Novel ANO1 Inhibitor from Mallotus apelta Extract Exerts Anticancer Activity through Downregulation of ANO1


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Isolation of Compounds
2.2. Structural Elucidations of Compounds
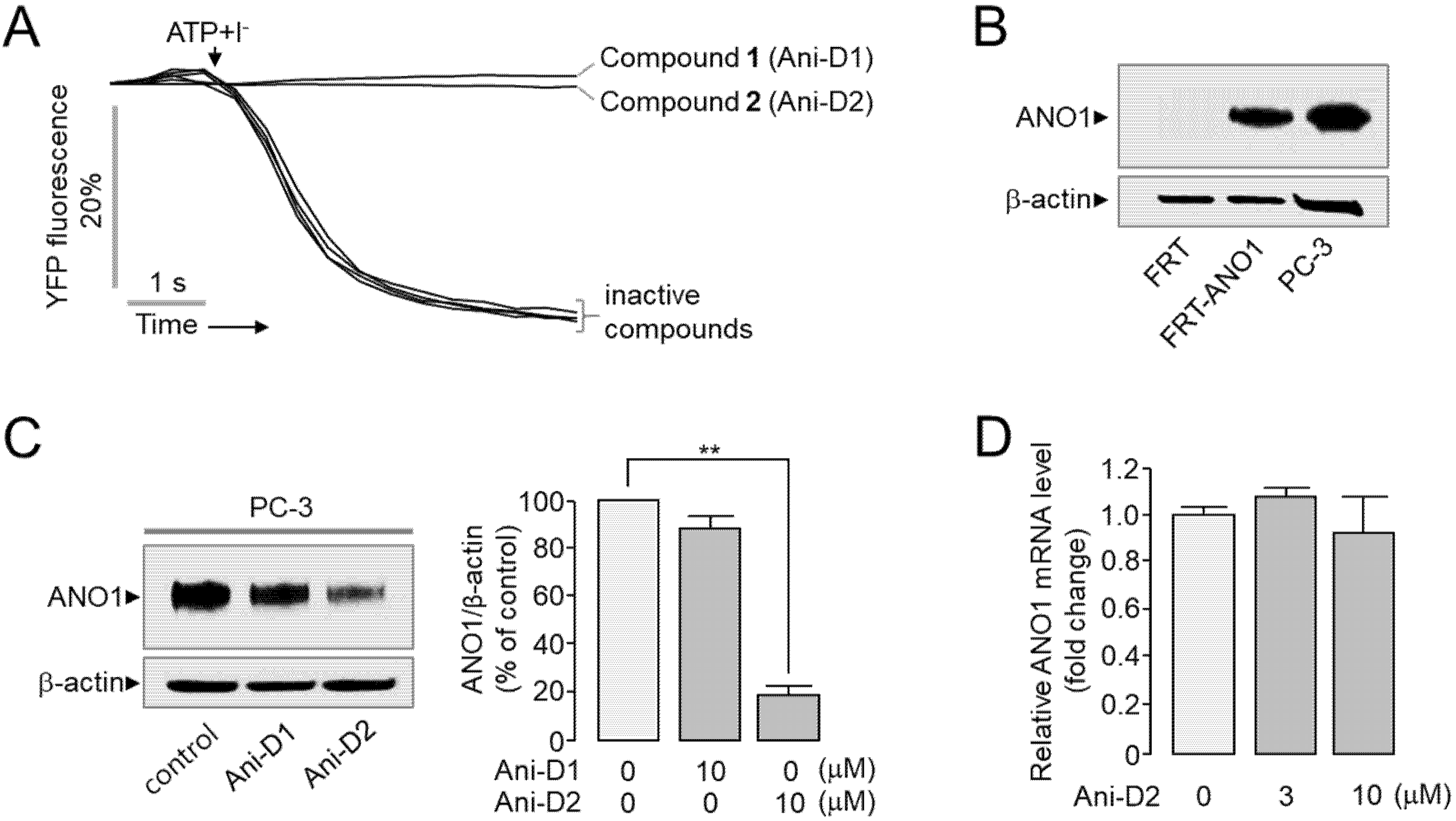
2.3. Identification and Characterization of Novel ANO1 Inhibitors
2.4. Cytotoxic Effect of Compounds on PC-3 Cells
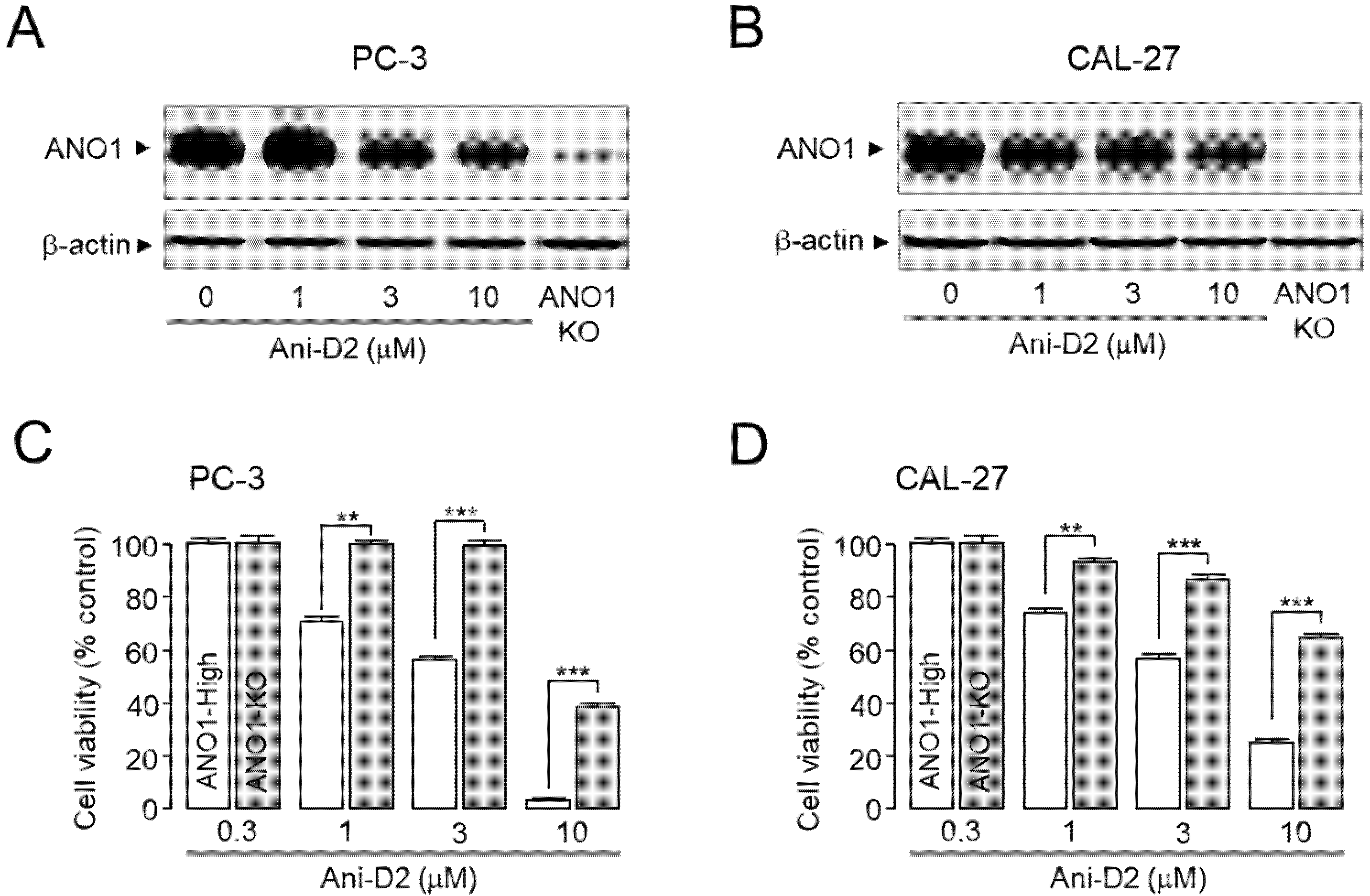
2.5. Selective Inhibition of ANO1 by Ani-D2
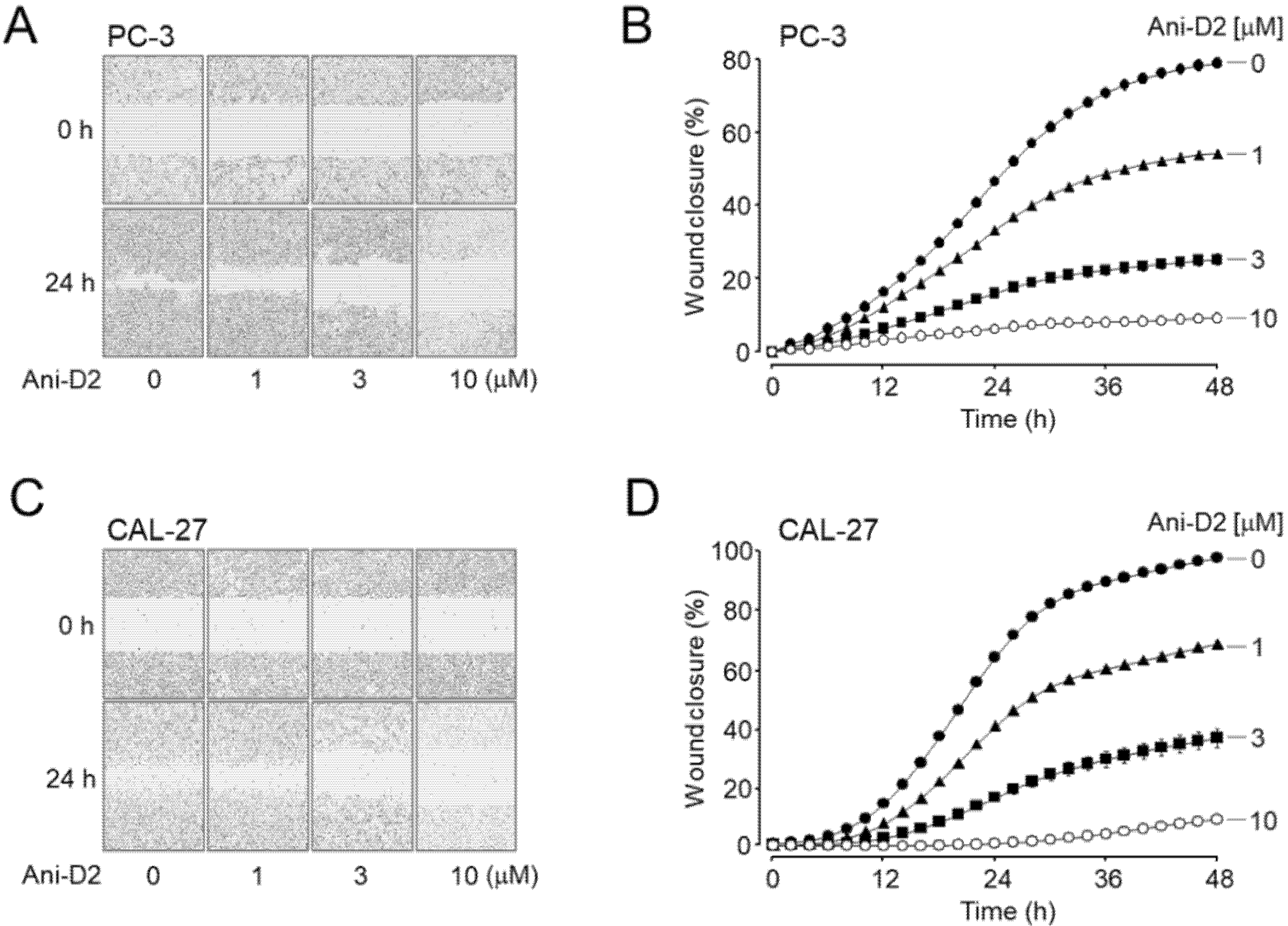
2.6. Inhibitory Effects of Ani-D2 on Cell Proliferation and Migration in PC-3 and CAL-27 Cells
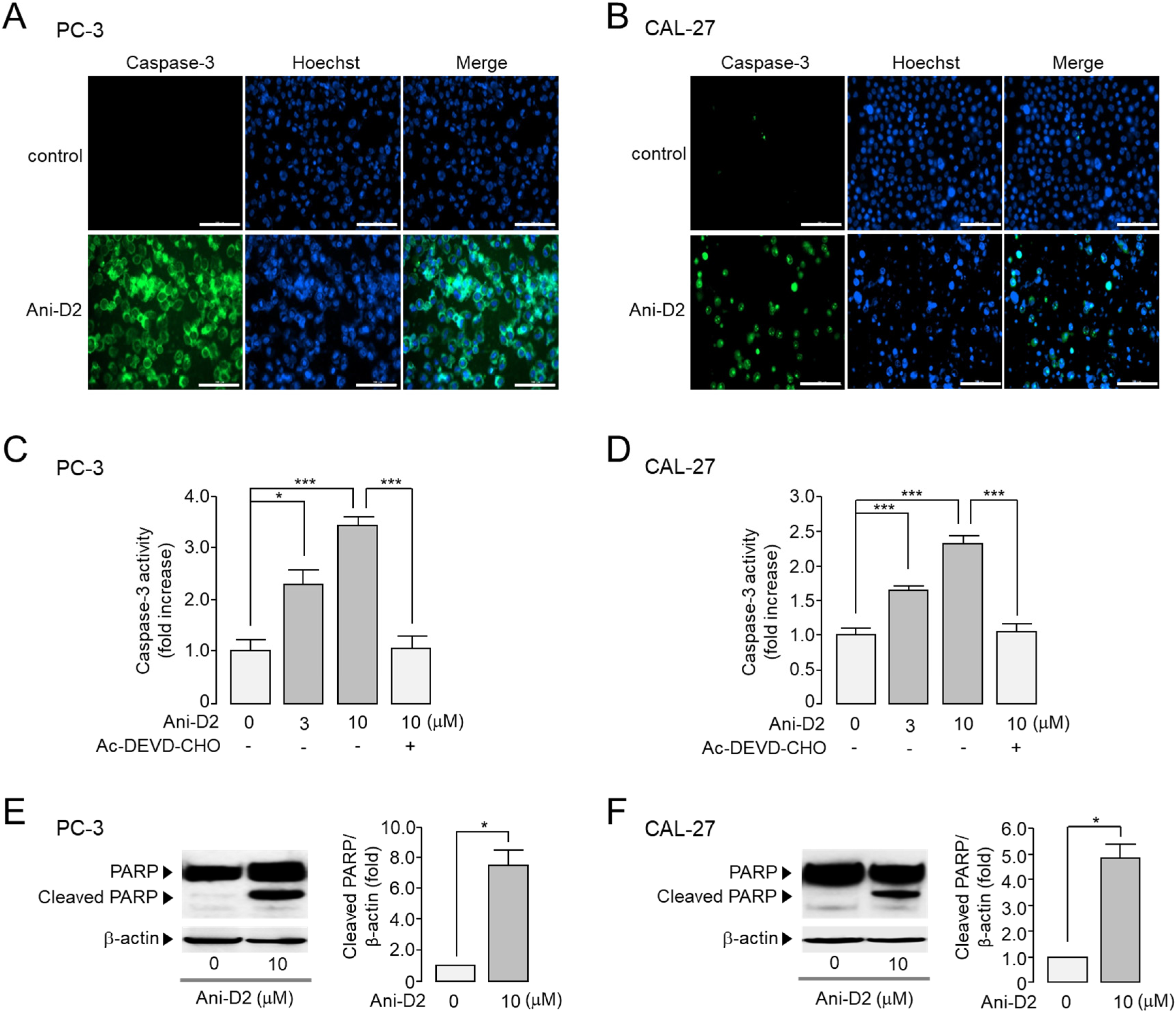
2.7. Increase in Caspase-3 Activity and PARP Cleavage by Ani-D2 in PC-3 and CAL-27 Cells
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Material
4.3. Extraction and Isolation
4.3.1. Malloapelta I (1, Ani-D1)
4.3.2. Malloapelta II (2, Ani-D2)
4.4. Cell Culture
4.5. Construction of ANO1 Knockout (KO) Cells
4.6. YFP Fluorescence Quenching Analysis
4.7. Short-Circuit Current
4.8. Intracellular Calcium Measurement
4.9. Western Blot Analysis
4.10. Real-Time RT-PCR Analysis
4.11. Cell Viability Assay
4.12. Wound Healing Assay
4.13. Caspase-3 Activity Assay
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANO1 | anoctamin1 |
| CaCC | calcium-activated chloride channels |
| CAMKII | Ca2+/calmodulin-dependent protein kinase II |
| CC | column chromatography |
| CFTR | cystic fibrosis transmembrane conductance regulator |
| FRT | Fisher rat thyroid |
| HNSCC | head and neck squamous cell carcinoma |
| HPLC | high performance liquid chromatography |
| HR-ESI-MS | high-resolution electrospray ionization mass spectrometry |
| HSQC | heteronuclear single quantum coherence |
| NMR | nuclear magnetic resonance |
| OSCC | oral squamous cell carcinoma |
| PC3 | prostate cancer cell line |
| Q-TOF LC/MS | quad time of flight liquid chromatography–mass spectrometry |
| VAST | Vietnam Academy of Science and Technology |
References
- Caputo, A.; Caci, E.; Ferrera, L.; Pedemonte, N.; Barsanti, C.; Sondo, E.; Pfeffer, U.; Ravazzolo, R.; Zegarra-Moran, O.; Galietta, L.J. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 2008, 322, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.C.; Cheng, T.; Jan, Y.N.; Jan, L.Y. Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 2008, 134, 1019–1029. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.D.; Cho, H.; Koo, J.Y.; Tak, M.H.; Cho, Y.; Shim, W.S.; Park, S.P.; Lee, J.; Lee, B.; Kim, B.M.; et al. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 2008, 455, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Cole, W.C. ANO1-ther brick in the wall—Role of Ca2+-activated Cl− channels of interstitial cells of Cajal in cholinergic motor control of gastrointestinal smooth muscle. J. Physiol. 2011, 589, 4641–4642. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Yang, Y.D.; Lee, J.; Lee, B.; Kim, T.; Jang, Y.; Back, S.K.; Na, H.S.; Harfe, B.D.; Wang, F.; et al. The calcium-activated chloride channel anoctamin 1 acts as a heat sensor in nociceptive neurons. Nat. Neurosci. 2012, 15, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lu, M.; Liu, B.; Huang, Y.; Wang, K. Inhibition of Ca2+-activated Cl− channel ANO1/TMEM16A expression suppresses tumor growth and invasiveness in human prostate carcinoma. Cancer Lett. 2012, 326, 41–51. [Google Scholar] [CrossRef]
- West, R.B.; Corless, C.L.; Chen, X.; Rubin, B.P.; Subramanian, S.; Montgomery, K.; Zhu, S.; Ball, C.A.; Nielsen, T.O.; Patel, R.; et al. The novel marker, DOG1, is expressed ubiquitously in gastrointestinal stromal tumors irrespective of KIT or PDGFRA mutation status. Am. J. Pathol. 2004, 165, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Britschgi, A.; Bill, A.; Brinkhaus, H.; Rothwell, C.; Clay, I.; Duss, S.; Rebhan, M.; Raman, P.; Guy, C.T.; Wetzel, K.; et al. Calcium-activated chloride channel ANO1 promotes breast cancer progression by activating EGFR and CAMK signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, C.; Martins, J.R.; Rudin, F.; Schneider, S.; Dietsche, T.; Fischer, C.A.; Tornillo, L.; Terracciano, L.M.; Schreiber, R.; Bubendorf, L.; et al. Enhanced expression of ANO1 in head and neck squamous cell carcinoma causes cell migration and correlates with poor prognosis. PLoS ONE 2012, 7, e43265. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, J.; Hong, S. ANO1 as a marker of oral squamous cell carcinoma and silencing ANO1 suppresses migration of human SCC-25 cells. Med. Oral Patol. Oral Cir. Bucal 2014, 19, 313–319. [Google Scholar] [CrossRef]
- Mazzone, A.; Eisenman, S.T.; Strege, P.R.; Yao, Z.; Ordog, T.; Gibbons, S.J.; Farrugia, G. Inhibition of cell proliferation by a selective inhibitor of the Ca2+-activated Cl− channel, Ano1. Biochem. Biophys. Res. Commun. 2012, 427, 248–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.; Park, J.; Kim, M.; Lee, H.K.; Kim, J.H.; Jeong, J.H.; Namkung, W. Inhibition of ANO1/TMEM16A Chloride Channel by Idebenone and Its Cytotoxicity to Cancer Cell Lines. PLoS ONE 2015, 10, e0133656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.; Kim, J.; Chang, J.; Kim, S.S.; Namkung, W.; Kim, I. Synthesis and biological evaluation of novel Ani9 derivatives as potent and selective ANO1 inhibitors. Eur. J. Med. Chem. 2018, 160, 245–255. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, R.; Namkung, W.; Mills, A.; Verkman, A.S. Small-molecule screen identifies inhibitors of a human intestinal calcium-activated chloride channel. Mol. Pharmacol. 2008, 73, 758–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fintha, A.; Gasparics, A.; Fang, L.; Erdei, Z.; Hamar, P.; Mozes, M.M.; Kokeny, G.; Rosivall, L.; Sebe, A. Characterization and role of SCAI during renal fibrosis and epithelial-to-mesenchymal transition. Am. J. Pathol. 2013, 182, 388–400. [Google Scholar] [CrossRef]
- Namkung, W.; Thiagarajah, J.R.; Phuan, P.W.; Verkman, A.S. Inhibition of Ca2+-activated Cl- channels by gallotannins as a possible molecular basis for health benefits of red wine and green tea. FASEB J. 2010, 11, 4178–4186. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Lee, H.K.; Park, J.; Jeon, D.K.; Jo, S.; Jo, M.; Namkung, W. Ani9, A Novel Potent Small-Molecule ANO1 Inhibitor with Negligible Effect on ANO2. PLoS ONE 2016, 11, e0155771. [Google Scholar] [CrossRef] [Green Version]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.; Wang, S. Malloapeltic acid, a new benzopyran derivative from Mallotus Apelta. Chem. Nat. Comput. 2010, 46, 7–9. [Google Scholar] [CrossRef]
- Xu, J.-F.; Feng, Z.-M.; Liu, J.; Zhang, P.-C. New hepatoprotective coumarinolignoids from Mallotus apelta. Chem. Biodivers. 2008, 5, 591–597. [Google Scholar] [CrossRef]
- Lu, T.; Deng, S.; Li, C.; Wu, L.; Yang, R.; Li, J. A new chromone from the twig of Mallotus apelta. Nat. Prod. Res. 2014, 28, 1864–1868. [Google Scholar] [CrossRef]
- Ma, J.; Shi, H.; Mi, C.; Li, H.L.; Lee, J.J.; Jin, X. Malloapelta B suppresses LPS-induced NF-κB activation and NF-κB-regulated target gene products. Int. Immunopharmacol. 2015, 24, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Kiem, P.V.; Dang, N.H.; Bao, H.V.; Huong, H.T.; Minh, C.V.; Huong, L.M.; Lee, J.J.; Kim, Y.H. New cytotoxic benzopyrans from the leaves of Mallotus apelta. Arch. Pharmacal Res. 2005, 28, 1131–1134. [Google Scholar] [CrossRef] [PubMed]
- Kiem, P.V.; Minh, C.V.; Huong, H.T.; Nam, N.H.; Lee, J.J.; Kim, Y.H. Chemical investigation and biological studies of Mallotus apelta III-New chromene derivative from Mallotus apelta. Vietnam J. Chem. 2005, 43, 652–656. [Google Scholar]
- Kubo, M.; Sasaki, H.; Endo, T.; Taguchi, H.; Yosioka, I. The constituents of Schizonepeta tenuifolia BRIQ. II. Structure of a new monoterpene glucoside, schizonepetoside C. Chem. Pharm. Bull. 1986, 34, 3097–3101. [Google Scholar] [CrossRef] [Green Version]
- Matsunami, K.; Otsuka, H.; Takeda, Y. Structural revisions of blumenol C glucoside and byzantionoside B. Chem. Pharm. Bull. 2010, 58, 438–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiem, P.V.; Minh, C.V.; Dat, N.T.; Cai, X.F.; Lee, J.J.; Kim, Y.H. Two new phenylpropanoid glycosides from the stem bark of Acanthopanax trifoliatus. Arch. Pharm. Res. 2003, 26, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Gao, J.; Guan, L.; Chen, X.; Gao, J.; Wang, K. Inhibition of ANO1/TMEM16A induces apoptosis in human prostate carcinoma cells by activating TNF-α signaling. Cell Death Dis. 2018, 9, 703. [Google Scholar] [CrossRef] [Green Version]
- Bill, A.; Gutierrez, A.; Kulkarni, S.; Kemp, C.; Bonenfant, D.; Voshol, H.; Duvvuri, U.; Gaither, L.A. ANO1/TMEM16A interacts with EGFR and correlates with sensitivity to EGFR-targeting therapy in head and neck cancer. Oncotarget 2015, 6, 9173–9188. [Google Scholar] [CrossRef] [Green Version]
- Duvvuri, U.; Shiwarski, D.J.; Xiao, D.; Bertrand, C.; Huang, X.; Edinger, R.S.; Rock, J.R.; Harfe, B.D.; Henson, B.J.; Kunzelmann, K.; et al. TMEM16A induces MAPK and contributes directly to tumorigenesis and cancer progression. Cancer Res. 2012, 72, 3270–3281. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zou, L.; Ma, K.; Yu, J.; Wu, H.; Wei, M.; Xiao, Q. Cell-specific mechanisms of TMEM16A Ca2+-activated chloride channel in cancer. Mol. Cancer 2017, 16, 152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lai, L.; Tan, Y.; Liang, Q.; Bai, F.; Mai, W.; Huang, Q.; Ye, Y. Hepatoprotective effect of total flavonoids of Mallotus apelta (Lour.) Muell.Arg. leaf against carbon tetrachloride-induced liver fibrosis in rats via modulation of TGF-β1/Smad and NF-κB signaling pathways. J. Ethnopharmacol. 2020, 254, 112714. [Google Scholar] [CrossRef]
- Rivière, C.; Hong, V.N.T.; Hong, Q.T.; Chataigné, G.; Hoai, N.N.; Dejaegher, B.; Tistaert, C.; Kim, T.N.T.; Vander Heyden, Y.; Van, M.C. Mallotus species from Vietnamese mountainous areas: Phytochemistry and pharmacological activities. Phytochem. Rev. 2010, 9, 217–253. [Google Scholar] [CrossRef]
- Minh, C.V.; Kiem, P.V.; Huong, H.T.; Dat, N.T.; Nam, N.H.; Lee, J.J.; Kim, Y.H. Chemical investigations and biological studies of Mallotus apelta. J. Chem. 2005, 43, 235–239. [Google Scholar]
- Seo, Y.; Ryu, K.; Park, J.; Jeon, D.K.; Jo, S.; Lee, H.K.; Namkung, W. Inhibition of ANO1 by luteolin and its cytotoxicity in human prostate cancer PC-3 cells. PLoS ONE 2017, 12, e0174935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, L.; Song, Y.; Gao, J.; Gao, J.; Wang, K. Inhibition of calcium-activated chloride channel ANO1 suppresses proliferation and induces apoptosis of epithelium originated cancer cells. Oncotarget 2016, 7, 78619–78630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galietta, L.J.; Springsteel, M.F.; Eda, M.; Niedzinski, E.J.; By, K.; Haddadin, M.J.; Kurth, M.J.; Nantz, M.H.; Verkman, A.S. Novel CFTR chloride channel activators identified by screening of combinatorial libraries based on flavone and benzoquinolizinium lead compounds. J. Biol. Chem. 2001, 276, 19723–19728. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | 1 | 2 | ||
|---|---|---|---|---|
| δC (a) | δH (a) (mult., J = Hz) | δC (b) | δH (b) (mult., J = Hz) | |
| 2 | 79.1 | - | 78.4 | - |
| 3 | 125.3 | 5.52 (d, 10.0) | 126.3 | 5.50 (d, 10.5) |
| 4 | 118.0 | 6.61 (d, 10.0) | 115.3 | 6.62 (d, 10.5) |
| 5 | 157.7 | - | 162.3 | - |
| 6 | 106.7 | - | 102.8 | - |
| 7 | 167.5 | - | 162.5 | - |
| 8 | 96.7 | 5.93 (s) | 96.3 | 5.95 (s) |
| 9 | 161.6 | - | 158.4 | - |
| 10 | 104.0 | - | 103.1 | - |
| 11 | 28.2 | 1.56 (s) | 28.5 | 1.44 (s) |
| 12 | 28.2 | 1.56 (s) | 28.4 | 1.43 (s) |
| 1′ | 194.1 | 196.1 | ||
| 2′ | 125.5 | 8.00 (d, 15.5) | 43.1 | 2.78 (dd, 3.0, 17.0) 3.07 (dd, 13.0, 17.0) |
| 3′ | 143.7 | 7.70 (d, 15.5) | 78.9 | 5.34 (dd, 3.0, 13.0) |
| 4′ | 128.4 | - | 130.5 | - |
| 5′, 9′ | 131.2 | 7.52 (d, 8.5) | 128.0 | 7.31 (d, 8.5) |
| 6′, 8′ | 117.0 | 6.86 (d, 8.5) | 115.7 | 6.87 (d, 8.5) |
| 7′ | 161.2 | - | 161.2 | - |
| Compounds | Cell Viability | IC50 (μM) |
|---|---|---|
| 1 | 30.5 ± 0.28 | 8.89 ± 0.17 |
| 2 | 12.0 ± 0.11 | 7.29 ± 0.20 |
| 3 | 1.9 ± 0.02 | 1.60 ± 0.05 |
| 4 | 78.3 ± 0.72 | >30 |
| 5 | 97.4 ± 0.90 | >30 |
| 6 | 102.5 ± 0.94 | >30 |
| Capecitabine | 27.1 ± 0.20 | 11.2 ± 1.44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, Y.; Anh, N.H.; Heo, Y.; Park, S.-H.; Kiem, P.V.; Lee, Y.; Yen, D.T.H.; Jo, S.; Jeon, D.; Tai, B.H.; et al. Novel ANO1 Inhibitor from Mallotus apelta Extract Exerts Anticancer Activity through Downregulation of ANO1. Int. J. Mol. Sci. 2020, 21, 6470. https://doi.org/10.3390/ijms21186470
Seo Y, Anh NH, Heo Y, Park S-H, Kiem PV, Lee Y, Yen DTH, Jo S, Jeon D, Tai BH, et al. Novel ANO1 Inhibitor from Mallotus apelta Extract Exerts Anticancer Activity through Downregulation of ANO1. International Journal of Molecular Sciences. 2020; 21(18):6470. https://doi.org/10.3390/ijms21186470
Chicago/Turabian StyleSeo, Yohan, Nguyen Hoang Anh, Yunkyung Heo, So-Hyeon Park, Phan Van Kiem, Yechan Lee, Duong Thi Hai Yen, Sungwoo Jo, Dongkyu Jeon, Bui Huu Tai, and et al. 2020. "Novel ANO1 Inhibitor from Mallotus apelta Extract Exerts Anticancer Activity through Downregulation of ANO1" International Journal of Molecular Sciences 21, no. 18: 6470. https://doi.org/10.3390/ijms21186470