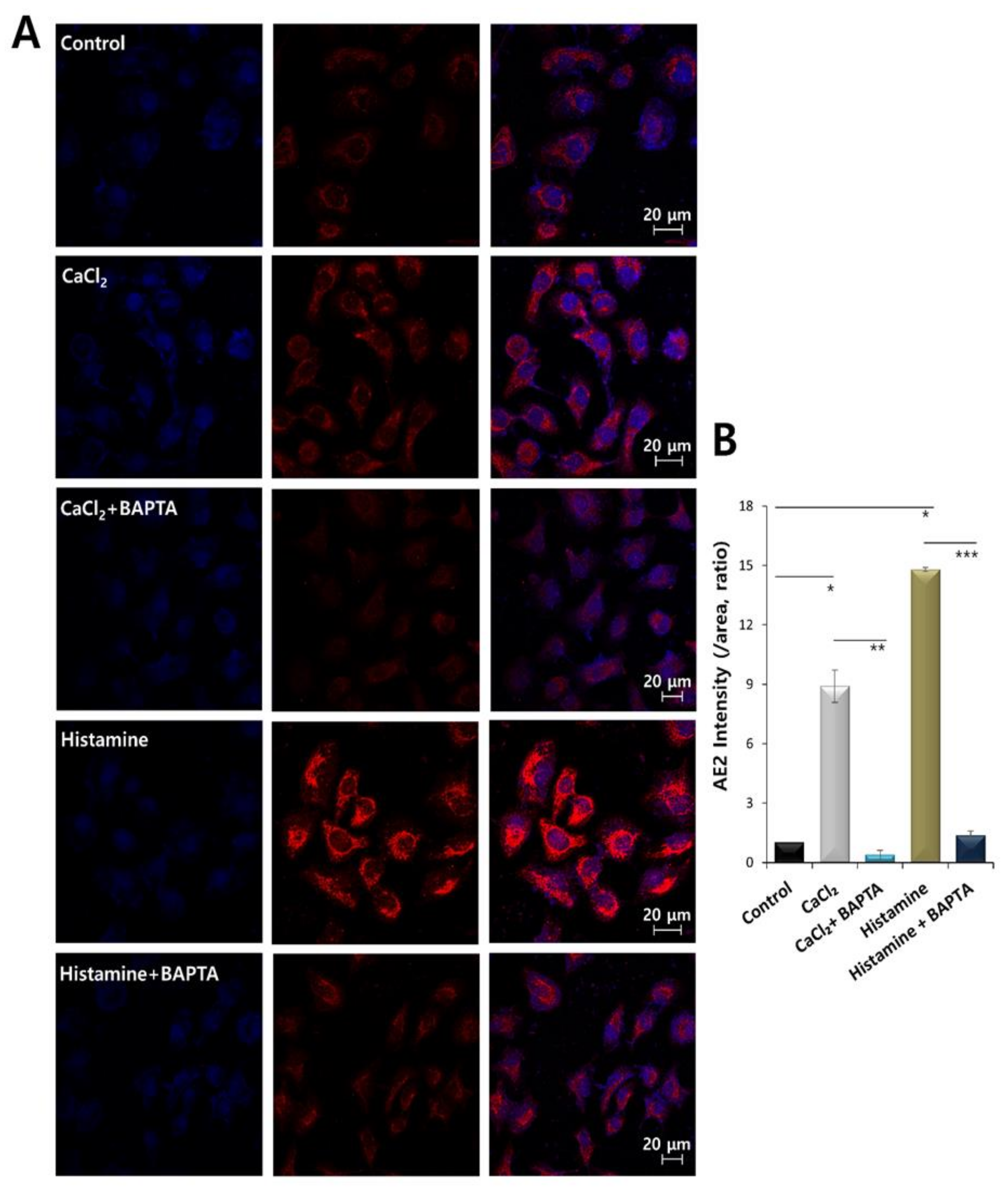
2.2. Increased Calcium Levels Induced by Histamine Stimulation Enhanced CBE Activity and AE2 Expression in HaCaT Keratinocytes
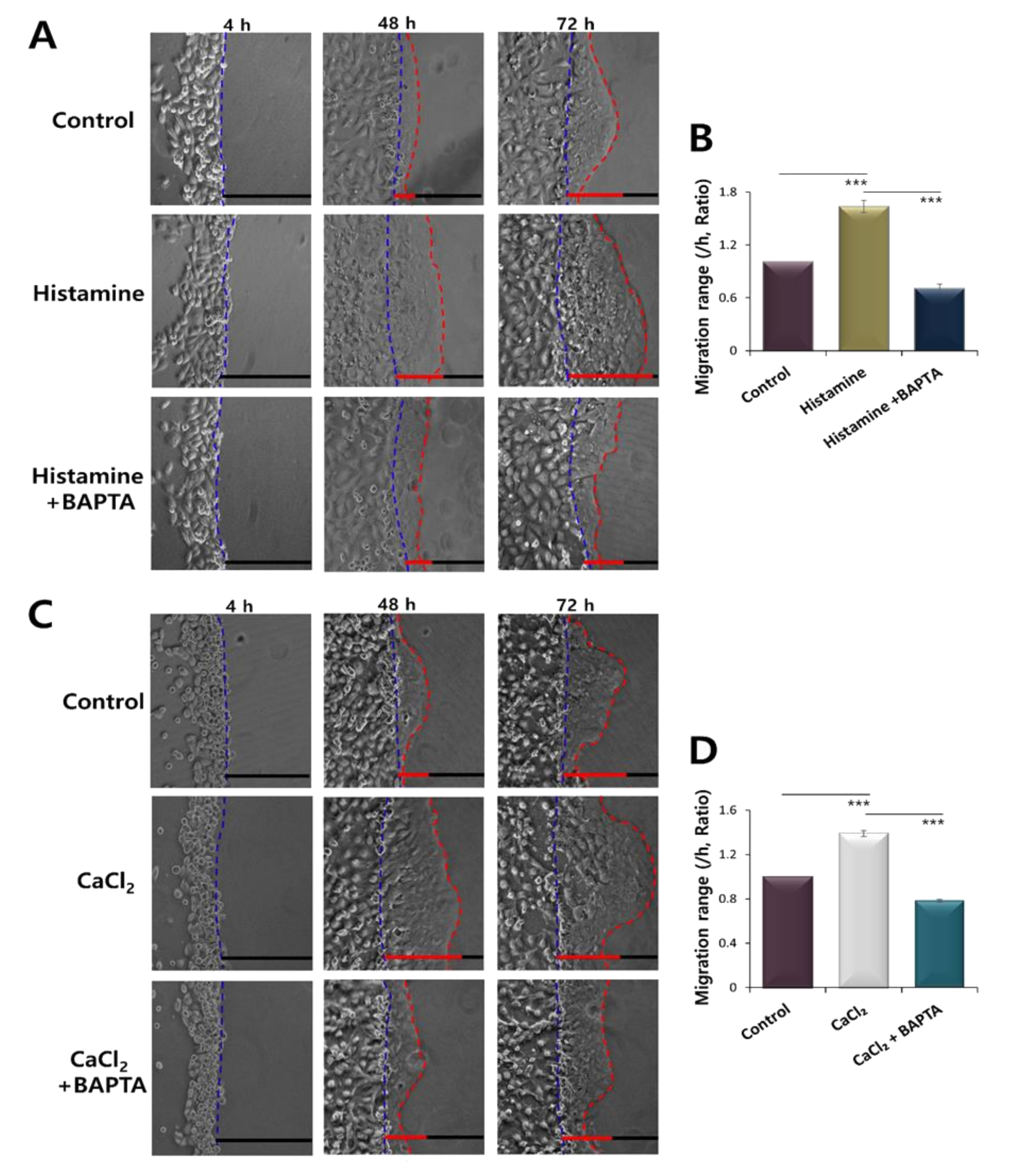
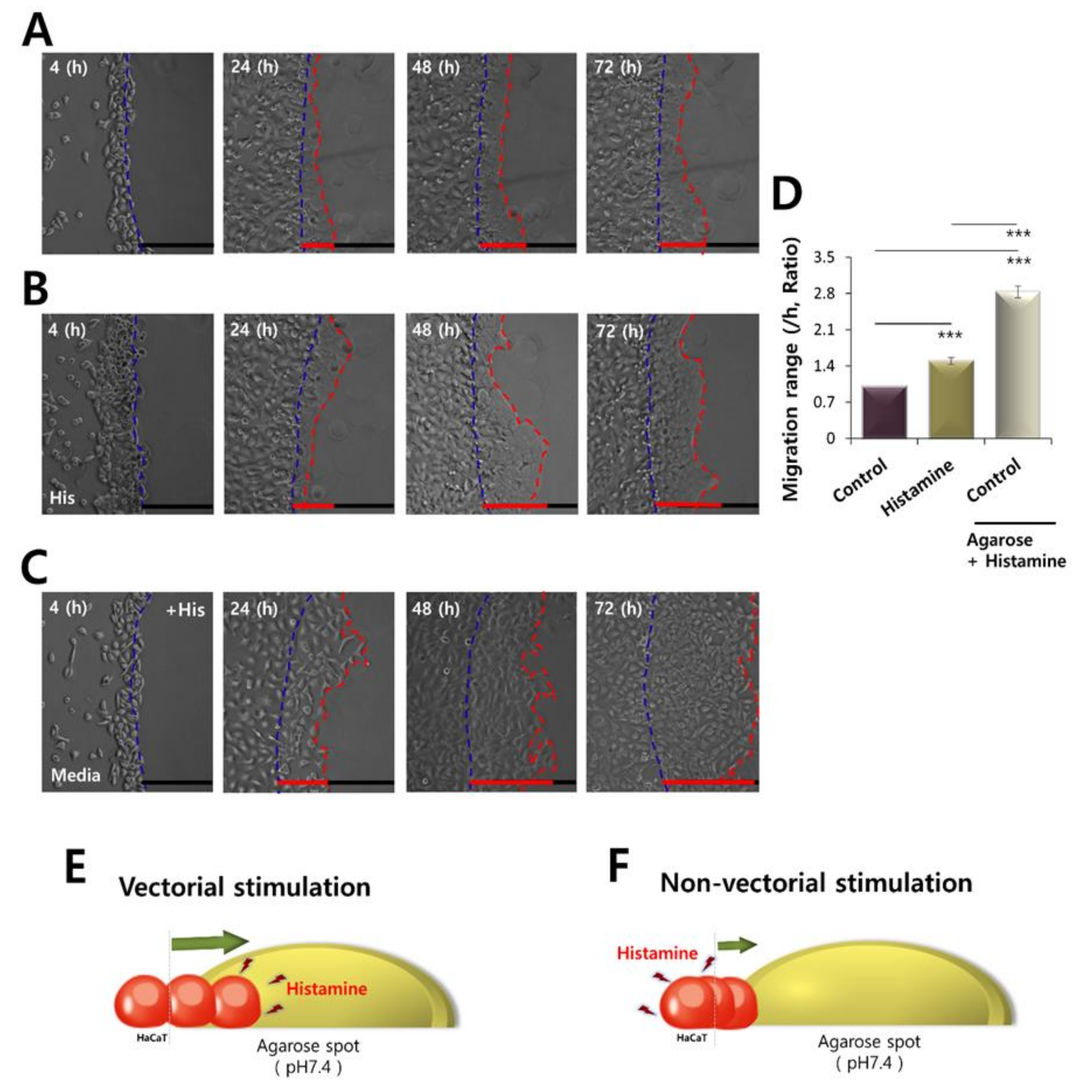
2.3. CaCl2 and Histamine Stimulation Enhanced HaCaT Cell Migration
2.4. AE2 Expression Was Dependent on Histamine and CaCl2 Stimulation
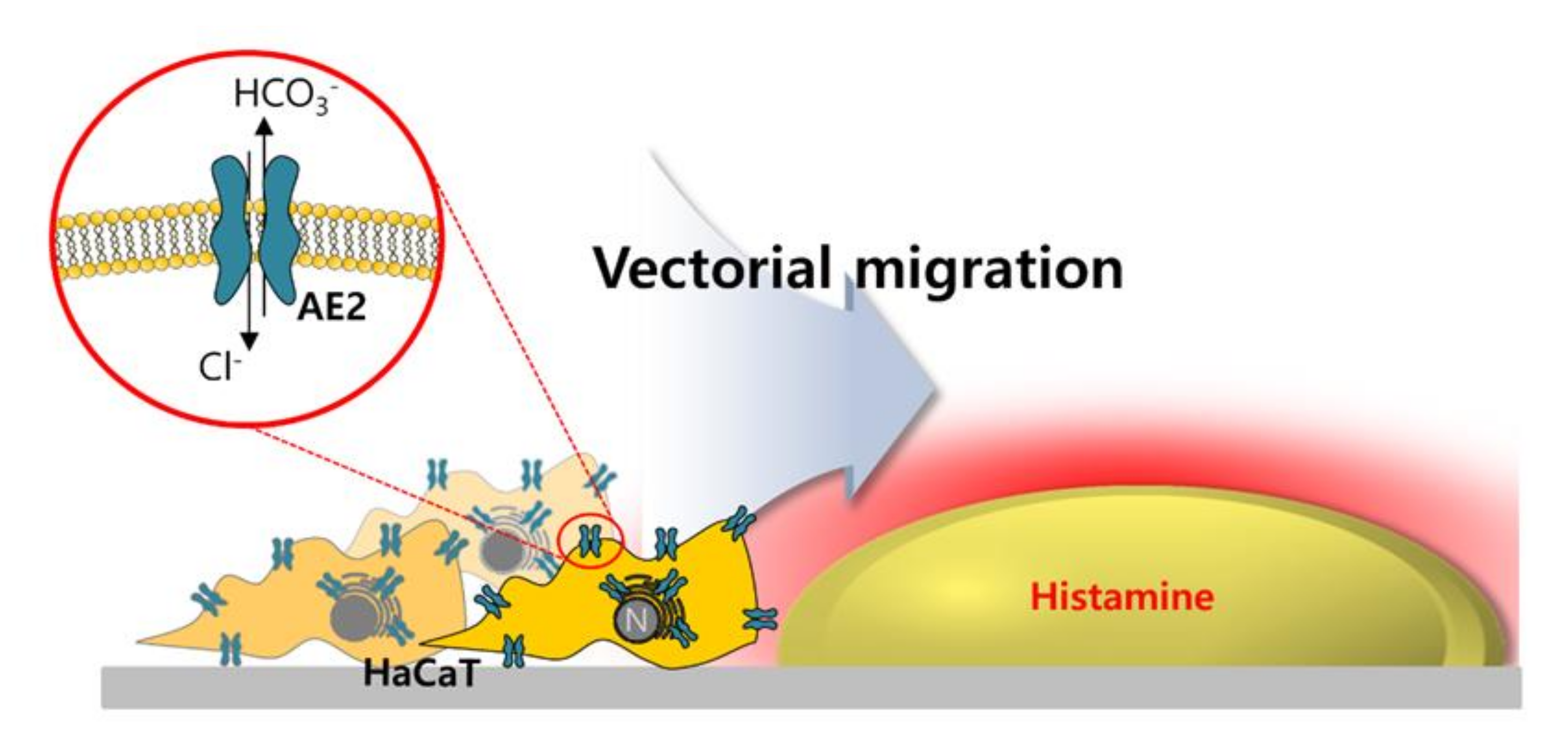
2.5. Motility of HaCaT Cells Was Driven More by Vectorial Chemotaxis
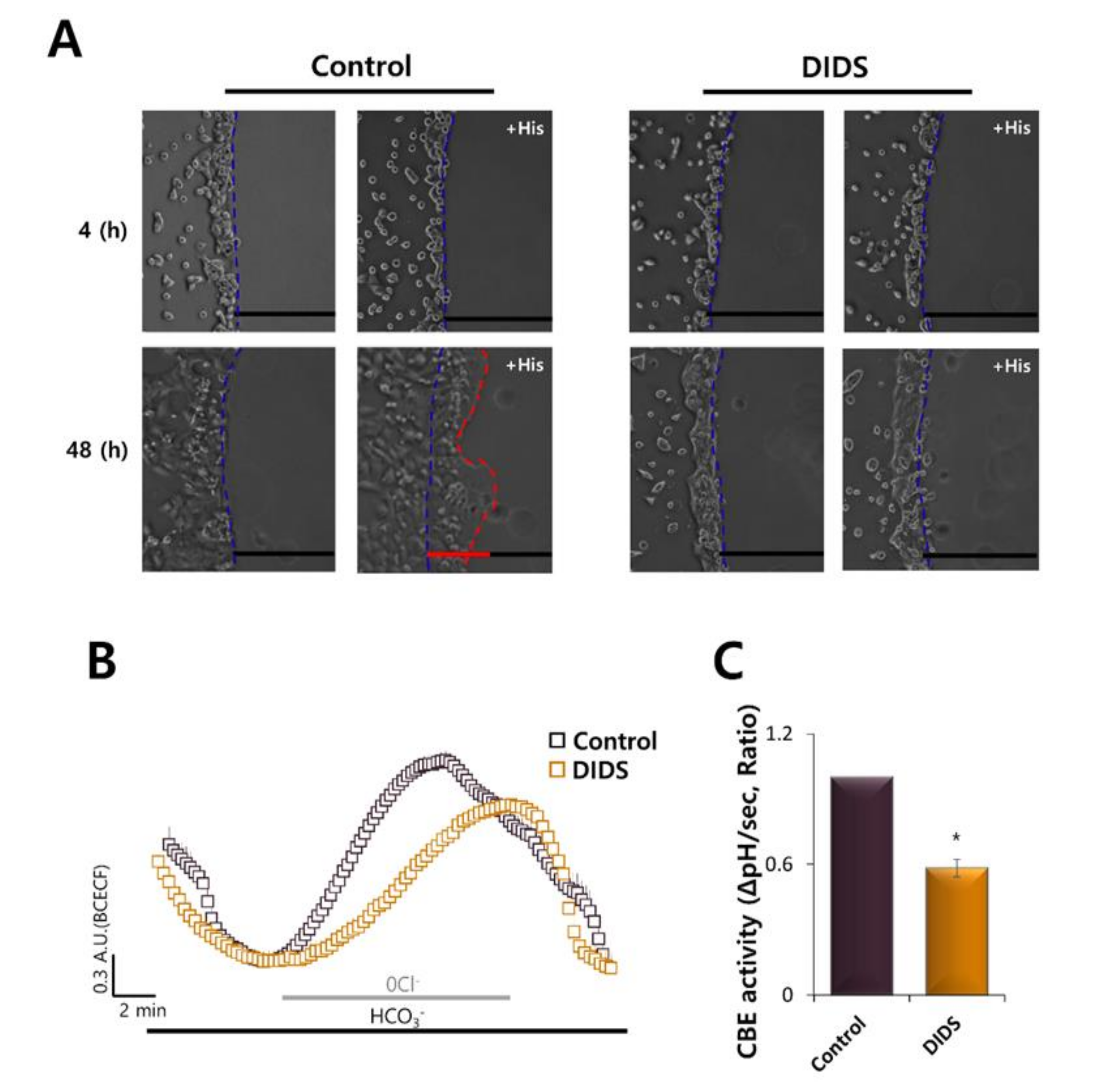
2.6. Inhibition of Transporters by DIDS Attenuated HaCaT Cell Migration
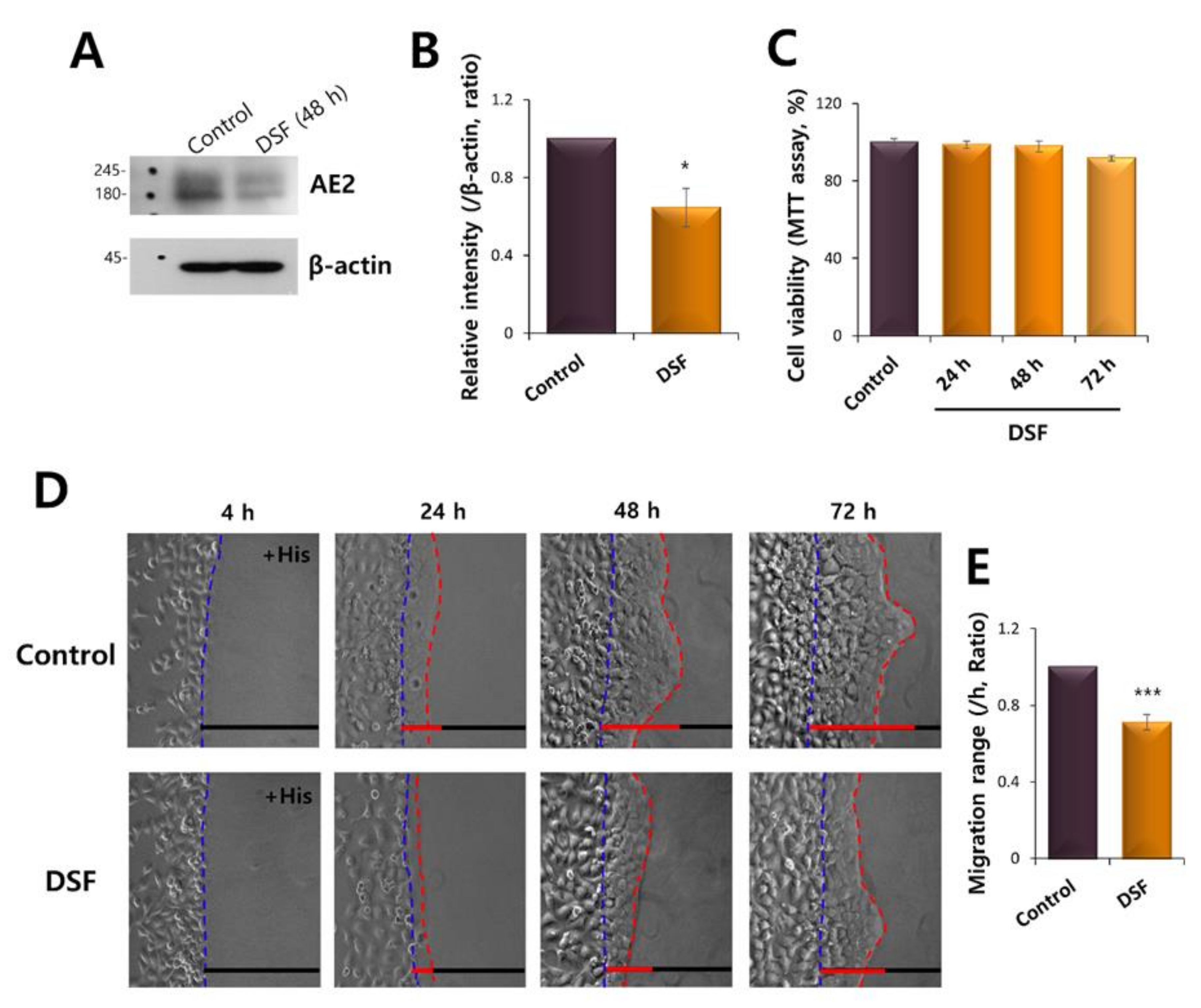
2.7. AE2 Inhibition Attenuated the Vectorial Movement of HaCaT Cells
2.8. Overexpressed AE2 Enhanced Vectorial HaCaT Migration
3. Discussion
4. Materials and Methods
4.1. Reagents and Plasmids
4.2. HaCaT and Primary Keratinocytes Culture
4.3. Agarose Spot Assay for Cell Migration
4.4. Measurement of the CBE Activity of AE2
4.5. Measurement of Intracellular Calcium Increase
4.6. Western Blotting
4.7. Confocal Imaging
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) and Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.9. DNA Transfection
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| His | Histamine |
| AE2 | Anion exchanger 2 |
| AE3 | Anion exchanger 3 |
| CBE | Chloride bicarbonate exchanger |
| H1R | Histamine receptor 1 |
| H2R | Histamine receptor 2 |
| H3R | Histamine receptor 3 |
| H4R | Histamine receptor 4 |
| GPCR | G protein-coupled receptor |
| DIDS | 4,4′-Diisothiocyano-2,2′-stilbenedisulfonic acid |
| DSF | Disulfiram |
| PBS | Phosphate-buffered saline |
| DAPI | 4,6-diamidino-2-phenylindole |
| BAPTA-AM | 1,2-bis (2-aminophenoxy) ethane-N,N,N′,N′-tetraacetic acid tetrakis, acetoxymethyl ester |
References
- O’Toole, E.A. Extracellular matrix and keratinocyte migration. Clin. Exp. Dermatol. 2001, 26, 525–530. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A Review of the Contribution of Mast Cells in Wound Healing: Involved Molecular and Cellular Mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef]
- Glatzer, F.; Gschwandtner, M.; Ehling, S.; Rossbach, K.; Janik, K.; Klos, A.; Baumer, W.; Kietzmann, M.; Werfel, T.; Gutzmer, R. Histamine induces proliferation in keratinocytes from patients with atopic dermatitis through the histamine 4 receptor. J. Allergy Clin. Immunol. 2013, 132, 1358–1367. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.D.; Boron, W.F. The divergence, actions, roles, and relatives of sodium-coupled bicarbonate transporters. Physiol. Rev. 2013, 93, 803–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Zhao, L.; Yang, Y.; Tian, H.; Suo, W.H.; Yan, M.; Fu, G.H. EGR1 is critical for gastrin-dependent upregulation of anion exchanger 2 in gastric cancer cells. FEBS J. 2013, 280, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Schwab, A.; Fabian, A.; Hanley, P.J.; Stock, C. Role of ion channels and transporters in cell migration. Physiol. Rev. 2012, 92, 1865–1913. [Google Scholar] [CrossRef] [PubMed]
- Simmen, H.P.; Blaser, J. Analysis of pH and pO2 in abscesses, peritoneal fluid, and drainage fluid in the presence or absence of bacterial infection during and after abdominal surgery. Am. J. Surg. 1993, 166, 24–27. [Google Scholar] [CrossRef]
- Komuro, H.; Kumada, T. Ca2+ transients control CNS neuronal migration. Cell Calcium 2005, 37, 387–393. [Google Scholar] [CrossRef]
- Wang, F.; Chen, S.; Liu, H.B.; Parent, C.A.; Coulombe, P.A. Keratin 6 regulates collective keratinocyte migration by altering cell-cell and cell-matrix adhesion. J. Cell. Biol. 2018, 217, 4314–4330. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, G.; Jia, N.; Wang, W.; Wang, Y.; Yin, M.; Jiang, X.; Huang, Y.; Zhang, J. CD9 regulates keratinocyte migration by negatively modulating the sheddase activity of ADAM17. Int. J. Biol. Sci. 2019, 15, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Kular, L.; Vij, M.; Herter, E.K.; Li, X.; Wang, A.; Chu, T.; Toma, M.A.; Zhang, L.; Liapi, E.; et al. Human skin long noncoding RNA WAKMAR1 regulates wound healing by enhancing keratinocyte migration. Proc. Natl. Acad. Sci. USA 2019, 116, 9443–9452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, D.; Yan, T.; Jiang, X.; Zhang, C.; Zhao, L.; Li, L.; Tang, D.; Zhang, Q.; Jia, J.; et al. BNIP3 promotes the motility and migration of keratinocyte under hypoxia. Exp. Dermatol. 2017, 26, 416–422. [Google Scholar] [CrossRef]
- Na, J.; Shin, J.Y.; Jeong, H.; Lee, J.Y.; Kim, B.J.; Kim, W.S.; Yune, T.Y.; Ju, B.G. JMJD3 and NF-kappaB-dependent activation of Notch1 gene is required for keratinocyte migration during skin wound healing. Sci. Rep. 2017, 7, 6494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, R.; Yang, S.; He, W.; Wang, F.; Tan, J.; Zhou, J.; Yang, S.; Yao, Z.; Wu, J.; Luo, G. Nitric oxide enhances keratinocyte cell migration by regulating Rho GTPase via cGMP-PKG signalling. PLoS ONE 2015, 10, e0121551. [Google Scholar] [CrossRef]
- Schwab, A.; Stock, C. Ion channels and transporters in tumour cell migration and invasion. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, M.F.; Fulton, C.M.; Boron, W.F. The SLC4 family of HCO3− Transporters. Pflug. Arch. 2004, 447, 495–509. [Google Scholar] [CrossRef]
- Jeong, Y.S.; Hong, J.H. Governing effect of regulatory proteins for Cl(−)/HCO3(−) exchanger 2 activity. Channels 2016, 10, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Coruzzi, G.; Adami, M.; Guaita, E.; de Esch, I.J.; Leurs, R. Antiinflammatory and antinociceptive effects of the selective histamine H4-receptor antagonists JNJ7777120 and VUF6002 in a rat model of carrageenan-induced acute inflammation. Eur. J. Pharmacol. 2007, 563, 240–244. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Purwar, R.; Wittmann, M.; Baumer, W.; Kietzmann, M.; Werfel, T.; Gutzmer, R. Histamine upregulates keratinocyte MMP-9 production via the histamine H1 receptor. J. Investig. Dermatol. 2008, 128, 2783–2791. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Shin, D.M.; Hong, J.H. Drug Repurposing as an Antitumor Agent: Disulfiram-Mediated Carbonic Anhydrase 12 and Anion Exchanger 2 Modulation to Inhibit Cancer Cell Migration. Molecules 2019, 24, 3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behne, M.J.; Meyer, J.W.; Hanson, K.M.; Barry, N.P.; Murata, S.; Crumrine, D.; Clegg, R.W.; Gratton, E.; Holleran, W.M.; Elias, P.M.; et al. NHE1 regulates the stratum corneum permeability barrier homeostasis. Microenvironment acidification assessed with fluorescence lifetime imaging. J. Biol. Chem. 2002, 277, 47399–47406. [Google Scholar] [CrossRef] [Green Version]
- Hachem, J.P.; Behne, M.; Aronchik, I.; Demerjian, M.; Feingold, K.R.; Elias, P.M.; Mauro, T.M. Extracellular pH Controls NHE1 expression in epidermis and keratinocytes: Implications for barrier repair. J. Investig. Dermatol. 2005, 125, 790–797. [Google Scholar] [CrossRef] [Green Version]
- Ng, F.L.; Boedtkjer, E.; Witkowska, K.; Ren, M.; Zhang, R.; Tucker, A.; Aalkjaer, C.; Caulfield, M.J.; Ye, S. Increased NBCn1 expression, Na+/HCO3− co-transport and intracellular pH in human vascular smooth muscle cells with a risk allele for hypertension. Hum. Mol. Genet. 2017, 26, 989–1002. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes. | Tm (°C) | Sequences (5′ → 3′) |
|---|---|---|
| SLC4A1 | 60 | (F) CAC ACA ACT TCA GGC CCC TC |
| (R) AGA GCC TGC TGT CTC CTA CC | ||
| SLC4A2 | 59 | (F) AGT TGG GAG AAG TTG GGA GC |
| (R) CAT AAC CCG CTC GCT CTG G | ||
| SLC4A3 | 58 | (F) GTT TGG GGA CCT CAT CAG CA |
| (R) ATG TGT GCC GGT GAA ACT CA | ||
| SLC4A9 | 58 | (F) GAC CCC AGG AAA CAG CAT GA |
| (R) CAC CCT CAG GTC AGG AGG TA | ||
| GAPDH | 59 | (F) CCG CAT CTT CTT TTG CGT CG |
| (R) TTC CCG TTC TCA GCC TTG AC |
| Genes | Tm (°C) | Sequences (5′ → 3′) |
|---|---|---|
| Histamine 1 receptor (R) | 58 | (F) GAC TGT GTA GCC GTC AAC CGG A |
| (R) TGA AGG CTG CCA TGA TAA AAC C | ||
| Histamine 2R | 56 | (F) TCG TGT CCT TGG CTA TCA C |
| (R) CTT TGC TGG TCT CGT TCC T | ||
| Histamine 3R | 70 | (F) TCA GCT ACG ACC GCT TCC TGT CGG TCA C |
| (R) TTG AGT GAG CGC GGC CTC TCA GTG CCC C | ||
| Histamine 4R | 63 | (F) GAA TTG TCT GGC TGG ATT AAT TTG CTA ATT TG |
| (R) AAG AAT GAT GTG GTG ATG GCA AGG ATG TAC C | ||
| GAPDH | 62 | (F) CAT GGC ACC GTC AAG GCT GAG |
| (R) CTT GGC CAG GGG TGC TAA GC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.; Shin, D.M.; Hong, J.H. Intracellular Ca2+-Mediated AE2 Is Involved in the Vectorial Movement of HaCaT Keratinocyte. Int. J. Mol. Sci. 2020, 21, 8429. https://doi.org/10.3390/ijms21228429
Hwang S, Shin DM, Hong JH. Intracellular Ca2+-Mediated AE2 Is Involved in the Vectorial Movement of HaCaT Keratinocyte. International Journal of Molecular Sciences. 2020; 21(22):8429. https://doi.org/10.3390/ijms21228429
Chicago/Turabian StyleHwang, Soyoung, Dong Min Shin, and Jeong Hee Hong. 2020. "Intracellular Ca2+-Mediated AE2 Is Involved in the Vectorial Movement of HaCaT Keratinocyte" International Journal of Molecular Sciences 21, no. 22: 8429. https://doi.org/10.3390/ijms21228429