Bordetella holmesii Lipopolysaccharide Hide and Seek Game with Pertussis: Structural Analysis of the O-Specific Polysaccharide and the Core Oligosaccharide of the Type Strain ATCC 51541


Abstract
:1. Introduction
2. Results
2.1. Isolation of Lipopolysaccharides and O-Specific Polysaccharides
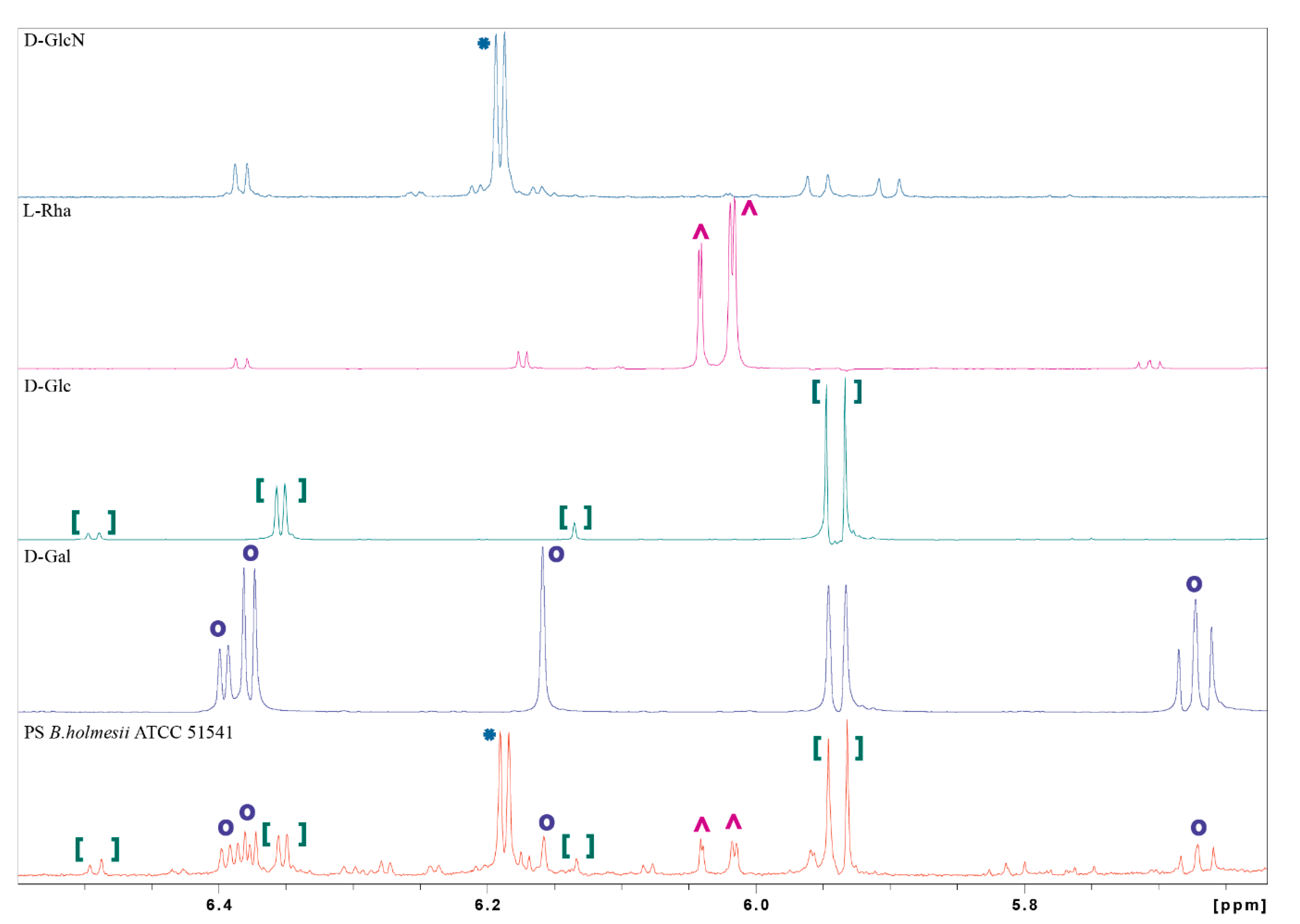
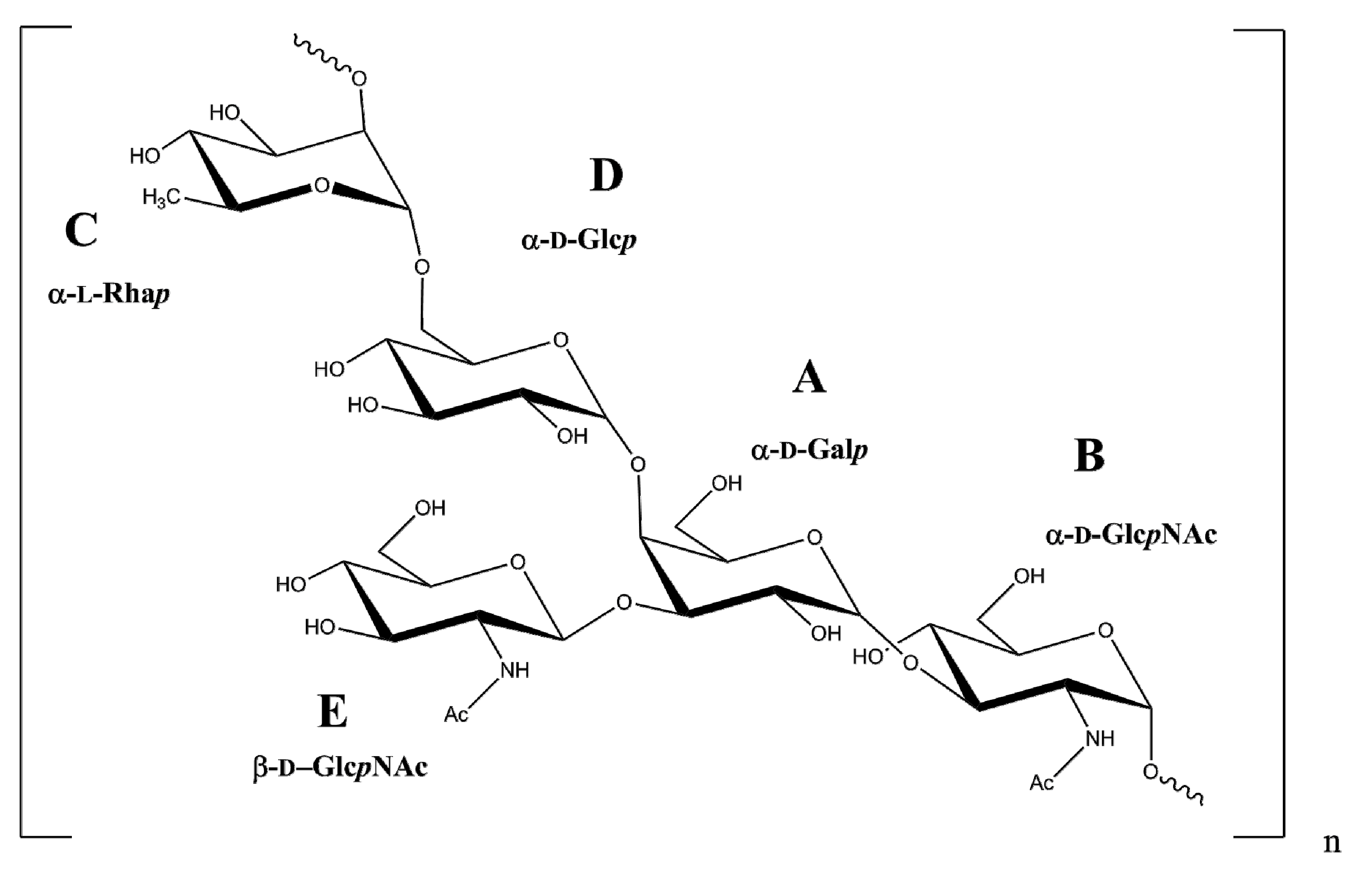
2.2. NMR Spectroscopy and Chemical Analysis of the O-PS I
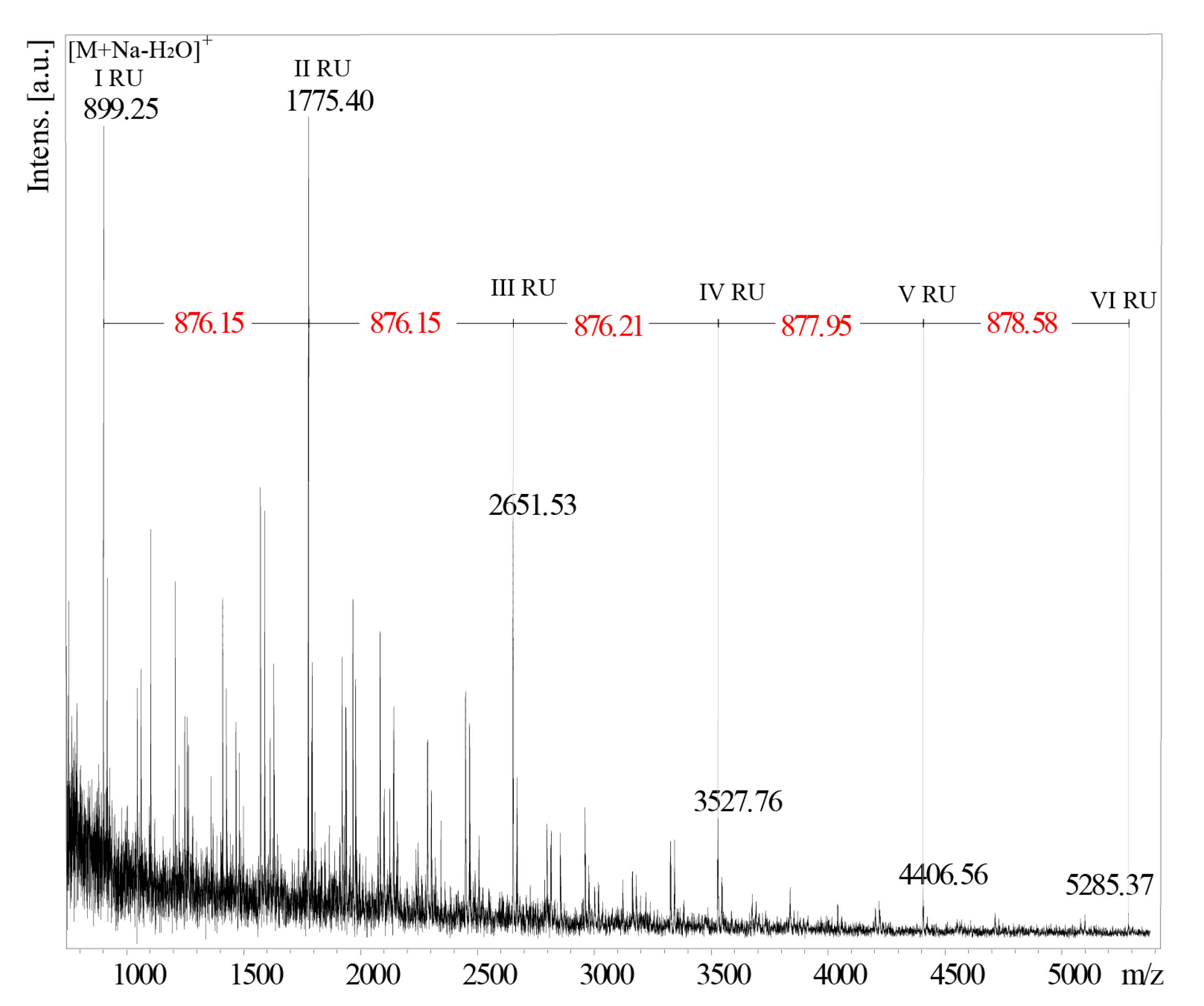
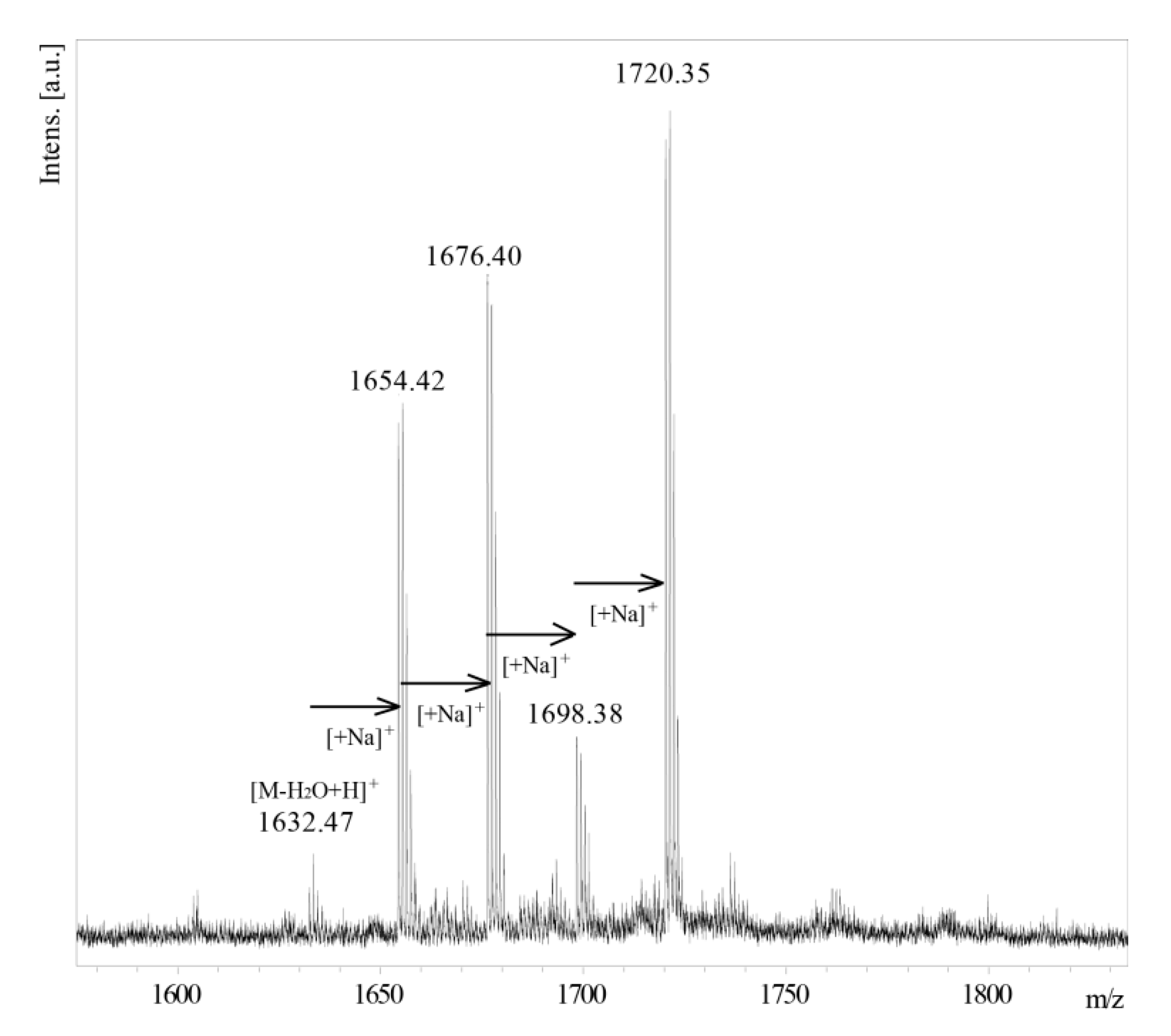
2.3. Structural Analysis of the O-PS I by Mass Spectrometry
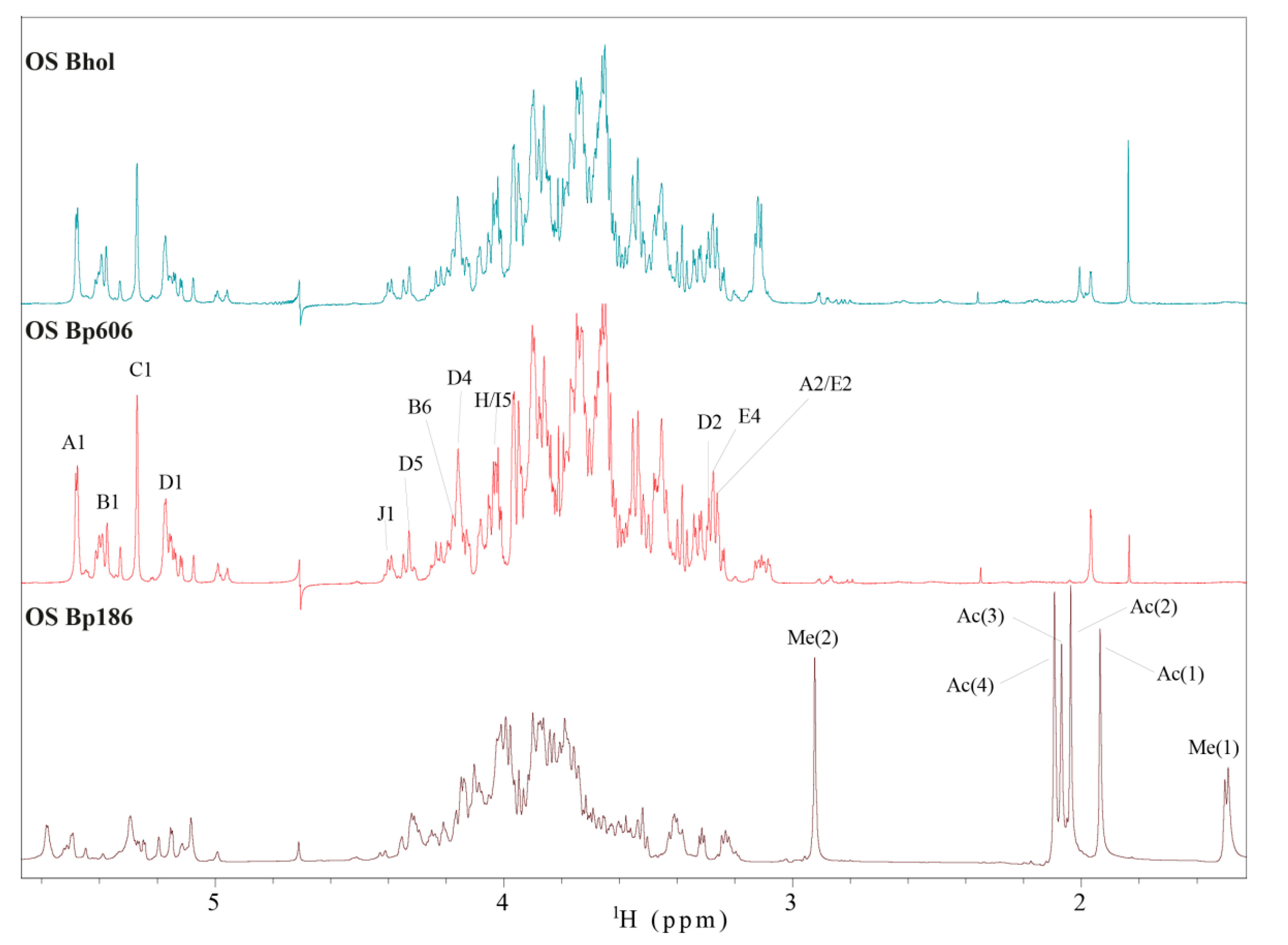
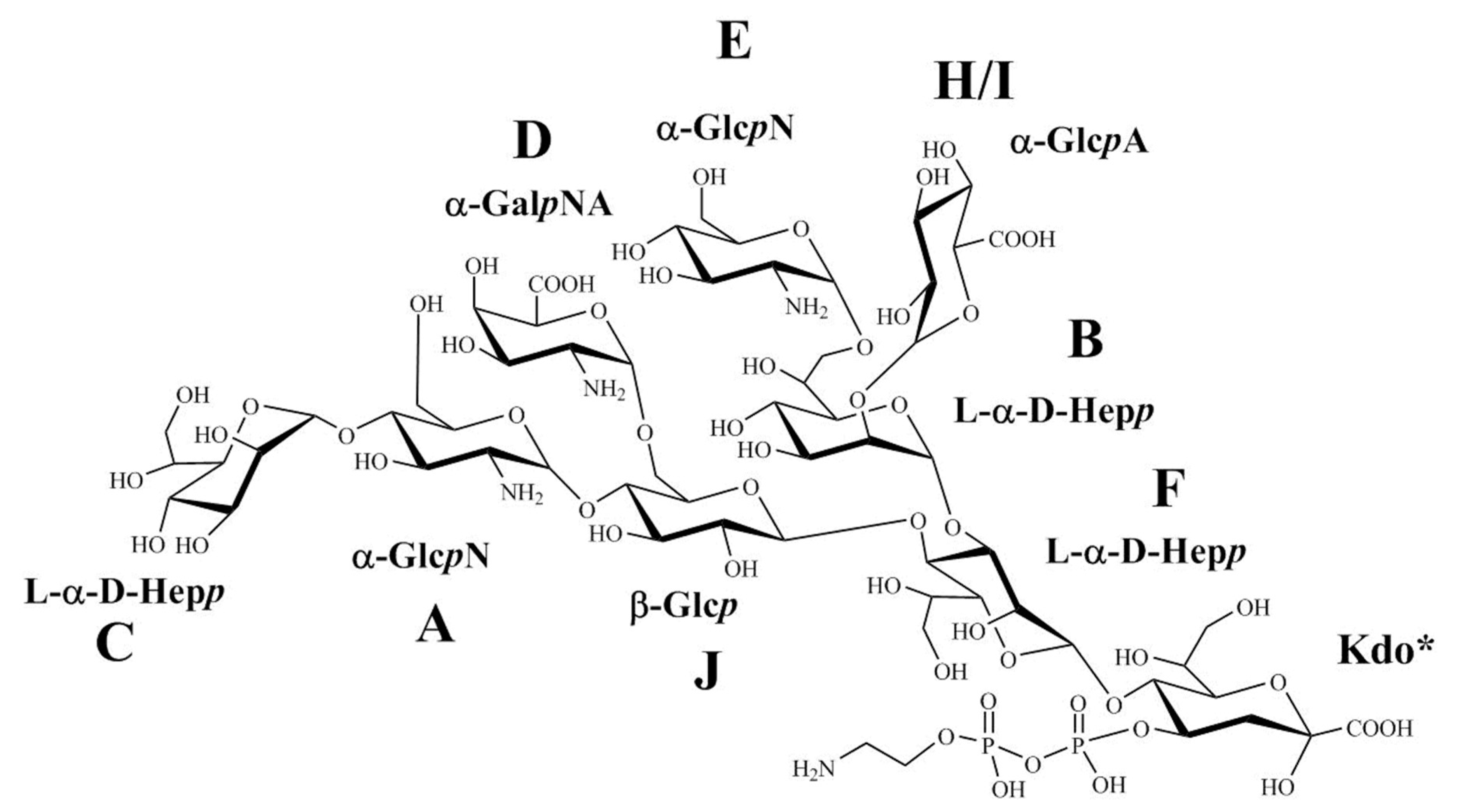
2.4. Structural Analysis of the OS Core
3. Discussion
4. Materials and Methods
4.1. Bacteria
4.2. Lipopolysaccharides and O-Specific Polysaccharide Fractions
4.3. Analytical Procedures
4.4. Partial HF Hydrolysis
4.5. SDS-PAGE and Serological Analysis
4.6. Mass Spectrometry
4.7. NMR Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LPS | Lipopolysaccharide |
| O-PS | O-specific polysaccharide |
| OS | Oligosaccharide |
| NMR | Nuclear Magnetic Resonance |
| COSY | Correlation spectroscopy |
| TOCSY | Total correlation spectroscopy |
| NOESY | Nuclear Overhauser Effect spectroscopy |
| HSQC | Heteronuclear Single Quantum Coherence |
| DEPT | Distortionless Enhancement by polarization transfer |
| HMBC | Heteronuclear Multiple Bond Correlation |
| GC | Gas chromatography |
| MS | Mass spectrometry |
| MALDI-TOF | Matrix-assisted laser desorption ionization time-of-flight |
| SDS | Sodium dodecyl sulfate |
References
- Austin, B. The Prokaryotes: Alphaproteobacteria and Betaproteobacteria. In The Family Alcaligenaceae; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer International Publishing: Berlin, Germany, 2014; pp. 729–757. [Google Scholar]
- Hamidou, S.I.; Linz, B.; Harvill, E.T. Environmental Origin of the Genus Bordetella. Front. Microbiol. 2017, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Weyant, R.S.; Hollis, D.G.; Weaver, R.E.; Amin, M.F.; Steigerwalt, A.G.; O’Connor, S.P.; Whitney, A.M.; Daneshvar, M.I.; Moss, C.W.; Brenner, D.J. Bordetella holmesii Sp. Nov., a New Gram-Negative Species Associated with Septicemia. J. Clin. Microbiol. 1995, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yih, W.K.; Silva, E.A.; Ida, J.; Harrington, N.; Lett, S.M.; George, H. Bordetella holmesii-Like Organisms Isolated from Massachusetts Patients with Pertussis-Like Symptoms. Emerg Infect. Dis. 1999, 5, 441–443. [Google Scholar] [CrossRef]
- Weiss, A. Proteobacteria: Alpha and Beta Subclasses. In The Genus Bordetella; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer International Publishing: New York, NY, USA, 2006; Volume 5, pp. 648–674. [Google Scholar] [CrossRef]
- Linz, B.; Ivanov, Y.V.; Preston, A.; Brinkac, L.; Parkhill, J.; Kim, M.; Harris, S.R.; Goodfield, L.L.; Fry, N.K.; Gorringe, A.R.; et al. Acquisition and Loss of Virulence-Associated Factors During Genome Evolution and Speciation in Three Clades of Bordetella Species. BMC Genom. 2016, 17, 767. [Google Scholar] [CrossRef] [PubMed]
- Shepard, C.W.; Daneshvar, M.I.; Kaiser, R.M.; Ashford, D.A.; Lonsway, D.; Patel, J.B.; Morey, R.E.; Jordan, J.G.; Weyant, R.S.; Fischer, M. Bordetella holmesii Bacteremia: A Newly Recognized Clinical Entity among Asplenic Patients. Clin. Infect. Dis. 2004, 38, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Diavatopoulos, D.A.; Cummings, C.A.; van der Heide, H.G.J.; Gent, M.V.; Liew, S.; Relman, D.A.; Mooi1, F.R. Characterization of a Highly Conserved Island in the Otherwise Divergent Bordetella holmesii and Bordetella pertussis Genomes. J. Bacteriol. 2006, 188, 8385–8394. [Google Scholar] [CrossRef]
- Harvill, E.T.; Goodfield, L.L.; Ivanov, Y.; Smallridge, W.E.; Meyer, J.A.; Cassiday, P.K.; Tondella, M.L.; Brinkac, L.; Sanka, R.; Kim, M.; et al. Genome Sequences of Nine Bordetella holmesii Strains Isolated in the United States. Genome Announc. 2014, 2, 14. [Google Scholar] [CrossRef]
- Mir-Cros, A.; Codina, G.; Martín-Gómez, M.T.; Fàbrega, A.; Martínez, X.; Jané, M.; Esso, D.V.; Cornejo, T.; Rodrigo, C.; Campins, M.; et al. Emergence of Bordetella holmesii as a Causative Agent of Whooping Cough, Barcelona, Spain. Emerg Infect. Dis. 2017, 23, 1856–1859. [Google Scholar] [CrossRef]
- Guthrie, J.L.; Robertson, A.V.; Tang, P.; Jamieson, F.; Drews, S.J. Novel Duplex Real-Time PCR Assay Detects Bordetella holmesii in Specimens from Patients with Pertussis-Like Symptoms in Ontario, Canada. J. Clin. Microbiol. 2010, 48, 1435–1437. [Google Scholar] [CrossRef]
- Rodgers, L.; Martin, S.W.; Cohn, A.; Budd, J.; Marcon, M.; Terranella, A.; Mandal, S.; Salamon, D.; Leber, A.; Tondella, M.L.; et al. Epidemiologic and Laboratory Features of a Large Outbreak of Pertussis-Like Illnesses Associated with Cocirculating Bordetella holmesii and Bordetella pertussis—Ohio, 2010–2011. Clin. Infect. Dis. 2013, 56, 322–331. [Google Scholar] [CrossRef]
- Njamkepo, E.; Bonacorsi, S.; Debruyne, M.; Gibaud, S.A.; Guillot, S.; Guiso, N. Significant Finding of Bordetella holmesii DNA in Nasopharyngeal Samples from French Patients with Suspected Pertussis. J. Clin. Microbiol. 2011, 49, 4347–4348. [Google Scholar] [CrossRef] [PubMed]
- Preston, A.; Petersen, B.O.; Duus, J.Ø.; Kubler-Kielb, J.; Ben-Menachem, G.; Li, J.J.; Vinogradov, E. Complete Structures of Bordetella bronchiseptica and Bordetella parapertussis Lipopolysaccharides. J. Biol. Chem. 2006, 281, 18135–18144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Weyrich, L.S.; Lavine, J.S.; Karanikas, A.T.; Harvill, E.T. Lack of Cross-protection against Bordetella holmesii after Pertussis Vaccination. Emerg. Infect. Dis. 2012, 18, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Bouchez, V.; AlBitar-Nehmé, S.; Novikov, A.; Guiso, N.; Caroff, M. Bordetella holmesii: Lipid A Structures and Corresponding Genomic Sequences Comparison in Three Clinical Isolates and the Reference Strain ATCC 51541. Int. J. Mol. Sci. 2017, 18, 1080. [Google Scholar] [CrossRef]
- Caroff, M.; Brisson, J.; Martin, A.; Karibian, D. Structure of the Bordetella pertussis 1414 Endotoxin. FEBS Lett. 2000, 477, 8–14. [Google Scholar] [CrossRef]
- Caroff, M.; Aussel, L.; Zarrouk, H. Structural Variability and Originality of the Bordetella endotoxins. J. Endotoxin Res. 2001, 7, 63–68. [Google Scholar] [CrossRef]
- Niedziela, T.; Letowska, I.; Lukasiewicz, J.; Kaszowska, M.; Czarnecka, A.; Kenne, L.; Lugowski, C. Epitope of the Vaccine-Type Bordetella pertussis Strain 186 Lipooligosaccharide and Antiendotoxin Activity of Antibodies Directed against the Terminal Pentasaccharide-Tetanus Toxoid Conjugate. Infect. Immun. 2005, 73, 7381–7389. [Google Scholar] [CrossRef]
- Vinogradov, E. The Structure of the Carbohydrate Backbone of the Lipopolysaccharides from Bordetella hinzii and Bordetella bronchiseptica. Eur. J. Biochem. 2000, 267, 4577–4582. [Google Scholar] [CrossRef]
- Odanaka, K.; Iwatsuki, M.; Satho, T.; Watanabe, M. Identification and Characterization of a Brilliant Yellow Pigment Produced by Bordetella pertussis. Microbiol. Immunol. 2017, 61, 490–496. [Google Scholar] [CrossRef]
- Ciucanu, I.; Kerek, F. A Simple and Rapid Method for the Permethylation of Carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Petersson, C.; Niedziela, T.; Jachymek, W.; Kenne, L.; Zarzecki, P.; Lugowski, C. Structural Studies of the O-Specific Polysaccharide of Hafnia alvei strain PCM 1206 Lipopolysaccharide Containing D-allothreonine. Eur. J. Biochem. 1997, 244, 580–586. [Google Scholar] [CrossRef] [PubMed]
- York, W.S.; Hantus, S.; Albersheim, P.; Darvill, A.G. Determination of the Absolute Configuration of Monosaccharides by 1H NMR Spectroscopy of their Per-O-(S)-2-methylbutyrate Derivatives. Carbohydr. Res. 1997, 300, 199–206. [Google Scholar] [CrossRef]
- McNicholas, P.A.; Batley, M.; Redmond, J.W. The Reactions of 3-deoxy-d-manno-oct-2-ulosonic Acid (KDO) in Dilute Acid. Carbohydr. Res. 1987, 165, 17–22. [Google Scholar] [CrossRef]
- Volk, W.A.; Salomonsky, N.L.; Hunt, D. Xanthomonas Sinensis Cell Wall Lipopolysaccharide. I. Isolation of 4,7-anhydro- and 4,8-anhydro-3-deoxy-octulosonic Acid Following Acid Hydrolysis of Xanthomonas Sinensis Lipopolysaccharide. J. Biol. Chem. 1972, 247, 3881–3887. [Google Scholar] [PubMed]
- Clark, H.W.; Mackay, R.M.; Deadman, M.E.; Hood, D.W.; Madsen, J.; Moxon, E.R.; Townsend, J.P.; Reid, K.B.M.; Ahmed, A.; Shaw, A.J. Crystal Structure of a Complex of Surfactant Protein D (SP-D) and Haemophilus influenzae Lipopolysaccharide Reveals Shielding of Core Structures in SP-D-Resistant Strains. Infect. Immun. 2016, 84, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Valero-Rello, A.; Henares, D.; Acosta, L.; Jane, M.; Jordan, I.; Godoy, P.; Muñoz-Almagro, C. Validation and Implementation of a Diagnostic Algorithm for DNA Detection of Bordetella pertussis, B. parapertussis, and B. holmesii in a Pediatric Referral Hospital in Barcelona, Spain. J. Clin. Microbiol. 2019, 57, 18. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Odanaka, K.; Otsuka, N.; Kamachi, K.; Watanabe, M. Development of Vaccines Against Pertussis Caused by Bordetella holmesii Using a Mouse Intranasal Challenge Model. Microbiol. Immunol. 2016, 60, 599–608. [Google Scholar] [CrossRef]
- Polak, M.; Zasada, A.A.; Mosiej, E.; Krysztopa-Grzybowska, K.; Witkowski, L.; Rzeczkowska, M.; Piekarska, K.; Lutyńska, A. Pertactin-Deficient Bordetella pertussis Isolates in Poland—A Country with Whole-Cell Pertussis Primary Vaccination. Microbes Infect. 2019, 21, 170–175. [Google Scholar] [CrossRef]
- Johnson, K.G.; Perry, M.B. Improved Techniques for the Preparation of Bacterial Lipopolysaccharides. Can. J. Microbiol. 1976, 22, 29–34. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins During the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Tsai, C.M.; Frasch, C.E. A Sensitive Silver Stain for Detecting Lipopolysaccharides in Polyacrylamide Gels. Anal. Biochem. 1982, 119, 115–119. [Google Scholar] [CrossRef]
- Koj, S.; Niedziela, T.; Lugowski, C. Bordetella Pertussis LOS-Derived Oligosaccharide with Pertussis Toxin Glycoconjugate and its Application in the Prophylaxis and Treatment of Infections Caused by Bordetella pertussis. U.S. Patent 9,878,051, 30 Junuary 2018. [Google Scholar]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced software for biomolecular NMR spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | Chemical Shifts (ppm) | |||||
|---|---|---|---|---|---|---|
| H-1 | H-2 | H-3 | H-4 | H-5 | H-6, H-6′ | |
| C-1 | C-2 | C-3 | C-4 | C-5 | C-6 | |
| A →3,4)-α-d-Galp-(1→ | 5.51 | 3.93 | 3.80 | 4.28 | 3.98 | 3.82, 3.82 |
| 99.7 | 68.7 | 79.6 | 77.1 | 72.5 | 60.7 | |
| B →3)-α-d-GlcpNAc-(1→ | 5.03 | 4.11 | 4.01 | 3.82 | 4.09 | 3.78, 3.78 |
| 96.8 | 52.7 | 76.8 | 71.2 | 72.6 | 60.7 | |
| C →2)-α-l-Rhap-(1→ | 4.89 | 4.04 | 3.95 | 3.51 | 3.79 | 1.34 |
| 99.0 | 76.3 | 70.1 | 72.7 | 71.2 | 17.9 | |
| C′ α-l-Rhap-(1→ b | 4.89 c | 4.02 | 3.81 | 3.44 | 3.78 | 1.31 |
| 99.0 | 70.8 | 69.8 | 73.0 | 69.8 | 17.8 | |
| D →6)-α-d-Glcp-(1→ | 4.92 | 3.51 | 3.72 | 3.57 | 4.34 | 3.93, 3.97 |
| 100.0 | 72.7 | 73.5 | 69.7 | 71.1 | 67.4 | |
| E β-d-GlcpNAc-(1→ | 4.59 | 3.64 | 3.56 | 3.43 | 3.43 | 3.74, 3.82 |
| 104.8 | 56.5 | 74.9 | 71.0 | 76.5 | 61.5 | |
| Residue | Atom H-1/C-1 | Connectivities to | Inter-Residue | |
|---|---|---|---|---|
| (ppm) | δC | δH | Atom/Residue | |
| A →3,4)-α-d-Galp-(1→ | 5.51/99.7 | 76.8 | 4.01 | C-3, H-3 of B |
| B →3)-α-d-GlcpNAc-(1→ | 5.03/96.8 | 76.3 | 4.04 | C-2, H-2 of C |
| C →2)-α-l-Rhap-(1→ | 4.89/99.0 | 67.4 | 3.93/3.97 | C-6, H-6/6′ of D |
| D →6)-α-d-Glcp-(1→ | 4.92/100.0 | 77.1 | 4.28 | C-4, H-4 of A |
| E β-d-GlcpNAc-(1→ | 4.59/104.8 | 79.6 | 3.80 | C-3, H-3 of A |
| Residue | Chemical Shifts (ppm) | |||||||
|---|---|---|---|---|---|---|---|---|
| H-1 | H-2 | H-3 | H-4 | H-5 | H-6, H-6′ | H-7 | H-8, H-8′ | |
| C-1 | C-2 | C-3 | C-4 | C-5 | C-6 | C-7 | C-8 | |
| Kdo 4,7-anhKdof | 3.12 | 4.39 | 4.02 | 4.14 | 3.89 | 3.66, 3.60 | ||
| 202.9 | 42.8 | 77.4 | 83.8 | 75.4 | 83.7 | 61.1 | ||
| Kdo’ 4,7-anhKdof | 3.09 | 4.51 | 4.12 | 4.08 | 3.74 | 3.67, 3.58 | ||
| 203.2 | 38.8 | 75.8 | 80.2 | 74.7 | 84.8 | 61.8 | ||
| A 4-α-GlcpN | 5.48 | 3.33 | 3.94 | 3.63 | 3.74 | 3.78, 3.73 | ||
| 97.3 | 54.5 | 70.3 | 74.9 | 71.9 | 60.5 | |||
| B 2,7-l-α-d-Hepp | 5.39 | 3.85 | 3.91 | 3.84 | 3.45 | 4.17 | 3.70 | |
| 99.7 | 79.8 | 70.4 | 66.5 | 72.1 | 67.9 | 70.3 | ||
| Cl-α- d-Hepp | 5.27 | 3.96 | 3.74 | 3.81 | 3.54 | 3.94 | 3.62 | |
| 101.2 | 70.1 | 70.5 | 65.9 | 72.5 | 68.5 | 62.7 | ||
| D α-GalpNA | 5.17 | 3.47 | 4.04 | 4.16 | 4.33 | |||
| 94.4 | 50.5 | 66.7 | 69.7 | 72.3 | 175.1 | |||
| E α-GlcpN | 5.12 | 3.28 | 3.86 | 3.44 | 3.70 | 3.78, 3.72 | ||
| 96.0 | 54.1 | 69.8 | 69.4 | 72.2 | 60.2 | |||
| F 3,4-l-α- d-Hepp | 5.07 | 3.90 | 3.84 | 4.23 | 3.53 | 3.93 | 3.67, 3.65 | |
| 97.8 | 73.3 | 76.7 | 71.7 | 71.4 | 68.9 | 62.7 | ||
| H α-GlcpA | 4.99 | 3.52 | 3.68 | 3.39 | 4.03 | |||
| 101.0 | 71.9 | 71.6 | 72.3 | 73.8 | 176.6 | |||
| I b α-GlcpA | 4.95 | 3.52 | 3.67 | 3.38 | 4.01 | |||
| 101.2 | 72.1 | 71.6 | 72.2 | 73.9 | 176.7 | |||
| J 4,6-β-Glcp | 4.40 | 3.27 | 3.55 | 3.44 | 3.66 | 3.76, 3.89 | ||
| 101.4 | 73.4 | 76.4 | 79.0 | 73.0 | 67.3 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ucieklak, K.; Koj, S.; Niedziela, T. Bordetella holmesii Lipopolysaccharide Hide and Seek Game with Pertussis: Structural Analysis of the O-Specific Polysaccharide and the Core Oligosaccharide of the Type Strain ATCC 51541. Int. J. Mol. Sci. 2020, 21, 6433. https://doi.org/10.3390/ijms21176433
Ucieklak K, Koj S, Niedziela T. Bordetella holmesii Lipopolysaccharide Hide and Seek Game with Pertussis: Structural Analysis of the O-Specific Polysaccharide and the Core Oligosaccharide of the Type Strain ATCC 51541. International Journal of Molecular Sciences. 2020; 21(17):6433. https://doi.org/10.3390/ijms21176433
Chicago/Turabian StyleUcieklak, Karolina, Sabina Koj, and Tomasz Niedziela. 2020. "Bordetella holmesii Lipopolysaccharide Hide and Seek Game with Pertussis: Structural Analysis of the O-Specific Polysaccharide and the Core Oligosaccharide of the Type Strain ATCC 51541" International Journal of Molecular Sciences 21, no. 17: 6433. https://doi.org/10.3390/ijms21176433