Identification and Characterization of a β-N-Acetylhexosaminidase with a Biosynthetic Activity from the Marine Bacterium Paraglaciecola hydrolytica S66T

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Putative β-NAHAs in P. hydrolytica and Organization of Vicinal Genomic Regions
2.2. Phylogenetic Analysis of PhNah20A and PhNah20B
2.3. Cloning and Production of β-NAHA
2.4. Characterization of PhNah20A
2.4.1. Enzyme Stability
2.4.2. pH and Temperature Optima
2.4.3. Substrate Specificity and Kinetic Parameters of PhNah20A
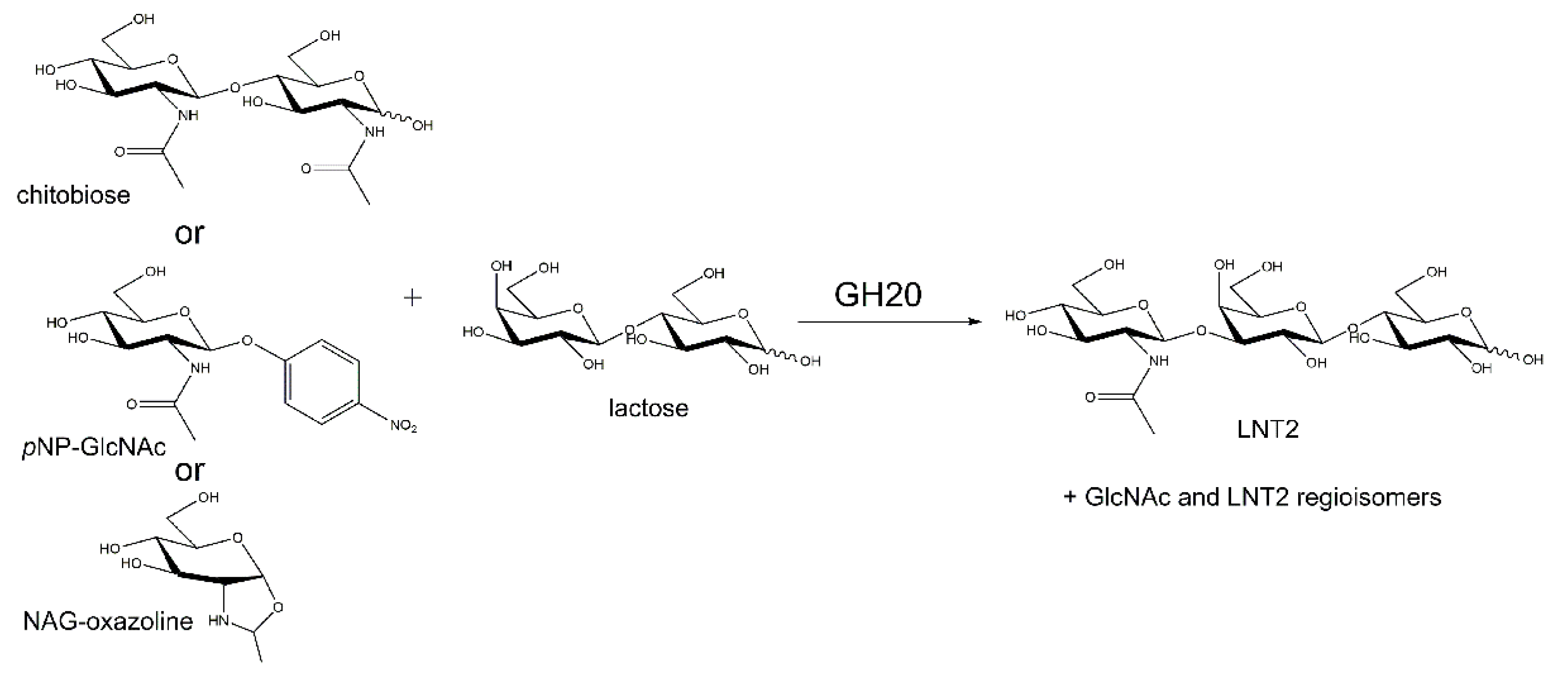
2.5. Transglycosylation by PhNah20A
3. Materials and Methods
3.1. Materials
3.2. Bacterial Strains and Media
3.3. Molecular Cloning and Plasmids
3.4. Recombinant Protein Production and Purification
3.5. Activity Assays
3.6. Kinetics
3.7. Synthesis of NAG-Oxazoline
3.8. Transglycosylation
3.9. Chromatographic Methods
3.10. Nuclear Magnetic Resonance (NMR)
3.11. In Silico Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | bovine serum albumin |
| DCM | dichloromethane |
| GH | glycoside hydrolase |
| GlcNAc | N-acetylglucosamine |
| (GlcNAc)2 | N,N’-diacetylchitobiose, chitobiose |
| GPC | gel permeation chromatography |
| HMOs | human milk oligosaccharides |
| HPAEC-PAD | high-performance anion exchange chromatography with pulsed amperometric detector |
| IPTG | isopropyl thio-β-d-galactoside |
| LNT2 | lacto-N-triose II |
| NAG-oxazoline | 2-methyl-(1,2-dideoxy-α-d-glucopyrano)-oxazoline |
| β-NAHA | β-N-acetylhexosaminidases |
| NCBI | National Center for Biotechnology Information |
| NMR | nuclear magnetic resonance |
| pNPGlcNAc | p-nitrophenyl-GlcNAc |
| pNPGalNAc | p-nitrophenyl-GalNAc |
| TLC | thin layer chromatography |
| X-GlcNAc | 5-bromo-4-chloro-3-indolyl N-acetyl-β-d-glucosaminide |
References
- Schultz-Johansen, M.; Glaring, M.A.; Bech, P.K.; Stougaard, P. Draft genome sequence of a novel marine bacterium, Paraglaciecola sp. strain S66, with hydrolytic activity against seaweed. Genome Announc. 2016, 4, e00304-16. [Google Scholar] [CrossRef] [Green Version]
- Bech, P.K.; Schultz-Johansen, M.; Glaring, M.A.; Barbeyron, T.; Czjzek, M.; Stougaard, P. Paraglaciecola hydrolytica sp. nov., a bacterium with hydrolytic activity against multiple seaweed-derived polysaccharides. Int. J. Syst. Evol. Microbiol. 2017, 67, 2242–2247. [Google Scholar] [CrossRef]
- Schultz-Johansen, M.; Bech, P.; Hennessy, R.; Glaring, M.; Barbeyron, T.; Czjzek, M.; Stougaard, P. A novel enzyme portfolio for red algal polysaccharide degradation in the marine bacterium Paraglaciecola hydrolytica S66T encoded in a sizeable polysaccharide. Front. Microbiol. 2018, 9, 839. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Slámová, K.; Bojarová, P. Engineered N-acetylhexosamine-active enzymes in glycoscience. Biochim. Biophys. Acta 2017, 1861, 2070–2087. [Google Scholar] [CrossRef]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef]
- Kunz, C.; Kuntz, S.; Rudloff, S. Bioactivity of human milk oligosaccharides. In Food Oligosaccharides: Production, Analysis and Bioactivity; Moreno, J.F., Sanz, M.L., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2014; pp. 1–20. [Google Scholar]
- Barile, D.; Rastall, R.A. Human milk and related oligosaccharides as prebiotics. Curr. Opin. Biotechnol. 2013, 24, 214–219. [Google Scholar] [CrossRef]
- Verspreet, J.; Damen, B.; Broekaert, W.F.; Verbeke, K.; Delcour, J.A.; Courtin, C.M. A critical look at prebiotics within the dietary fiber concept. Annu. Rev. Food Sci. Technol. 2016, 7, 167–190. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Boltje, T.J.; Buskas, T.; Boons, G.J. Opportunities and challenges in synthetic oligosaccharide and glycoconjugate research. Nat. Chem. 2009, 1, 611–622. [Google Scholar] [CrossRef]
- Zeuner, B.; Teze, D.; Muschiol, J.; Meyer, A.S. Synthesis of human milk oligosaccharides: Protein engineering strategies for improved enzymatic transglycosylation. Molecules 2019, 24, 2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeuner, B.; Jers, C.; Mikkelsen, J.D.; Meyer, A.S. Methods for improving enzymatic trans-glycosylation for synthesis of human milk oligosaccharide biomimetics. J. Agric. Food Chem. 2014, 62, 9615–9631. [Google Scholar] [CrossRef] [PubMed]
- Jamek, S.B.; Nyffenegger, C.; Muschiol, J.; Holck, J.; Meyer, A.S.; Mikkelsen, J.D. Characterization of two novel bacterial type A exo-chitobiose hydrolases having C-terminal 5/12-type carbohydrate-binding modules. Appl. Microbiol. Biotechnol. 2017, 101, 4533–4546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Xu, L.; Jin, L.; Sun, B.; Gu, G.; Lu, L.; Xiao, M. Efficient and regioselective synthesis of β-GalNAc/GlcNAc-lactose by a bifunctional transglycosylating β-N-acetylhexosaminidase from Bifidobacterium bifidum. Appl. Environ. Microbiol. 2016, 82, 5642–5652. [Google Scholar] [CrossRef] [Green Version]
- Baumgärtner, F.; Conrad, J.; Sprenger, G.A.; Albermann, C. Synthesis of the human milk oligosaccharide lacto-N-tetraose in metabolically engineered, plasmid-free E. coli. ChemBioChem 2014, 15, 1896–1900. [Google Scholar] [CrossRef]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Nyffenegger, C.; Nordvang, R.T.; Zeuner, B.; Łężyk, M.; Difilippo, E.; Logtenberg, M.J.; Schols, H.A.; Meyer, A.S.; Mikkelsen, J.D. Backbone structures in human milk oligosaccharides: Trans-glycosylation by metagenomic β-N-acetylhexosaminidases. Appl. Microbiol. Biotechnol. 2015, 99, 7997–8009. [Google Scholar] [CrossRef]
- Murata, T.; Tashiro, A.; Itoh, T.; Usui, T. Enzymic synthesis of 3′-O- and 6′-O-N-acetylglucosaminyl-N-acetyllactosaminide glycosides catalyzed by β-N-acetyl-d-hexosaminidase from Nocardia orientalis. Biochim. Biophys. Acta 1997, 1335, 326–334. [Google Scholar] [CrossRef]
- Slámová, K.; Gažák, R.; Bojarová, P.; Kulik, N.; Ettrich, R.; Pelantová, H.; Sedmera, P.; Křen, V. 4-deoxy-substrates for β-N-acetylhexosaminidases: How to make use of their loose specificity. Glycobiology 2010, 20, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Tsujibo, H.; Kondo, N.; Tanaka, K.; Miyamoto, K.; Baba, N.; Inamori, Y. Molecular analysis of the gene encoding a novel transglycosylative enzyme from Alteromonas sp. strain O-7 and its physiological role in the chitinolytic system. J. Bacteriol. 1999, 181, 5461–5466. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, T.; Loganathan, D. Enzymatic synthesis of N-glycoprotein linkage region disaccharide mimetics using β-N-acetylhexosaminidases from Aspergillus oryzae and Vigna radiata. Tetrahedron Asymmetry 2005, 16, 255–260. [Google Scholar] [CrossRef]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef] [Green Version]
- Slámová, K.; Bojarová, P.; Petrásková, L.; Křen, V. β-N-Acetylhexosaminidase: What’s in a name...? Biotechnol. Adv. 2010, 28, 682–693. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, J.; Song, Z.; Huang, Z. Enzymatic properties of beta-N-acetylglucosaminidases. Appl. Microbiol. Biotechnol. 2018, 102, 93–103. [Google Scholar] [CrossRef]
- Choi, K.H.; Seo, J.Y.; Park, K.M.; Park, C.S.; Cha, J. Characterization of glycosyl hydrolase family 3β-N-acetylglucosaminidases from Thermotoga maritima and Thermotoga neapolitana. J. Biosci. Bioeng. 2009, 108, 455–459. [Google Scholar] [CrossRef]
- Ryšlavá, H.; Kalendová, A.; Doubnerová, V.; Skočdopol, P.; Kumar, V.; Kukačka, Z.; Pompach, P.; Vaněk, O.; Slámová, K.; Bojarová, P.; et al. Enzymatic characterization and molecular modeling of an evolutionarily interesting fungal β-N-acetylhexosaminidase. FEBS J. 2011, 278, 2469–2484. [Google Scholar] [CrossRef]
- Piszkiewicz, D.; Bruice, T.C.; Glycoside, H., III. Intramolecular acetamido group participation in the specific acid catalyzed hydrolysis of methyl 2-acetamido-2-deoxy-β-d-glucopyranoside. J. Am. Chem. Soc. 1968, 378, 5844–5848. [Google Scholar] [CrossRef]
- Knapp, S.; Vocadlo, D.; Gao, Z.; Kirk, B.; Lou, J.; Withers, S.G. NAG-thiazoline, an N-acetyl-β-hexosaminidase inhibitor that implicates acetamido participation. J. Am. Chem. Soc. 1996, 118, 6804–6805. [Google Scholar] [CrossRef]
- Ferrara, M.C.; Cobucci-Ponzano, B.; Carpentieri, A.; Henrissat, B.; Rossi, M.; Amoresano, A.; Moracci, M. The identification and molecular characterization of the first archaeal bifunctional exo-β-glucosidase/N-acetyl-β-glucosaminidase demonstrate that family GH116 is made of three functionally distinct subfamilies. Biochim. Biophys. Acta 2014, 1840, 367–377. [Google Scholar] [CrossRef]
- Teze, D.; Shuoker, B.; Chaberski, E.K.; Kunstmann, R.S.; Fredslund, F.; Peters, G.H.J.; Karlsson, E.N.; Welner, D.H.; Abou Hachem, M. The catalytic acid-base in GH109 resides in a conserved GGHGG loop and allows for comparable α-retaining and β-inverting activity in an N-acetylgalactosaminidase from Akkermansia muciniphila. ChemRxiv 2019, preprint. [Google Scholar]
- Placzek, S.; Schomburg, I.; Chang, A.; Jeske, L.; Ulbrich, M.; Tillack, J.; Schomburg, D. BRENDA in 2017: New perspectives and new tools in BRENDA. Nucleic Acids Res. 2017, 45, D380–D388. [Google Scholar] [CrossRef] [PubMed]
- Keyhani, N.O.; Roseman, S. The chitin catabolic cascade in the marine bacterium Vibrio furnissii. J. Biol. Chem. 1996, 271, 33425–33432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, X.; Zhang, X.; Kodaira, R.; Zhou, Z.; Shimosaka, M. Gene cloning, expression, and characterization of a second β-N-acetylglucosaminidase from the chitinolytic bacterium Aeromonas hydrophila strain SUWA-9. Biosci. Biotechnol. Biochem. 2008, 72, 492–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsujibo, H.; Miyamoto, K.; Yoshimura, M.; Takata, M.; Miyamoto, J.; Inamori, Y. Molecular cloning of the gene encoding a novel β-N-acetylhexosaminidase from a marine bacterium, Alteromonas sp. strain O-7, and characterization of the cloned enzyme. Biosci. Biotechnol. Biochem. 2002, 66, 471–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, X.; Ozawa, N.; Nishiwaki, N.; Kodaira, R.; Okazaki, M.; Shimosaka, M. Purification, cloning, and sequence analysis of β-N-acetylglucosaminidase from the chitinolytic bacterium Aeromonas hydrophila strain SUWA-9. Biosci. Biotechnol. Biochem. 2004, 68, 1082–1090. [Google Scholar] [CrossRef]
- Soto-Gil, R.W.; Zyskind, J.W. N,N’-Diacetylchitobiase of Vibrio harveyi. Primary structure, processing, and evolutionary relationships. J. Biol. Chem. 1989, 264, 14778–14783. [Google Scholar]
- Mark, B.L.; Wasney, G.A.; Salo, T.J.; Khan, A.R.; Cao, Z.; Robbins, P.W.; James, M.N.; Triggs-Raine, B.L. Structural and functional characterization of Streptomyces plicatus beta-N-acetylhexosaminidase by comparative molecular modeling and site-directed mutagenesis. J. Biol. Chem. 1998, 273, 19618–19624. [Google Scholar] [CrossRef] [Green Version]
- Mayer, C.; Vocadlo, D.J.; Mah, M.; Rupitz, K.; Stoll, D.; Warren, R.A.J.; Withers, S.G. Characterization of a β-N-acetylhexosaminidase and a β-N-acetylglucosaminidase/β-glucosidase from Cellulomonas fimi. FEBS J. 2006, 273, 2929–2941. [Google Scholar] [CrossRef]
- Miwa, M.; Horimoto, T.; Kiyohara, M.; Katayama, T.; Kitaoka, M.; Ashida, H.; Yamamoto, K. Cooperation of β-galactosidase and β-N-acetylhexosaminidase from bifidobacteria in assimilation of human milk oligosaccharides with type 2 structure. Glycobiology 2010, 20, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Gutternigg, M.; Rendić, D.; Voglauer, R.; Iskratsch, T.; Wilson, I.B.H. Mammalian cells contain a second nucleocytoplasmic hexosaminidase. Biochem. J. 2009, 419, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Verpoorte, J.A. Isolation and characterization of the major beta-N-acetyl-d-glucosaminidase from human plasma. Biochemistry 1974, 13, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alonso, J.; Reglero, A.; Cabezas, J.A. Purification and properties of β-N-acetylhexosaminidase a from pig brain. Int. J. Biochem. 1990, 22, 645–651. [Google Scholar] [CrossRef]
- Zhou, J.; Song, Z.; Zhang, R.; Liu, R.; Wu, Q.; Li, J.; Tang, X.; Xu, B.; Ding, J.; Han, N.; et al. Distinctive molecular and biochemical characteristics of a glycoside hydrolase family 20 β-N-acetylglucosaminidase and salt tolerance. BMC Biotechnol. 2017, 17, 37. [Google Scholar] [CrossRef] [PubMed]
- Sumida, T.; Ishii, R.; Yanagisawa, T.; Yokoyama, S.; Ito, M. Molecular coning and crystal structural analysis of a novel β-N-acetylhexosaminidase from Paenibacillus sp. TS12 capable of degrading glycosphingolipids. J. Mol. Biol. 2009, 392, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-L.; Yu, W.-L.; Zhang, J.-W.; Frolet, C.; Di Guilmi, A.-M.; Zhou, C.-Z.; Vernet, T.; Chen, Y. Structural basis for the substrate specificity of a novel β-N-acetylhexosaminidase StrH protein from Streptococcus pneumoniae R6. J. Biol. Chem. 2011, 286, 43004–43012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thi, N.N.; Offen, W.A.; Shareck, F.; Davies, G.J.; Doucet, N. Structure and activity of the Streptomyces coelicolor A3(2) β-N-acetylhexosaminidase provides further insight into GH20 family catalysis and inhibition. Biochemistry 2014, 53, 1789–1800. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, J.; Liu, M.; Yang, W.; Wang, Y.; Tang, R.; Zhang, M. Crystallographic evidence for substrate-assisted catalysis of β-N-acetylhexosaminidas from Akkermansia muciniphila. Biochem. Biophys. Res. Commun. 2019, 511, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Lenart, A.; Dudkiewicz, M.; Grynberg, M.; Pawłowski, K. CLCAs—A family of metalloproteases of intriguing phylogenetic distribution and with cases of substituted catalytic sites. PLoS ONE 2013, 8, e62272. [Google Scholar] [CrossRef]
- Val-Cid, C.; Biarnés, X.; Faijes, M.; Planas, A. Structural-functional analysis reveals a specific domain organization in family GH20 hexosaminidases. PLoS ONE 2015, 10, e0128075. [Google Scholar] [CrossRef]
- Tews, I.; Perrakis, A.; Wilson, K.S.; Oppenheim, A.; Dauter, Z.; Vorgias, C.E.; Tews, I. Bacterial chitobiase structure provides insight into catalytic mechanism and the basis of Tay–Sachs disease. Nat. Struct. Biol. 1996, 3, 638–648. [Google Scholar] [CrossRef]
- Verma, S.C.; Mahadevan, S. The ChbG gene of the chitobiose (chb) operon of Escherichia coli encodes a chitooligosaccharide deacetylase. J. Bacteriol. 2012, 194, 4959–4971. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xi, H.; Bi, Q.; Hu, Y.; Zhang, Y.; Ni, M. Cloning, expression and characterization of D-aminoacylase from Achromobacter xylosoxidans subsp. denitrificans ATCC 15173. Microbiol. Res. 2013, 168, 360–366. [Google Scholar] [CrossRef]
- Graham, T.R.; Zassenhaus, H.P.; Kaplan, A. Molecular cloning of the cDNA which encodes β-N-acetylhexosaminidase A from Dictyostelium discoideum. Complete amino acid sequence and homology with the human sequence. J. Biol. Chem. 1988, 263, 16823–16829. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studier, F.W. Protein production by auto-induction in high-density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Yem, D.W.; Wu, H.C. Purification and properties of beta-N-acetylglucosaminidase from Escherichia coli. J. Bacteriol. 1976, 125, 324–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulton, J.E.; Thomas, M.A.; Ottwell, K.K.; McCormick, S.J. Partial purification and characterization of a β-N-acetylhexosaminidase from black cherry (Prunus serotina EHRH.) seeds. Plant Sci. 1985, 42, 107–114. [Google Scholar] [CrossRef]
- Vehpoorte, J.A. Purification of two β-N-acetyl-d-glucosaminidases from beef spleen. J. Biol. Chem. 1972, 247, 4787–4793. [Google Scholar]
- Mitchell, E.D.; Houston, W.C.; Latimer, S.B. Purification and properties of a β-N-acetylaminoglucohydrolase from malted barley. Phytochemistry 1976, 15, 1869–1871. [Google Scholar] [CrossRef]
- Trimble, R.B.; Evans, G.; Maley, F. Purification and properties of endo-beta-N-acetylglucosaminidase L from Streptomyces plicatus. J. Biol. Chem. 1979, 254, 9708–9713. [Google Scholar]
- Lisboa De Marco, J.; Valadares-Inglis, M.C.; Felix, C.R. Purification and characterization of an N-acetylglucosaminidase produced by a Trichoderma harzianum strain which controls Crinipellis perniciosa. Appl. Microbiol. Biotechnol. 2004, 64, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Chen, X.-Z.; Qin, L.-N.; Tao, Y.; Dong, Z.-Y. Characterization and homologous overexpression of an N-acetylglucosaminidase Nag1 from Trichoderma reesei. Biochem. Biophys. Res. Commun. 2015, 459, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, I.; Kim, S.; Yamamoto, Y.; Ajisaka, K.; Maruyama, J.; Nakajima, H.; Kitamoto, K. Cloning and overexpression of beta-N-acetylglucosaminidase encoding gene nagA from Aspergillus oryzae and enzyme-catalyzed synthesis of human milk oligosaccharide. Biosci. Biotechnol. Biochem. 2003, 67, 646–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aunpad, R.; Rice, D.W.; Sedelnikova, S.; Panbangred, W. Biochemical characterisation of two forms of halo- and thermo-tolerant chitinase C of Salinivibrio costicola expressed in Escherichia coli. Ann. Microbiol. 2007, 57, 249–257. [Google Scholar] [CrossRef]
- Kurakake, M.; Goto, T.; Ashiki, K.; Suenaga, Y.; Komaki, T. Synthesis of new glycosides by transglycosylation of N-acetylhexosaminidase from Serratia marcescens YS-1. J. Agric. Food Chem. 2003, 51, 1701–1705. [Google Scholar] [CrossRef]
- Flockenhaus, B.; Kieß, M.; Müller, M.C.M.; Leippe, M.; Scholze, H.; Riekenberg, S.; Vahrmann, A. The β-N-acetylhexosaminidase of Entamoeba histolytica is composed of two homologous chains and has been localized to cytoplasmic granules. Mol. Biochem. Parasitol. 2004, 138, 217–225. [Google Scholar]
- Dimond, R.L.; Loomis, W.F. Vegetative isozyme of N-acetylglucosaminidase in Dictyostelium discoideum. J. Biol. Chem. 1974, 249, 5628–5632. [Google Scholar]
- Drouillard, S.; Armand, S.; Davies, J.G.; Vorgias, E.C.; Henrissat, B. Serratia marcescens chitobiase is a retaining glycosidase utilizing substrate acetamido group participation. Biochem. J. 1997, 328, 945–949. [Google Scholar] [CrossRef] [Green Version]
- Kobata, A. Structures and application of oligosaccharides in human milk. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 731–747. [Google Scholar] [CrossRef] [Green Version]
- André-Miral, C.; Koné, F.M.; Solleux, C.; Grandjean, C.; Dion, M.; Tran, V.; Tellier, C. De novo design of a trans-β-N-acetylglucosaminidase activity from a GH1 β-glycosidase by mechanism engineering. Glycobiology 2015, 25, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.A.; Nguyen, T.; K Křen, V.; Eijsink, V.G.H.; Haltrich, D.; Peterbauer, C.K. Heterologous expression and characterization of an N-acetyl-β-d-hexosaminidase from Lactococcus lactis ssp. lactis IL1403. J. Agric. Food Chem. 2012, 60, 3275–3281. [Google Scholar] [CrossRef] [PubMed]
- Rauvolfová, J.; Kuzma, M.; Weignerová, L.; Fialová, P.; Přikrylová, V.; Pišvejcová, A.; Macková, M.; Křen, V. β-N-Acetylhexosaminidase-catalysed synthesis of non-reducing oligosaccharides. J. Mol. Catal. B Enzym. 2004, 29, 233–239. [Google Scholar] [CrossRef]
- Sidorczyk, Z.; Senchenkova, S.N.; Perepelov, A.V.; Kondakova, A.N.; Kaca, W.; Rozalski, A.; Knirel, Y.A. Structure and serology of O-antigens as the basis for classification of Proteus strains. Innate Immun. 2010, 17, 70–96. [Google Scholar]
- Katzenellenbogen, E.; Kocharova, N.A.; Korzeniowska-Kowal, A.; Bogulska, M.; Rybka, J.; Gamian, A.; Kachala, V.V.; Shashkov, A.S.; Knirel, Y.A. Structure of the glycerol phosphate-containing O-specific polysaccharide and serological studies on the lipopolysaccharides of Citrobacter werkmanii PCM 1548 and PCM 1549 (serogroup O14). FEMS Immunol. Med. Microbiol. 2008, 54, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzenellenbogen, E.; Kocharova, N.A.; Zatonsky, G.V.; Shashkov, A.S.; Bogulska, M.; Knirel, Y.A. Structures of the biological repeating units in the O-chain polysaccharides of Hafnia alvei strains having a typical lipopolysaccharide outer core region. FEMS Immunol. Med. Microbiol. 2005, 45, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Barbeyron, T.; Michel, G. Evaluation of reference genes for real-time quantitative PCR in the marine flavobacterium Zobellia galactanivorans. J. Microbiol. Methods 2011, 84, 61–66. [Google Scholar] [CrossRef]
- Ashida, H.; Miyake, A.; Kiyohara, M.; Yoshida, E.; Kumagai, H.; Yamamoto, K. Two distinct α-L -fucosidases from Bifidobacterium bifidum are essential for the utilization of fucosylated milk oligosaccharides and glycoconjugates. Glycobiology 2009, 19, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Curiel, J.A.; De Las Rivas, B.; Mancheño, J.M.; Muñoz, R. The pURI family of expression vectors: A versatile set of ligation independent cloning plasmids for producing recombinant His-fusion proteins. Protein Expr. Purif. 2011, 76, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Stingele, F.; Newell, J.W.; Neeser, J.R. Unraveling the function of glycosyltransferases in Streptococcus thermophilus Sfi6. J. Bacteriol. 1999, 181, 6354–6360. [Google Scholar] [CrossRef] [Green Version]
- Viigand, K.; Visnapuu, T.; Mardo, K.; Aasamets, A.; Alamäe, T. Maltase protein of Ogataea (Hansenula) polymorpha is a counterpart to the resurrected ancestor protein ancMALS of yeast maltases and isomaltases. Yeast 2016, 33, 415–432. [Google Scholar] [CrossRef] [Green Version]
- Reiffová, K.; Nemcová, R. Thin-layer chromatography analysis of fructooligosaccharides in biological samples. J. Chromatogr. A 2006, 1110, 214–221. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; McGeough, P. A Comparison of polysaccharide substrates and reducing sugar methods for the measurement of endo-1,4-β-xylanase. Appl. Biochem. Biotechnol. 2015, 177, 1152–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | KM (mM) | Vmax (µmoL·mg−1·min−1) | kcat (s−1) | kcat/KM (mM−1·s−1) |
|---|---|---|---|---|---|
| PhNah20A | pNPGlcNAc | 0.43 ± 0.07 | 93.7 ± 5.0 | 146.8 | 341 |
| pNPGalNAc | 0.56 ± 0.11 | 123.0 ± 7.0 | 192.7 | 344 | |
| SmChb 1 | pNPGlcNAc | 56.7 ± 4.3 | NI | 111.0 | 1.95 |
| AhNag20A 2 | pNPGlcNAc | 0.52 | 115 | NI | NI |
| pNPGalNAc | 0.5 | 7.6 | NI | NI | |
| AhNagB 2 | pNPGlcNAc | 8.57 | 25 | NI | NI |
| pNPGalNAc | 11.1 | 11 | NI | NI | |
| BbhI of Bb 3 | pNPGlcNAc | 120.0 ± 0.2 | NI | 213 | 178 |
| pNPGalNAc | NA | NA | NA | NA | |
| CfHex20 4 | pNPGlcNAc | 0.053 | NI | 482.3 | 9090 |
| pNPGalNAc | 0.066 | NI | 129.1 | 1950 | |
| VfExoI 5 | pNPGlcNAc | 0.09 | 270 | NI | NI |
| pNPGalNAc | 0.33 | 130 | NI | NI | |
| Hex2 6 | pNPGlcNAc | 0.48 | NI | 60.0 ± 1.7 | NI |
| EhHexA 7 | pNPGlcNAc | 0.1 | 3.8 | NI | NI |
| TrNag1 | pNPGlcNAc | 69.4 ± 4.0 | NI | NI | 1023 ± 23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visnapuu, T.; Teze, D.; Kjeldsen, C.; Lie, A.; Duus, J.Ø.; André-Miral, C.; Pedersen, L.H.; Stougaard, P.; Svensson, B. Identification and Characterization of a β-N-Acetylhexosaminidase with a Biosynthetic Activity from the Marine Bacterium Paraglaciecola hydrolytica S66T. Int. J. Mol. Sci. 2020, 21, 417. https://doi.org/10.3390/ijms21020417
Visnapuu T, Teze D, Kjeldsen C, Lie A, Duus JØ, André-Miral C, Pedersen LH, Stougaard P, Svensson B. Identification and Characterization of a β-N-Acetylhexosaminidase with a Biosynthetic Activity from the Marine Bacterium Paraglaciecola hydrolytica S66T. International Journal of Molecular Sciences. 2020; 21(2):417. https://doi.org/10.3390/ijms21020417
Chicago/Turabian StyleVisnapuu, Triinu, David Teze, Christian Kjeldsen, Aleksander Lie, Jens Øllgaard Duus, Corinne André-Miral, Lars Haastrup Pedersen, Peter Stougaard, and Birte Svensson. 2020. "Identification and Characterization of a β-N-Acetylhexosaminidase with a Biosynthetic Activity from the Marine Bacterium Paraglaciecola hydrolytica S66T" International Journal of Molecular Sciences 21, no. 2: 417. https://doi.org/10.3390/ijms21020417