Dysregulation of Key Proteins Associated with Sperm Motility and Fertility Potential in Cancer Patients



Abstract
:1. Introduction
2. Results
2.1. Semen Parameters
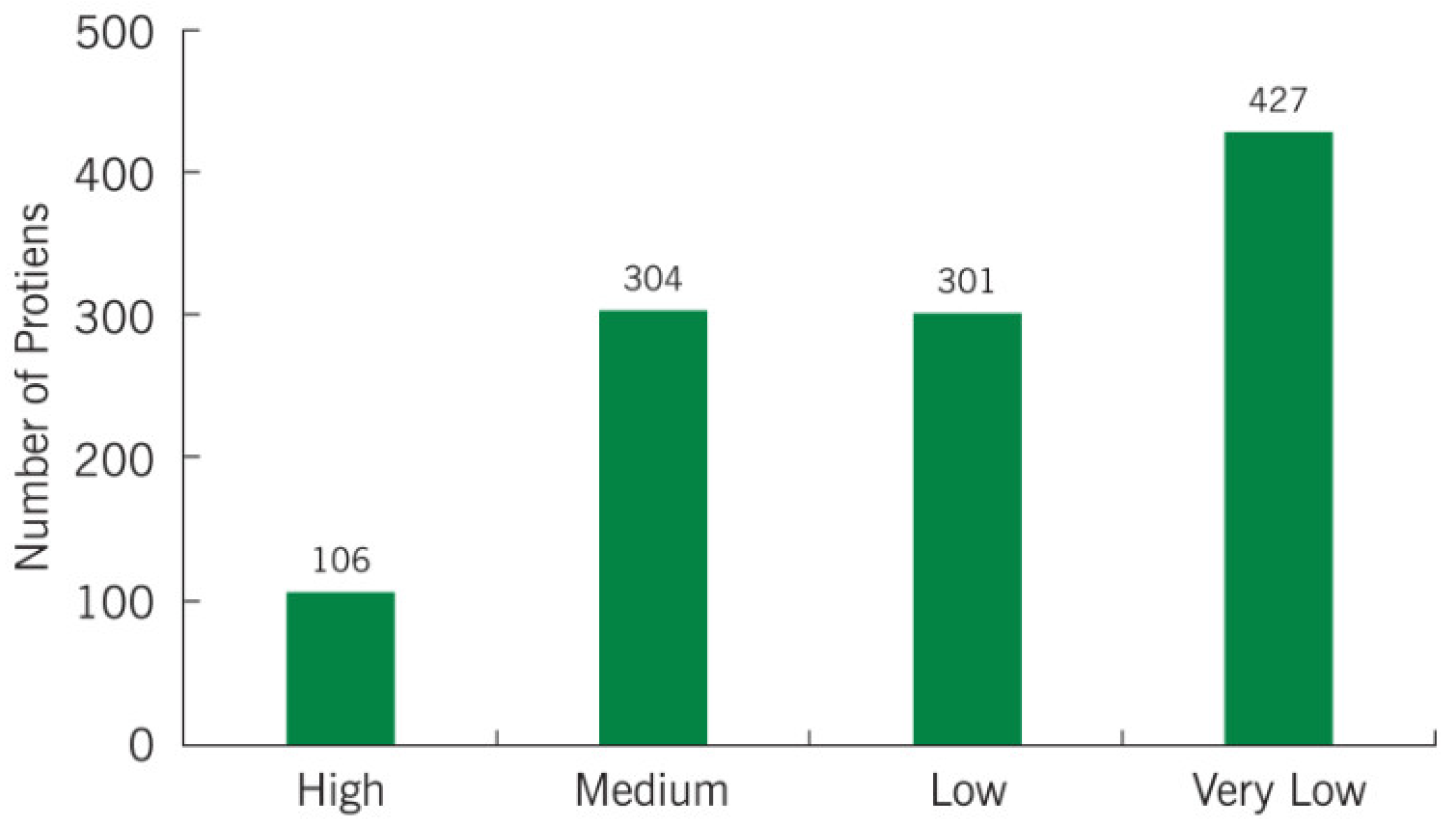
2.2. Protein Profile of Cancer Patients
2.3. Canonical Pathways
2.4. Reproductive Pathways Dysregulated in Cancer Patients
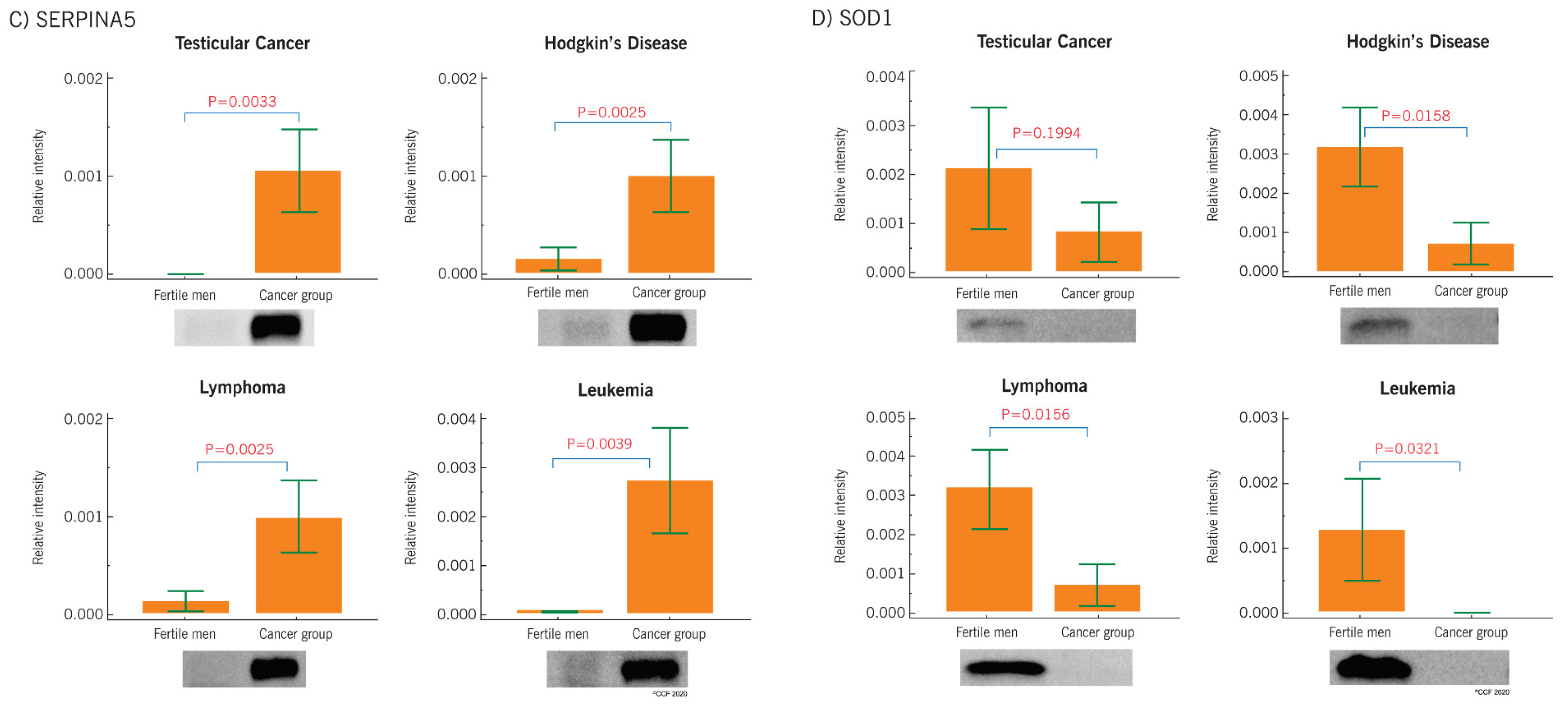
2.5. Western Blot Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Semen Analysis
4.3. Cryopreservation and Thawing
4.4. Sperm Protein Extraction, Quantification, and Separation
4.5. Liquid Chromatography-Tandem Mass Spectrometer Analysis (LC-MS/MS)
4.6. Proteomic Data Analysis
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DEPs | Differentially expressed proteins |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometer analysis |
| NSAF | Normalized spectral abundance factor |
| SpC | Spectral counts |
| TCA | Tricarboxylic acid cycle |
References
- American Cancer Society. Cancer Facts & Figures 2020; American Cancer Society: Atlanta, Georgia, 2020. [Google Scholar]
- Salati, M.; Cesaretti, M.; Macchia, M.; El Mistiri, M.; Federico, M. Epidemiological overview of Hodgkin lymphoma across the Mediterranean basin. Mediterr. J. Hematol. Infect. Dis. 2014, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Allamaneni, S.S.R. Disruption of spermatogenesis by the cancer disease process. J. Natl. Cancer Inst. Monogr. 2005, 2005, 9–12. [Google Scholar] [CrossRef]
- Osterberg, E.C.; Ramasamy, R.; Masson, P.; Brannigan, R.E. Current practices in fertility preservation in male cancer patients. Urol. Ann. 2014, 6, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Arnon, J.; Meirow, D.; Lewis-Roness, H.; Ornoy, A. Genetic and teratogenic effects of cancer treatments on gametes and embryos. Hum. Reprod. Update 2001, 7, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.J.; Shalet, S.M. Spermatogenesis after cancer treatment: Damage and recovery. J. Natl. Cancer Inst. Monogr. 2005, 2005, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.H., IV; Karpman, E.; Sander, J.C.; Spiess, P.E.; Pisters, L.L.; Lipshultz, L.I. Pretreatment semen parameters in men with cancer. J. Urol. 2009, 181, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Gandini, L.; Lombardo, F.; Salacone, P.; Paoli, D.; Anselmo, A.P.; Culasso, F.; Dondero, F.; Lenzi, A. Testicular cancer and Hodgkin’s disease: Evaluation of semen quality. Hum. Reprod. 2003, 18, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Hallak, J.; Mahran, A.; Chae, J.; Agarwal, A. Poor semen quality from patients with malignancies does not rule out sperm banking. Urol. Res. 2000, 28, 281–284. [Google Scholar] [CrossRef]
- Auger, J.; Sermondade, N.; Eustache, F. Semen quality of 4480 young cancer and systemic disease patients: Baseline data and clinical considerations. Basic Clin. Androl. 2016, 26, 3. [Google Scholar] [CrossRef] [Green Version]
- Hamano, I.; Hatakeyama, S.; Nakamura, R.; Fukuhara, R.; Noro, D.; Tanaka, T.; Yoneyama, T.; Yamamoto, H.; Yoneyama, T.; Hashimoto, Y.; et al. Differences in semen characteristics between patients with testicular cancer and other malignancies using various cut-off values. Int. J. Urol. 2018, 25, 817–824. [Google Scholar] [CrossRef]
- Bussen, S.; Sütterlin, M.; Steck, T.; Dietl, J. Semen parameters in patients, with unilateral testicular cancer compared to patients with other malignancies. Arch. Gynecol. Obstet. 2004, 269, 196–198. [Google Scholar] [CrossRef] [PubMed]
- MacKenna, A.; Crosby, J.; Huidobro, C.; Correa, E.; Duque, G. Semen quality before cryopreservation and after thawing in 543 patients with testicular cancer. JBRA Assist. Reprod. 2017, 21, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Hallak, J.; Kolettis, P.N.; Sekhon, V.S.; Thomas, A.J.; Agarwal, A. Cryopreservation of sperm from patients with leukemia: Is it worth the effort? Cancer 1999, 85, 1973–1978. [Google Scholar] [CrossRef]
- Botchan, A.; Hauser, R.; Gamzu, R.; Yogev, L.; Lessing, J.B.; Paz, G.; Yavetz, H. Sperm quality in Hodgkin’s disease versus non-Hodgkin’s lymphoma. Hum. Reprod. 1997, 12, 73–76. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.; Irani, J.; Knee, G.; Efymow, B.; Blasco, L.; Patrizio, P. Sperm cryopreservation for male patients with cancer: An epidemiological analysis at the University of Pennsylvania. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 113, S7–S11. [Google Scholar] [CrossRef]
- Paoli, D.; Rizzo, F.; Fiore, G.; Pallotti, F.; Pulsoni, A.; Annechini, G.; Lombardo, F.; Lenzi, A.; Gandini, L. Spermatogenesis in Hodgkin’s Lymphoma Patients: A Retrospective Study of Semen Quality before and after Different Chemotherapy Regimens. Hum. Reprod. 2016, 31, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Larson, K.; Sharma, R.K.; Nelson, D.R.; Evenson, D.P.; Toma, H.; Thomas, A.J.; Agarwal, A. DNA damage in patients with untreated cancer as measured by the sperm chromatin structure assay. Fertil. Steril. 2001, 75, 469–475. [Google Scholar] [CrossRef]
- Milardi, D.; Grande, G.; Vincenzoni, F.; Castagnola, M.; Marana, R. Proteomics of human seminal plasma: Identification of biomarker candidates for fertility and infertility and the evolution of technology. Mol. Reprod. Dev. 2013, 80, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Selvam, P.; Henkel, R.; Finelli, R.; Agarwal, A. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2020, e13711. [Google Scholar] [CrossRef]
- Dias, T.R.; Agarwal, A.; Pushparaj, P.N.; Ahmad, G.; Sharma, R. New insights on the mechanisms affecting fertility in men with non-seminoma testicular cancer before cancer therapy. World J. Mens. Health 2018, 36. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Agarwal, A.; Pushparaj, P.N. Altered molecular pathways in the proteome of cryopreserved sperm in testicular cancer patients before treatment. Int. J. Mol. Sci. 2019, 20, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, A.D.; Agarwal, A.; Baskaran, S.; Pushparaj, P.N.; Ahmad, G.; Panner Selvam, M.K. Alterations of spermatozoa proteomic profile in men with Hodgkin’s disease prior to cancer therapy. World J. Mens. Health 2019, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, P.; Ojha, S.K.; Kumar, V.; Bakhshi, S.; Singh, S.; Yadav, S. Differential seminal plasma proteome signatures of acute lymphoblastic leukemia survivors. Reprod. Biol. 2019, 19, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.; Schiffer, E.; von Wilcke, P.; Bauer, H.W.; Leung, H.; Siwy, J.; Ulrici, W.; Paasch, U.; Horn, L.C.; Stolzenburg, J.U. Seminal plasma as a source of prostate cancer peptide biomarker candidates for detection of indolent and advanced disease. PLoS ONE 2013, 8, e67514. [Google Scholar] [CrossRef] [PubMed]
- Veveris-Lowe, T.L.; Kruger, S.J.; Walsh, T.; Gardiner, R.A.; Clements, J.A. Seminal fluid characterization for male fertility and prostate cancer: Kallikrein-related serine proteases and whole proteome approaches. Semin. Thromb. Hemost. 2007, 33, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Drabovich, A.P.; Saraon, P.; Drabovich, M.; Karakosta, T.D.; Dimitromanolakis, A.; Eric Hyndman, M.; Jarvi, K.; Diamandis, E.P. Multi-omics biomarker pipeline reveals elevated levels of protein-glutamine gammaglutamyltransferase 4 in seminal plasma of prostate cancer patients. Mol. Cell. Proteom. 2019, 18, 1807–1823. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Agarwal, A.; Pushparaj, P.N. A quantitative global proteomics approach to understanding the functional pathways dysregulated in the spermatozoa of asthenozoospermic testicular cancer patients. Andrology 2019, 7, 454–462. [Google Scholar] [CrossRef]
- Buchman-Shaked, O.; Kraiem, Z.; Gonen, Y.; Goldman, S. Presence of matrix metalloproteinases and tissue inhibitor of matrix metalloproteinase in human sperm. J. Androl. 2002, 23, 702–708. [Google Scholar]
- Hassan, R.; Thomas, A.; Alewine, C.; Le, D.T.; Jaffee, E.M.; Pastan, I. Mesothelin immunotherapy for cancer: Ready for prime time? J. Clin. Oncol. 2016, 34, 4171–4179. [Google Scholar] [CrossRef] [Green Version]
- Bera, T.K.; Pastan, I. Mesothelin Is Not Required for Normal Mouse Development or Reproduction. Mol. Cell. Biol. 2000, 20, 2902–2906. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Conejero, J.A.; Garrido, N.; Remohí, J.; Pellicer, A.; Simón, C.; Meseguer, M. MUC1 in human testis and ejaculated spermatozoa and its relationship to male fertility status. Fertil. Steril. 2008, 90, 450–452. [Google Scholar] [CrossRef]
- Kim, D.H.; Shin, N.; Kim, G.H.; Song, G.A.; Jeon, T.Y.; Kim, D.H.; Lauwers, G.Y.; Park, D.Y. Mucin expression in gastric cancer: Reappraisal of its clinicopathologic and prognostic significance. Arch. Pathol. Lab. Med. 2013, 137, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Betge, J.; Schneider, N.I.; Harbaum, L.; Pollheimer, M.J.; Lindtner, R.A.; Kornprat, P.; Ebert, M.P.; Langner, C. MUC1, MUC2, MUC5AC, and MUC6 in colorectal cancer: Expression profiles and clinical significance. Virchows Arch. 2016, 469, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, A.Y.; Hutter, C.M.; Trent, J.M.; Meltzer, P.S. Complete sequence analysis of a gene (OS-9) ubiquitously expressed in human tissues and amplified in sarcomas. Mol. Carcinog. 1996, 15, 270–275. [Google Scholar] [CrossRef]
- Alcock, F.; Swanton, E. Mammalian OS-9 Is upregulated in response to endoplasmic reticulum stress and facilitates ubiquitination of misfolded glycoproteins. J. Mol. Biol. 2009, 385, 1032–1042. [Google Scholar] [CrossRef]
- Rajamanickam, G.D.; Kastelic, J.P.; Thundathil, J.C. Testis-specific isoform of Na/K-ATPase (ATP1A4) interactome in raft and non-raft membrane fractions from capacitated bovine sperm. Int. J. Mol. Sci. 2019, 20, 3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saaber, F.; Chen, Y.; Cui, T.; Yang, L.; Mireskandari, M.; Petersen, I. Expression of desmogleins 1–3 and their clinical impacts on human lung cancer. Pathol. Res. Pract. 2015, 211, 208–213. [Google Scholar] [CrossRef]
- Woong-Shick, A.; Sung-Pil, P.; Su-Mi, B.; Joon-Mo, L.; Sung-Eun, N.; Gye-Hyun, N.; Young-Lae, C.; Ho-Sun, C.; Heung-Jae, J.; Chong-Kook, K.; et al. Identification of hemoglobin-α and -β subunits as potential serum biomarkers for the diagnosis and prognosis of ovarian cancer. Cancer Sci. 2005, 96, 197–201. [Google Scholar] [CrossRef]
- Choi, J.W.; Liu, H.; Shin, D.H.; Yu, G.I.; Hwang, J.S.; Kim, E.S.; Yun, J.W. Proteomic and cytokine plasma biomarkers for predicting progression from colorectal adenoma to carcinoma in human patients. Proteomics 2013, 13, 2361–2374. [Google Scholar] [CrossRef]
- Song, Q.-B.; Hu, W.G.; Wang, P.; Yao, Y.; Zeng, H.Z. Identification of serum biomarkers for lung cancer using magnetic bead-based SELDI-TOF-MS. Acta Pharmacol. Sin. 2011, 32, 1537–1542. [Google Scholar] [CrossRef] [Green Version]
- Ivry, S.L.; Knudsen, G.M.; Caiazza, F.; Sharib, J.M.; Jaradeh, K.; Ravalin, M.; O’Donoghue, A.J.; Kirkwood, K.S.; Craik, C.S. The lysosomal aminopeptidase tripeptidyl peptidase 1 displays increased activity in malignant pancreatic cysts. Biol. Chem. 2019, 400. [Google Scholar] [CrossRef]
- Camargo, M.; Intasqui, P.; Belardin, L.B.; Antoniassi, M.P.; Cardozo, K.H.M.; Carvalho, V.M.; Fraietta, R.; Bertolla, R.P. Molecular pathways of varicocele and its repair—A paired labelled shotgun proteomics approach. J. Proteom. 2019, 196, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, D.I.; Kim, J.; Kim, B.H.; Kim, A. Hornerin is involved in breast cancer progression. J. Breast Cancer 2016, 19, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.J.; Shen, S.L.; Li, S.Q.; Hua, Y.P.; Hu, W.J.; Guo, B.C.; Peng, B.G. Hornerin promotes tumor progression and is associated with poor prognosis in hepatocellular carcinoma. BMC Cancer 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, P.T.; Da Silva, B.F.; Lo Turco, E.G.; Fraietta, R.; Spaine, D.M.; Santos, L.F.A.; Pilau, E.J.; Gozzo, F.C.; Cedenho, A.P.; Bertolla, R.P. Changes in the seminal plasma proteome of adolescents before and after varicocelectomy. Fertil. Steril. 2013, 100, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Poersch, A.; Grassi, M.L.; de Carvalho, V.P.; Lanfredi, G.P.; de Souza Palma, C.; Greene, L.J.; de Sousa, C.B.; Carrara, H.H.A.; Dos Reis, F.J.C.; Faça, V.M. A proteomic signature of ovarian cancer tumor fluid identified by highthroughput and verified by targeted proteomics. J. Proteom. 2016, 145, 226–236. [Google Scholar] [CrossRef] [PubMed]
- de Souza Cavalcante, M.; Torres-Romero, J.C.; Lobo, M.D.P.; Moreno, F.B.M.B.; Bezerra, L.P.; Lima, D.S.; Matos, J.C.; de Azevedo Moreira, R.; de Oliveira Monteiro-Moreira, A.C. A panel of glycoproteins as candidate biomarkers for early diagnosis and treatment evaluation of B-cell acute lymphoblastic leukemia. Biomark. Res. 2016, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Chen, Z.; Wang, C.; Xiong, Z.; Zhao, W.; Jia, C.; Lin, J.; Lin, Y.; Yuan, W.; Zhao, A.Z.; et al. Rictor regulates spermatogenesis by controlling sertoli cell cytoskeletal organization and cell polarity in the mouse testis. Endocrinology 2015, 156, 4244–4256. [Google Scholar] [CrossRef]
- Oliveira, P.F.; Cheng, C.Y.; Alves, M.G. Emerging role for mammalian target of rapamycin in male fertility. Trends Endocrinol. Metab. 2017, 28, 165–167. [Google Scholar] [CrossRef] [Green Version]
- Shukla, K.K.; Kwon, W.S.; Rahman, M.S.; Park, Y.J.; You, Y.A.; Pang, M.G. Nutlin-3a decreases male fertility via UQCRC2. PLoS ONE 2013, 8, 76959. [Google Scholar] [CrossRef] [Green Version]
- Swain, N.; Samanta, L.; Agarwal, A.; Kumar, S.; Dixit, A.; Gopalan, B.; Durairajanayagam, D.; Sharma, R.; Pushparaj, P.N.; Baskaran, S. Aberrant upregulation of compensatory redox molecular machines may contribute to sperm dysfunction in infertile men with unilateral varicocele: A proteomic insight. Antioxid. Redox Signal. 2020, 32, 504–521. [Google Scholar] [CrossRef]
- Yu, B.; Huang, Z. Variations in antioxidant genes and male infertility. Biomed. Res. Int. 2015, 2015, 513196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- España, F.; Navarro, S.; Medina, P.; Zorio, E.; Estellés, A. The role of protein C inhibitor in human reproduction. Semin. Thromb. Hemost. 2007, 33, 41–45. [Google Scholar] [CrossRef]
- Wang, S.; Wang, W.; Xu, Y.; Tang, M.; Fang, J.; Sun, H.; Sun, Y.; Gu, M.; Liu, Z.; Zhang, Z.; et al. Proteomic characteristics of human sperm cryopreservation. Proteomics 2014, 14, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Bartolomé, S.; Deutsch, E.W.; Binz, P.A.; Jones, A.R.; Eisenacher, M.; Mayer, G.; Campos, A.; Canals, F.; Bech-Serra, J.J.; Carrascal, M.; et al. Guidelines for reporting quantitative mass spectrometry based experiments in proteomics. J. Proteom. 2013, 95, 84–88. [Google Scholar] [CrossRef]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO: Geneva, Switzerland, 2010. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects | Sperm Concentration (106/mL) | Total Motility (%) |
|---|---|---|
| Fertile Donors (n = 19) | 81 (70.4–107.1) | 71 (56.5–70.4) |
| Testicular cancer (n = 40) | 16 a (9.9–29.4) | 51 a (35–67.5) |
| Hodgkin’s disease (n = 32) | 26.8 a (13–62) | 60.5 (41.0–74.0) |
| Leukemia (n = 17) | 64.7 a (43–72) | 70 (33.8–95.7) |
| Lymphoma (n = 20) | 55.9 a (42.4–64.7) | 43.5 a (24.4–55.5) |
| S.N | UniProt ID | Protein | Abundance |
|---|---|---|---|
| Cancer vs. Fertile Men | |||
| 1. | Q13421 | mesothelin isoform X1 | L |
| 2. | Q6W4 × 9 | mucin-6 (MUC6) isoform X1, partial | L |
| 3. | Q13438 | protein OS-9 isoform X1 | L |
| 4. | Q02413 | desmoglein-1 preproprotein | VL |
| 5. | P68871 | hemoglobin subunit beta | VL |
| 6. | O14773 | tripeptidyl-peptidase 1 preproprotein | VL |
| 7. | Q86YZ3 | hornerin | VL |
| 8. | P08697 | alpha-2-antiplasmin isoform X6 | VL |
| Pathways | Z Score * |
|---|---|
| Oxidative Phosphorylation | −3.46 |
| Tricarboxylic acid (TCA) cycle II | −2.45 |
| Fatty acid β-oxidation I | −2.00 |
| Glycolysis I | −2.24 |
| Upstream regulators | |
| RICTOR | 4.785 |
| KDM5A | 3.464 |
| MAP4K4 | 3.162 |
| TRAP1 | 2.433 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panner Selvam, M.K.; Finelli, R.; Baskaran, S.; Agarwal, A. Dysregulation of Key Proteins Associated with Sperm Motility and Fertility Potential in Cancer Patients. Int. J. Mol. Sci. 2020, 21, 6754. https://doi.org/10.3390/ijms21186754
Panner Selvam MK, Finelli R, Baskaran S, Agarwal A. Dysregulation of Key Proteins Associated with Sperm Motility and Fertility Potential in Cancer Patients. International Journal of Molecular Sciences. 2020; 21(18):6754. https://doi.org/10.3390/ijms21186754
Chicago/Turabian StylePanner Selvam, Manesh Kumar, Renata Finelli, Saradha Baskaran, and Ashok Agarwal. 2020. "Dysregulation of Key Proteins Associated with Sperm Motility and Fertility Potential in Cancer Patients" International Journal of Molecular Sciences 21, no. 18: 6754. https://doi.org/10.3390/ijms21186754