Modulation of the Paracrine Kynurenic System in Bone as a New Regulator of Osteoblastogenesis and Bone Mineral Status in an Animal Model of Chronic Kidney Disease Treated with LP533401

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of LP533401 Treatment on Tryptophan Utilization via the Kynurenine Pathway in Serum, Urine, and Intestinal Homogenate
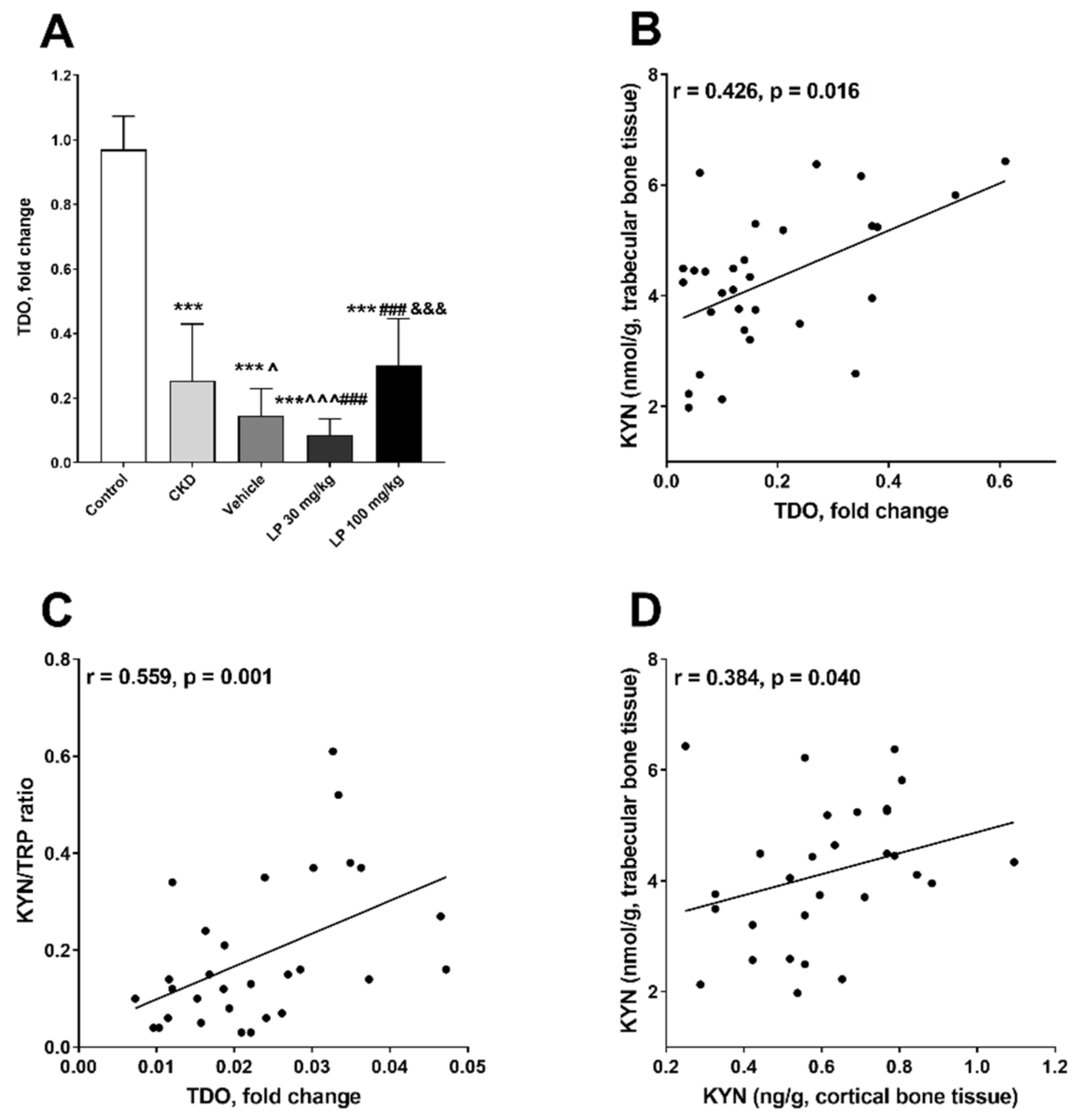
2.2. Effect of LP533401 Treatment on KYN Pathway Activation in Bone Tissue
2.3. The Associations between Bone Mineral Status and Components of the Bone Kynurenine Pathway
2.4. The Imbalance between Peripheral Serotonin and 25-Hydroxyvitamin D (25(OH)D) Status Affects TDO Expression in the Bone of CKD Rats Treated with LP533401
2.5. The Association between Serotonin-Dependent Molecular Pathway Involved in Osteoblast Formation and Activity of Kynurenic System in the Bone of Rats with CKD Treated with LP533401
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. TRP and KYN Quantification in Serum, Urine, and Bone and Intestinal Tissues
4.3. Tryptophan 2,3-Dioxygenase mRNA Expression Level Assessment
4.4. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 25(OH)D | 25-Hydroxyvitamin D |
| 5-HT | Serotonin |
| ATF4 | Activating transcription factor 4 |
| Bglap | Osteocalcin |
| BMA | Bone mineral area |
| BMC | Bone mineral content |
| BMD | Bone mineral density |
| cAMP | Cyclic adenosine monophosphate |
| CKD | Chronic kidney disease |
| CREB | cAMP-responsive element-binding protein |
| FOXO1 | Forkhead box protein O1 |
| HPLC | High-performance liquid chromatography |
| IDO | Indoleamine 2,3-dioxygenase |
| KYN | Kynurenine |
| MBD | Mineral and bone disorders |
| RNA | Ribonucleic acid |
| Sost | Sclerostin |
| TDO | Tryptophan 2,3-dioxygenase |
| TPH | Tryptophan hydroxylase |
| TRP | Tryptophan |
References
- Moe, S.M.; Drüeke, T.B.; Cunningham, J.; Goodman, W.; Martin, K.; Olgaard, K.; Ott, S.; Sprague, S.; Lameire, S.; Eknoyan, G. Definition, evaluation and classification of renal osteodystrophy: A position statement from Kidney Disease: Improving Global Outcomes (KDIGO). Kidney Int. 2006, 69, 1945–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moe, S.M.; Drüeke, T.B.; Block, G.A.; Cannata-Andia, J.B.; Elder, G.J.; Fukagawa, M.; Jorgetti, V.; Ketteler, M.; Langman, C.B.; Levin, A.; et al. KDIGO clinical practice guideline for the diagnosis, evaluation, prevention, and treatment of chronic kidney disease mineral and bone disorder (CKD-MBD). Kidney Int. Suppl. 2009, 76, S1–S130. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, D.; Nicholson, P.; Cheng, S.; Alen, M.; Mao, L.; Cheng, S. The associations of serum serotonin with bone traits are age- and gender-specific. PLoS ONE 2014, 9, e109028. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Karsenty, G. The two faces of serotonin in bone biology. J. Cell. Biol. 2010, 191, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Zofková, I.; Matucha, P. New insights into the physiology of bone regulation: The role of neurohormones. Physiol. Res. 2014, 63, 421–427. [Google Scholar]
- Carsote, M.; Radoi, V.; Galeriu, A.; Mihai, A.; Ferechide, D.; Opris, D.; Paun, D.; Poiana, C. Serotonin and the bone assessment. J. Med. Life 2014, 7, 49–53. [Google Scholar]
- Rauma, P.H.; Pasco, J.A.; Berk, M.; Stuart, A.L.; Koivumaa-Honkanen, H.; Honkanen, R.J.; Hodge, J.M.; Williams, L.J. The association between use of antidepressant and bone quality using quantitative heel ultrasound. Aust. N. Z. J. Psychiatry 2015, 49, 437–443. [Google Scholar] [CrossRef]
- Pawlak, D.; Oksztulska-Kolanek, E.; Znorko, B.; Domaniewski, T.; Rogalska, J.; Roszczenko, A.; Brzóska, M.M.; Pryczynicz, A.; Kemona, A.; Pawlak, K. The association between elevated levels of peripheral serotonin and its metabolite—5-hydroxyindoleacetic acid and bone strength and metabolism in growing rats with mild experimental chronic kidney disease. PLoS ONE 2016, 11, e0163526. [Google Scholar] [CrossRef]
- Pawlak, D.; Domaniewski, T.; Znorko, B.; Oksztulska-Kolanek, E.; Lipowicz, P.; Doroszko, M.; Karbowska, M.; Pawlak, K. The impact of peripheral serotonin on leptin-brain serotonin axis, bone metabolism and strength in growing rats with experimental chronic kidney disease. Bone 2017, 105, 1–10. [Google Scholar] [CrossRef]
- Yadav, V.K.; Balaji, S.; Suresh, P.S.; Liu, X.S.; Lu, X.; Li, Z.; Guo, E.; Mann, J.J.; Balapure, A.K.; Gershon, M.D.; et al. Pharmacological inhibition of gutderived serotonin synthesis is a potential bone anabolic treatment for osteoporosis. Nat. Med. 2010, 16, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, D.; Znorko, B.; Kalaska, B.; Domaniewski, T.; Zawadzki, R.; Lipowicz, P.; Doroszko, M.; Łebkowska, U.; Grabowski, P.; Pawlak, K. LP533401 restores bone health in 5/6 nephrectomized rats by a decrease of gut-derived serotonin and regulation of serum phosphate through the inhibition of phosphate co-transporters expression in the kidneys. Bone 2018, 113, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Schwarcz, R. The kynurenine pathway of tryptophan degradation as a drug target. Curr. Opin. Pharmacol. 2004, 4, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Thackray, S.J.; Mowat, C.G.; Chapman, S.K. Exploring the mechanism of tryptophan 2,3-dioxygenase. Biochem. Soc. Trans. 2008, 36, 1120–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badawy, A.A. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyes, M.P.; Saito, K.; Crowley, J.S.; Davis, L.E.; Demitrack, M.A.; Der, M.; Dilling, L.A.; Elia, J.; Kruesi, M.J.; Lackner, A.; et al. Quinolinic acid and kynurenine pathway metabolism in inflammatory and non-inflammatory neurological disease. Brain 1992, 115, 1249–1273. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccettiet, P. T cell apoptosis by tryptophan catabolism. Cell Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef]
- Topczewska-Bruns, J.; Pawlak, D.; Chabielska, E.; Tankiewicz, A.; Buczko, W. Increased levels of 3-hydroxykynurenine in different brain regions of rats with chronic renal insufficiency. Brain Res. Bull. 2002, 58, 423–428. [Google Scholar] [CrossRef]
- Topczewska-Bruns, J.; Tankiewicz, A.; Pawlak, D.; Buczko, W. Behavioral changes in the course of chronic renal insufficiency in rats. Pol. J. Pharmacol. 2001, 53, 263–269. [Google Scholar] [CrossRef]
- Forrest, C.M.; Mackay, G.M.; Oxford, L.; Stoy, N.; Stone, T.W.; Darlington, L.G. Kynurenine pathway metabolism in patients with osteoporosis after 2 years of drug treatment. Clin. Exp. Pharmacol. Physiol. 2006, 33, 1078–1087. [Google Scholar] [CrossRef]
- Dayer, M.R.; Safari, I.; Dayer, M.S. New evidence on hypoglycemic effect of quinolinic acid in diabetic rats. Pak. J. Biol. Sci. 2009, 12, 1025–1030. [Google Scholar] [CrossRef] [Green Version]
- Munipally, P.K.; Agraharm, S.G.; Valavala, V.K.; Gundae, S.; Turlapati, N.R. Evaluation of indoleamine 2,3-dioxygenase expression and kynurenine pathway metabolites levels in serum samples of diabetic retinopathy patients. Arch. Physiol. Biochem. 2011, 117, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, G.C. Cancer: Why tumours eat tryptophan. Nature 2011, 478, 192–194. [Google Scholar] [CrossRef]
- Apalset, E.M.; Gjesdal, C.G.; Ueland, P.M.; Midttun, O.; Ulvik, A.; Eide, G.E.; Tell, G.S. Interferon (IFN)-gamma-mediated inflammation and the kynurenine pathway in relation to bone mineral density: The Hordaland Health Study. Clin. Exp. Immunol. 2014, 176, 452–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez Esquivel, D.; Ramirez-Ortega, D.; Pineda, B.; Castro, N.; Rios, C.; Perez de la Cruz, V. Kynurenine pathway metabolites and enzymes involved in redox reactions. Neuropharmacology 2017, 112, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Li, W.; Santner-Nanan, B.; Lim, C.K.; Guillemin, G.J.; Ball, H.J.; Hunt, N.H.; Nanan, R.; Duqueet, G. The kynurenine pathway of tryptophan degradation is activated during osteoblastogenesis. Stem Cells 2015, 33, 111–121. [Google Scholar] [CrossRef]
- Al Saedi, A.; Sharma, S.; Summers, M.A.; Nurgali, K.; Duque, G. The multiple faces of tryptophan in bone biology. Exp. Gerontol. 2019, 129, 110778. [Google Scholar] [CrossRef]
- Kalaska, B.; Pawlak, K.; Domaniewski, T.; Oksztulska-Kolanek, E.; Znorko, B.; Roszczenko, A.; Rogalska, J.; Brzoska, M.M.; Lipowicz, P.; Doroszko, M.; et al. Elevated Levels of Peripheral Kynurenine Decrease Bone Strength in Rats with Chronic Kidney Disease. Front. Physiol. 2017, 8, 836. [Google Scholar] [CrossRef] [Green Version]
- Kalaska, B.; Pawlak, K.; Oksztulska-Kolanek, E.; Domaniewski, T.; Znorko, B.; Karbowska, M.; Citkowska, A.; Rogalska, J.; Roszczenko, A.; Brzoska, M.M.; et al. A link between central kynurenine metabolism and bone strength in rats with chronic kidney disease. Peer J. 2017, 5, e3199. [Google Scholar] [CrossRef] [Green Version]
- Green, A.R.; Curzon, G. Effects of hydrocortisone and immobilization on tryptophan metabolism in brain and liver of rats of different ages. Biochem. Pharmacol. 1975, 24, 713–716. [Google Scholar] [CrossRef]
- Comai, S.; Costa, C.V.L.; Ragazzi, E.; Bertazzo, A.; Allegri, G. The effect of age on the enzyme activities of tryptophan metabolism along the kynurenine pathway in rats. Clin. Chim. Acta 2005, 360, 67–80. [Google Scholar] [CrossRef]
- Pawlak, D.; Domaniewski, T.; Znorko, B.; Pawlak, K. The use of LP533401 as a therapeutic option for renal osteodystrophy affects, renal calcium handling, vitamin D metabolism, and bone health in uremic rats. Expert Opin. Ther. Targets 2019, 23, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, D.; Domaniewski, T.; Sieklucka, B.; Jakuc, M.; Pawlak, K. Inhibition of peripheral serotonin synthesis by LP533401 and disturbances in calciotropic hormones attenuated excessive osteoblastogenesis with simultaneous improvement of bone mineral status in 5/6 nephrectomized rats. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 165528. [Google Scholar] [CrossRef] [PubMed]
- El Refaey, M.; Watkins, C.P.; Kennedy, E.J.; Chang, A.; Zhong, Q.; Ding, K.H.; Shi, X.; Xu, J.; Bollag, W.B.; Hill, W.D.; et al. Oxidation of the aromatic amino acids tryptophan and tyrosine disrupts their anabolic effects on bone marrow mesenchymal stem cells. Mol. Cell. Endocrinol. 2015, 410, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moester, M.J.; Papapoulos, S.E.; Löwik, C.W.; van Bezooijen, R.L. Sclerostin: Current knowledge and future perspectives. Calcif. Tissue Int. 2010, 87, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, D.; Tankiewicz, A.; Buczko, W. Kynurenine and its metabolites in the rat with experimental renal insufficiency. J. Physiol. Pharmacol. 2001, 52, 755–766. [Google Scholar] [PubMed]
- Pawlak, D.; Tankiewicz, A.; Mysliwiec, P.; Buczko, W. Tryptophan metabolism via the kynurenine pathway in experimental chronic renal failure. Nephron 2002, 90, 328–335. [Google Scholar] [CrossRef]
- Pawlak, K.; Domaniewski, T.; Mysliwiec, M.; Pawlak, D. The kynurenines are associated with oxidative stress, inflammation and the prevalence of cardiovascular disease in patients with end-stage renal disease. Atherosclerosis 2009, 204, 309–314. [Google Scholar] [CrossRef]
- Pawlak, K.; Myśliwiec, M.; Pawlak, D. Kynurenine pathway—A new link between endothelial dysfunction and carotid atherosclerosis in chronic kidney disease patients. Adv. Med. Sci. 2010, 55, 196–203. [Google Scholar] [CrossRef]
- Schefold, J.C.; Zeden, J.P.; Fotopoulou, C.; von Haehling, S.; Pschowski, R.; Hasper, D.; Volk, H.D.; Schuett, C.; Reinke, P. Increased indoleamine 2,3-dioxygenase (IDO) activity and elevated serum levels of tryptophan catabolites in patients with chronic kidney disease: A possible link between chronic inflammation and uraemic symptoms. Nephrol. Dial. Transplant. 2009, 24, 1901–1908. [Google Scholar] [CrossRef] [Green Version]
- Danesch, U.; Glossl, B.; Schmid, W.; Schutz, G.; Schule, R.; Renkawitz, R. Glucocorticoid induction of the rat tryptophan oxygenase gene is mediated by two widely separated glucocorticoid-responsive elements. EMBO J. 1987, 6, 625–630. [Google Scholar] [CrossRef]
- Kanai, M.; Funakoshi, H.; Takahashi, H.; Hayakawa, T.; Mizuno, S.; Matsumoto, K.; Nakamura, T. Tryptophan 2,3-dioxygenase is a key modulator of physiological neurogenesis and anxiety-related behavior in mice. Mol. Brain 2009, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Refaey, M.; McGee-Lawrence, M.E.; Fulzele, S.; Kennedy, E.J.; Bollag, W.B.; Elsalanty, M.; Zhong, Q.; Ding, K.H.; Bendzunas, N.G.; Shi, X.M.; et al. Kynurenine, a Tryptophan Metabolite That Accumulates With Age, Induces Bone Loss. J. Bone Miner. Res. 2017, 32, 2182–2193. [Google Scholar] [CrossRef] [PubMed]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Kode, A.; Mosialou, I.; Silva, B.C.; Rached, M.T.; Zhou, B.; Wang, J.; Townes, T.M.; Hen, R.; DePinho, R.A.; Guo, X.E.; et al. FOXO1 orchestrates the bone-suppressing function of gut-derived serotonin. J. Clin. Investig. 2012, 122, 3490–3503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, N.S.; Pettway, G.J.; Chen, C.; Koh, A.J.; McCauley, L.K. Cyclin D1 as a target for the proliferative effects of PTH and PTHrP in early osteoblastic cells. J. Bone Miner. Res. 2007, 22, 951–964. [Google Scholar] [CrossRef]
- Pereira, R.C.; Delany, A.M.; Khouzam, N.M.; Bowen, R.E.; Freymiller, E.G.; Salusky, I.B.; Wesseling-Perry, K. Primary osteoblast-like cells from patients with end-stage kidney disease reflect gene expression, proliferation, and mineralization characteristics ex vivo. Kidney Int. 2015, 87, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Inose, H.; Zhou, B.; Yadav, V.K.; Guo, X.E.; Karsenty, G.; Ducy, P. Efficacy of serotonin inhibition in mouse models of bone loss. J. Bone Miner. Res. 2011, 26, 2002–2011. [Google Scholar] [CrossRef]
- Lima, G.M.; Corazza, B.J.M.; Moraes, R.M.; de Oliveira, F.E.; de Oliveira, L.; Franco, G.N.; Perrien, D.S.; Elefteriou, F.; Anbinder, A.L. The effect of an inhibitor of gut serotonin (LP533401) during the induction of peridontal disease. J. Periodontal Res. 2016, 51, 661–668. [Google Scholar] [CrossRef] [Green Version]
- Holmes, E.W. Determination of serum kynurenine and hepatic tryptophan dioxygenase activity by high-performance liquid chromatography. Anal. Biochem. 1988, 172, 518–525. [Google Scholar] [CrossRef]
- Znorko, B.; Pawlak, D.; Oksztulska-Kolanek, E.; Domaniewski, T.; Pryczynicz, A.; Roszczenko, A.; Rogalska, J.; Lipowicz, P.; Doroszko, M.; Brzoska, M.M.; et al. RANKL/OPG system regulation by endogenous PTH and PTH1R/ATF4 axis in bone: Implications for bone accrual and strength in growing rats with mild uremia. Cytokine 2018, 106, 19–28. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mor, A.; Pawlak, K.; Kalaska, B.; Domaniewski, T.; Sieklucka, B.; Zieminska, M.; Cylwik, B.; Pawlak, D. Modulation of the Paracrine Kynurenic System in Bone as a New Regulator of Osteoblastogenesis and Bone Mineral Status in an Animal Model of Chronic Kidney Disease Treated with LP533401. Int. J. Mol. Sci. 2020, 21, 5979. https://doi.org/10.3390/ijms21175979
Mor A, Pawlak K, Kalaska B, Domaniewski T, Sieklucka B, Zieminska M, Cylwik B, Pawlak D. Modulation of the Paracrine Kynurenic System in Bone as a New Regulator of Osteoblastogenesis and Bone Mineral Status in an Animal Model of Chronic Kidney Disease Treated with LP533401. International Journal of Molecular Sciences. 2020; 21(17):5979. https://doi.org/10.3390/ijms21175979
Chicago/Turabian StyleMor, Adrian, Krystyna Pawlak, Bartlomiej Kalaska, Tomasz Domaniewski, Beata Sieklucka, Marta Zieminska, Bogdan Cylwik, and Dariusz Pawlak. 2020. "Modulation of the Paracrine Kynurenic System in Bone as a New Regulator of Osteoblastogenesis and Bone Mineral Status in an Animal Model of Chronic Kidney Disease Treated with LP533401" International Journal of Molecular Sciences 21, no. 17: 5979. https://doi.org/10.3390/ijms21175979