Lactobacillus Strains Alleviated Hyperlipidemia and Liver Steatosis in Aging Rats via Activation of AMPK


Abstract
:1. Introduction
2. Results
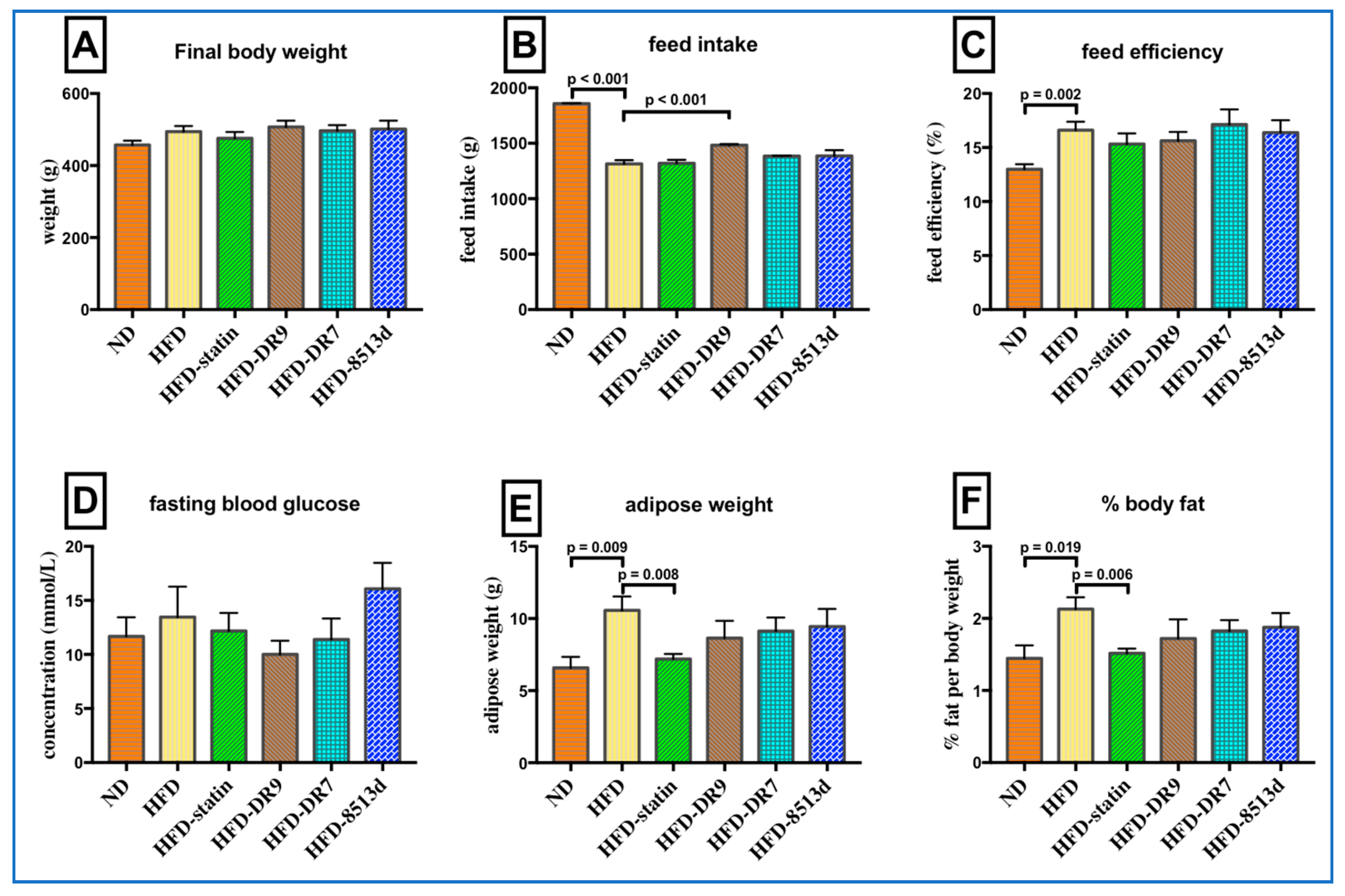
2.1. Body Weight, Feed Intake, Feed Efficacy, Fasting Blood Glucose, Adipose Weight, and Percent Body Fat
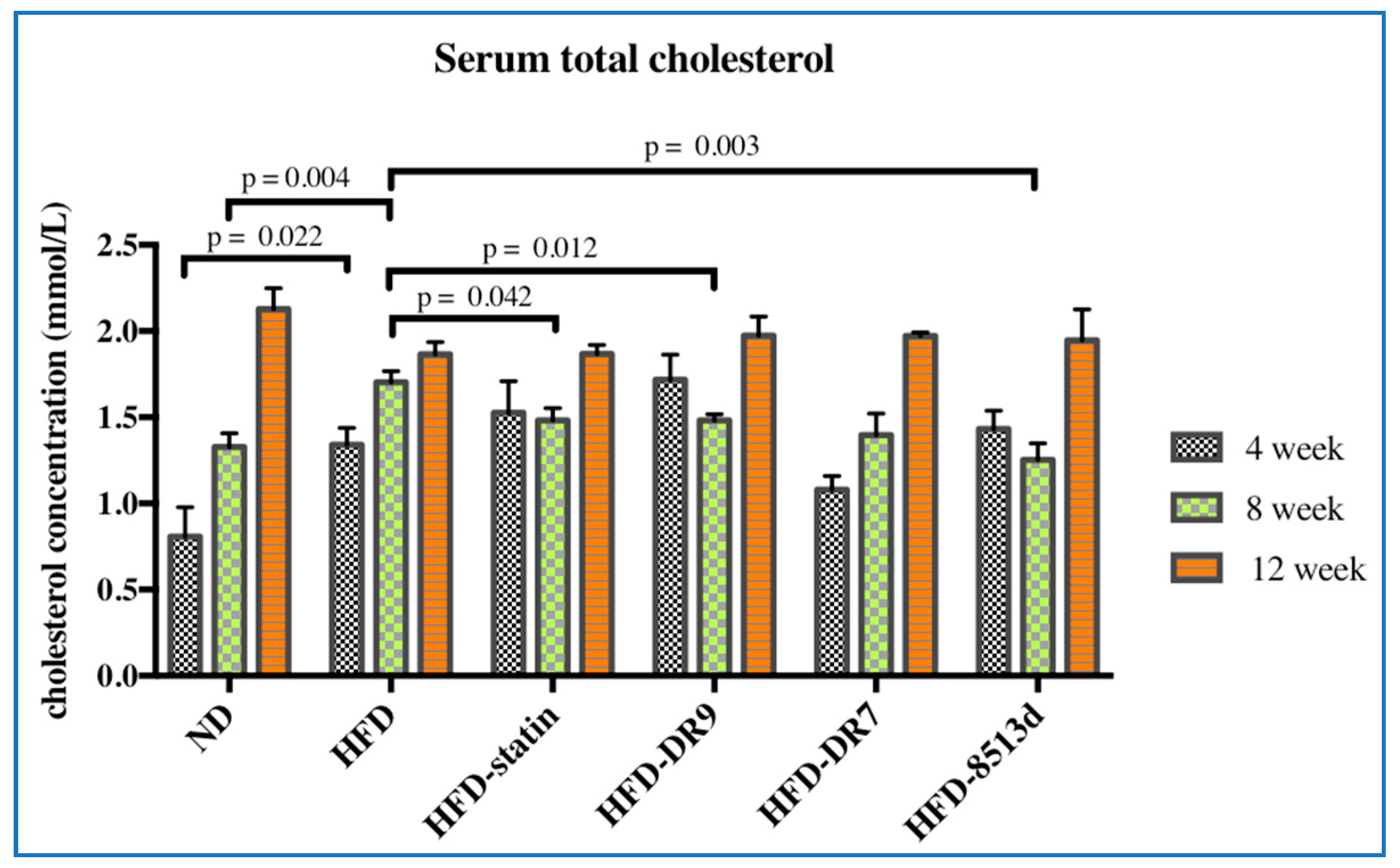
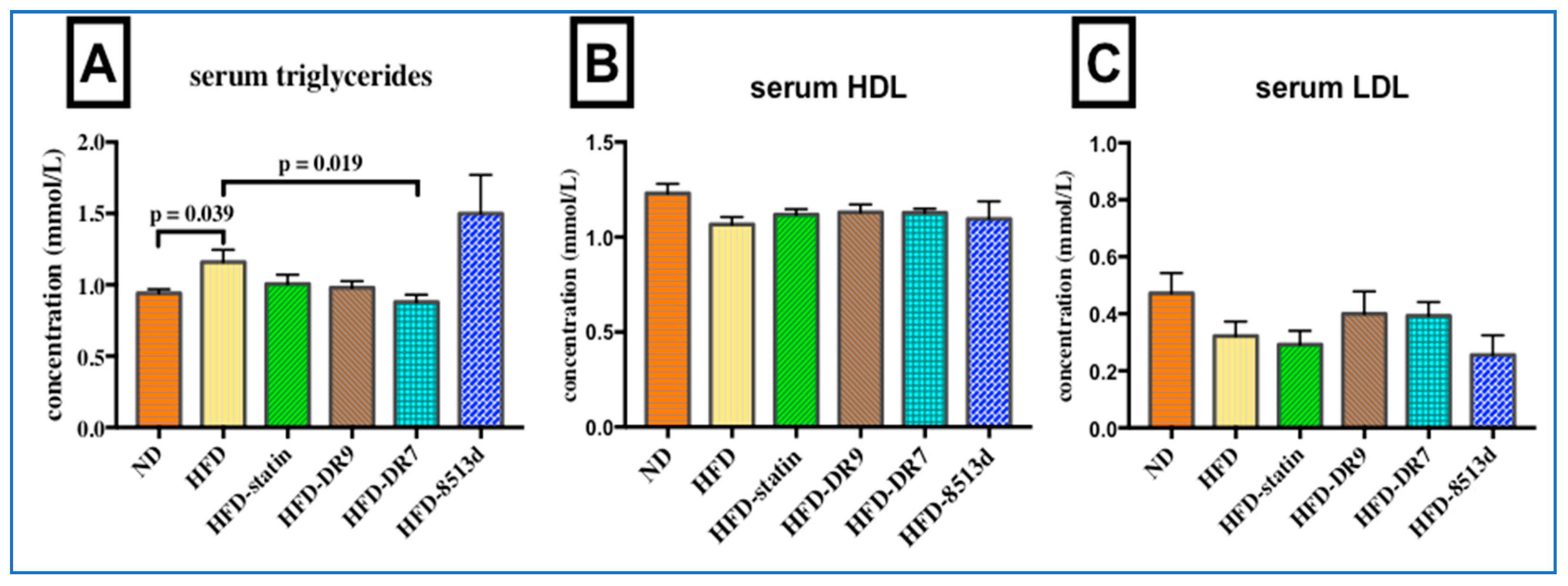
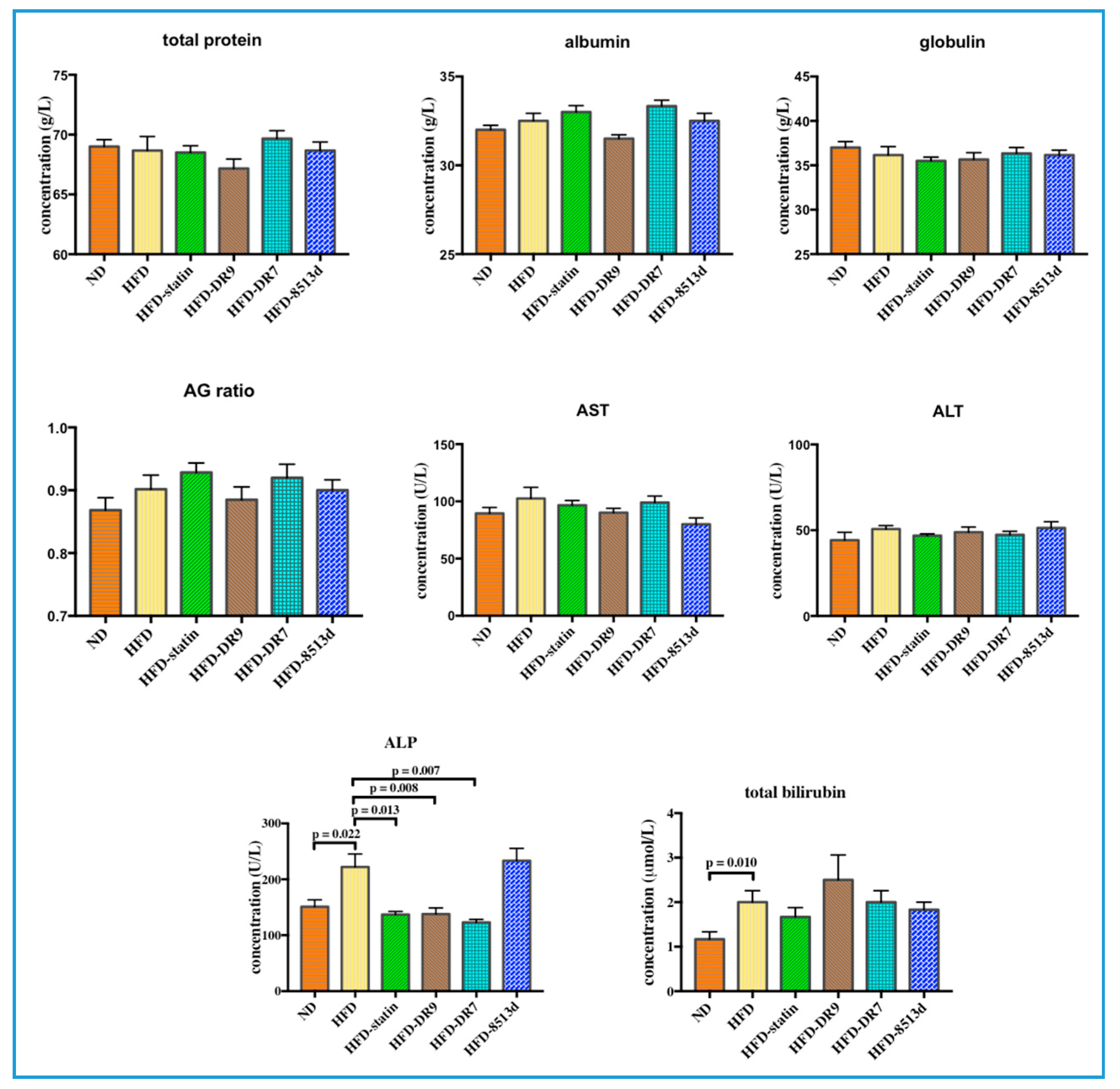
2.2. Serum Biochemical Analysis
2.2.1. Serum Lipid Profile
2.2.2. Liver Function Test
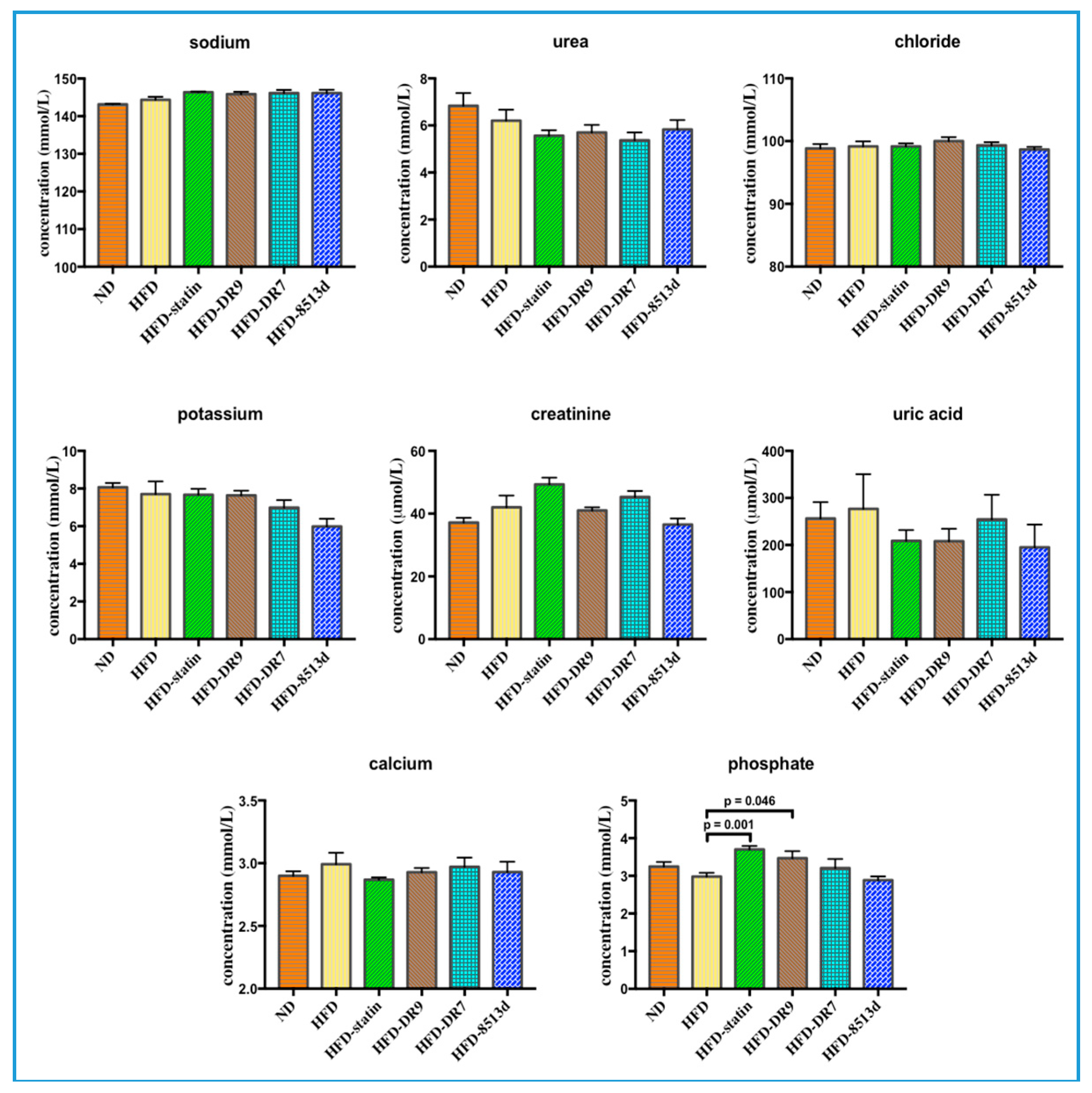
2.2.3. Renal Function Test
2.3. Gene Expression of Lipid Metabolism Pathway
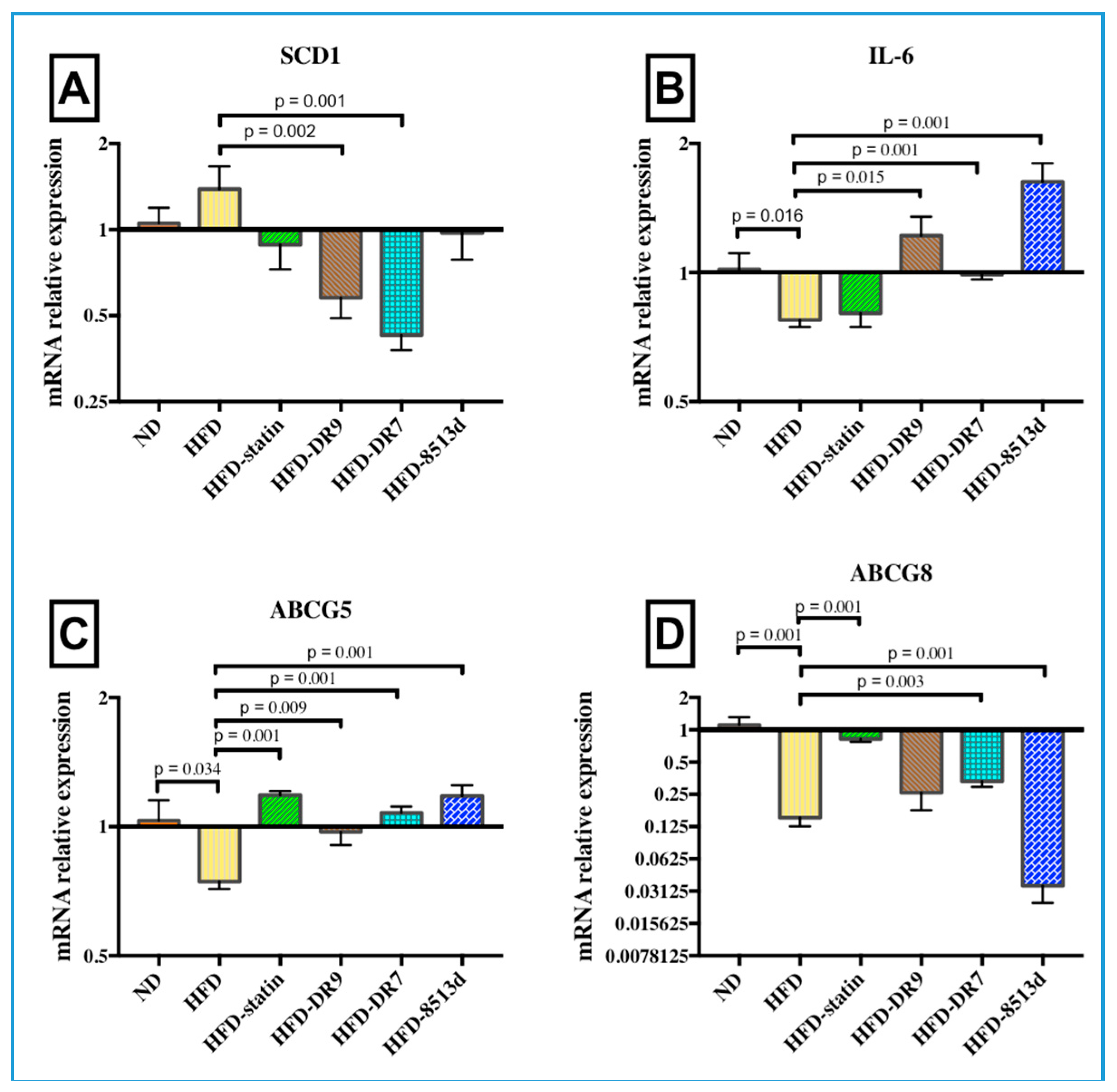
2.3.1. Stearoyl-CoA Desaturase 1 (SCD1)
2.3.2. Interleukin-6 (IL-6)
2.3.3. ATP-Binding Cassette Subfamily G Member 5 and 8 (ABCG5 and ABCG8)
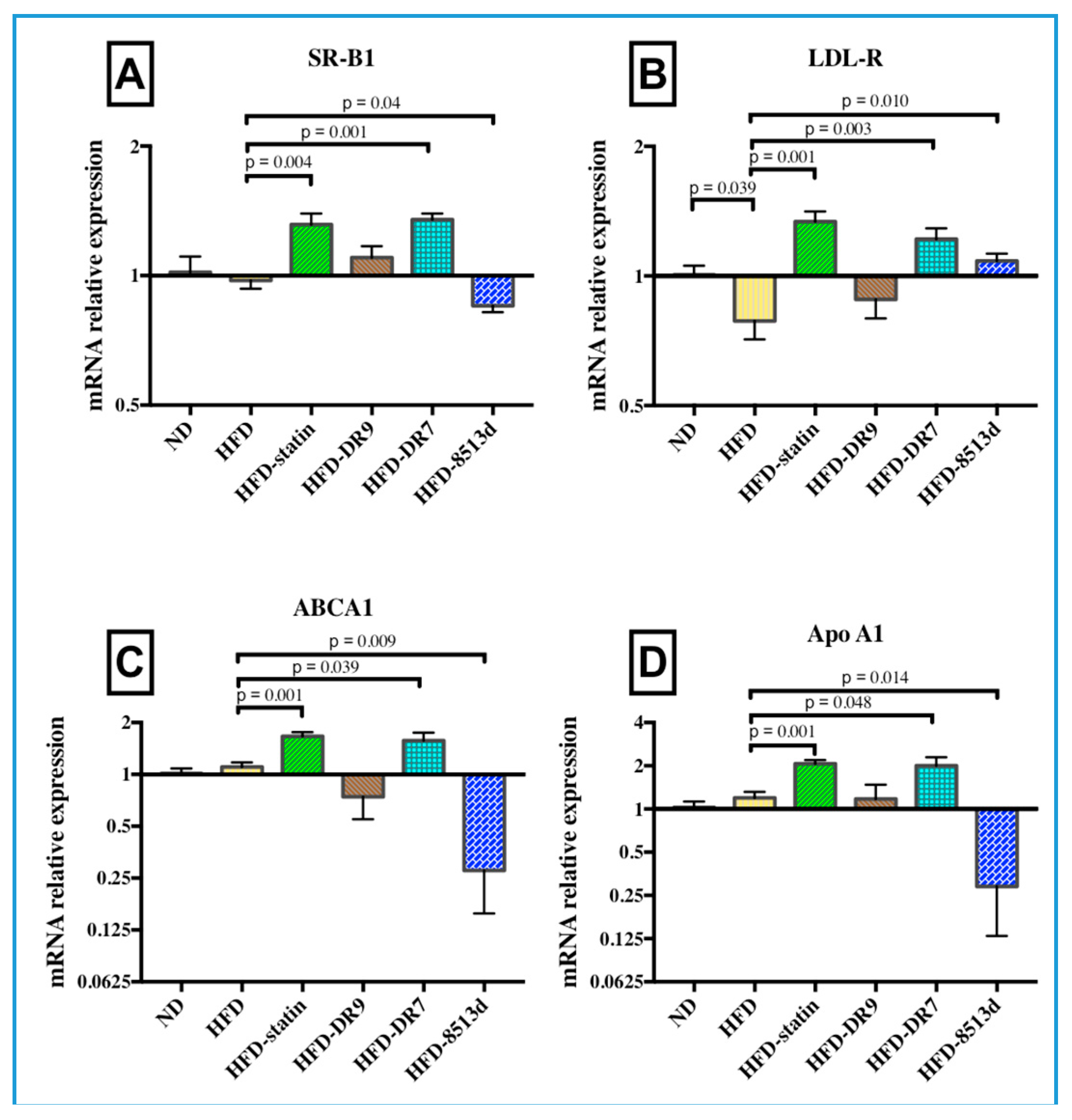
2.3.4. Scavenger Receptor B1 (SR-B1)
2.3.5. Low Density Lipoprotein Receptor (LDL-R)
2.3.6. ATP-Binding Cassette Subfamily A Member 1 (ABCA1)
2.3.7. Apolipoprotein A1 (Apo A1)
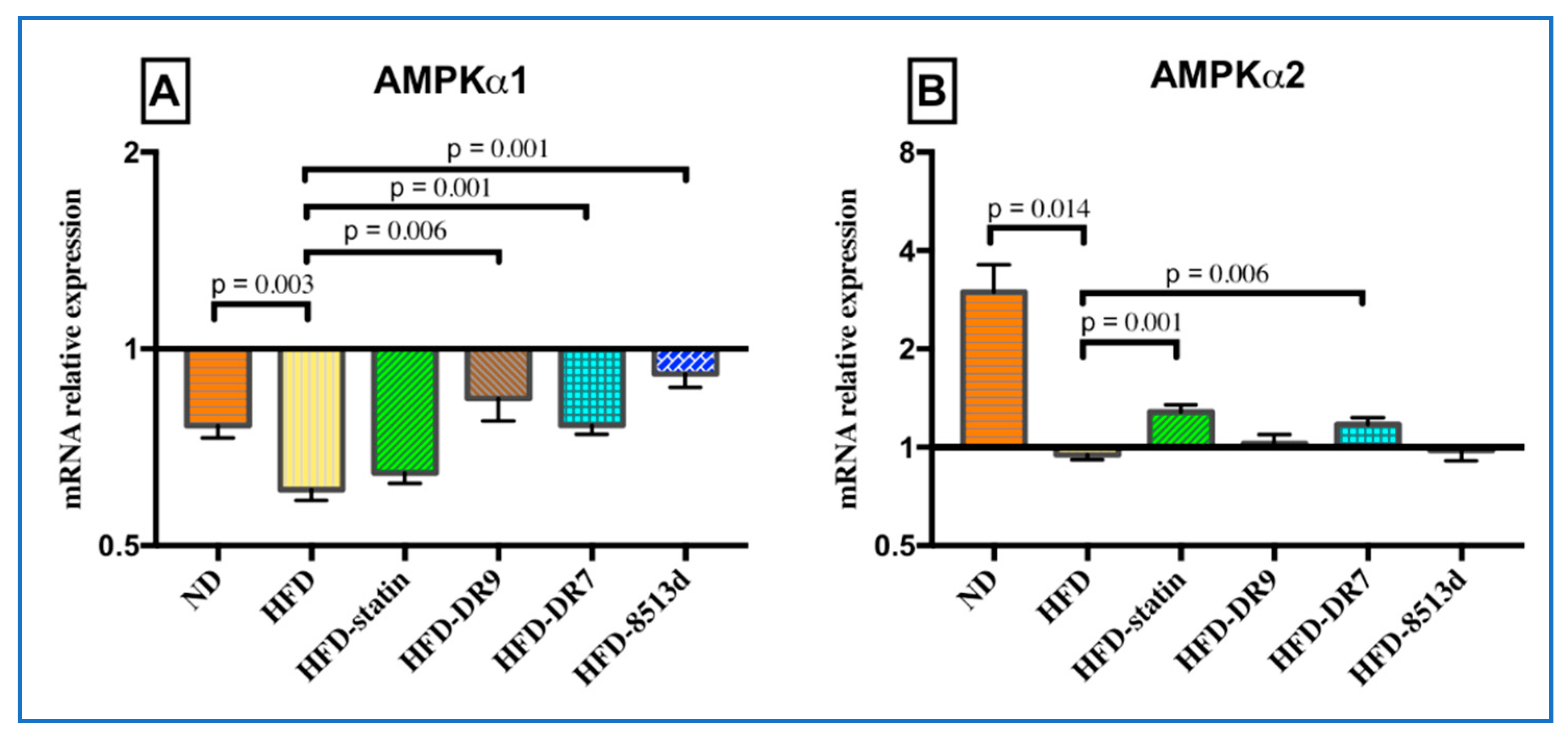
2.3.8. 5′ Adenosine Monophosphate-Activated Protein Kinase (AMPK)
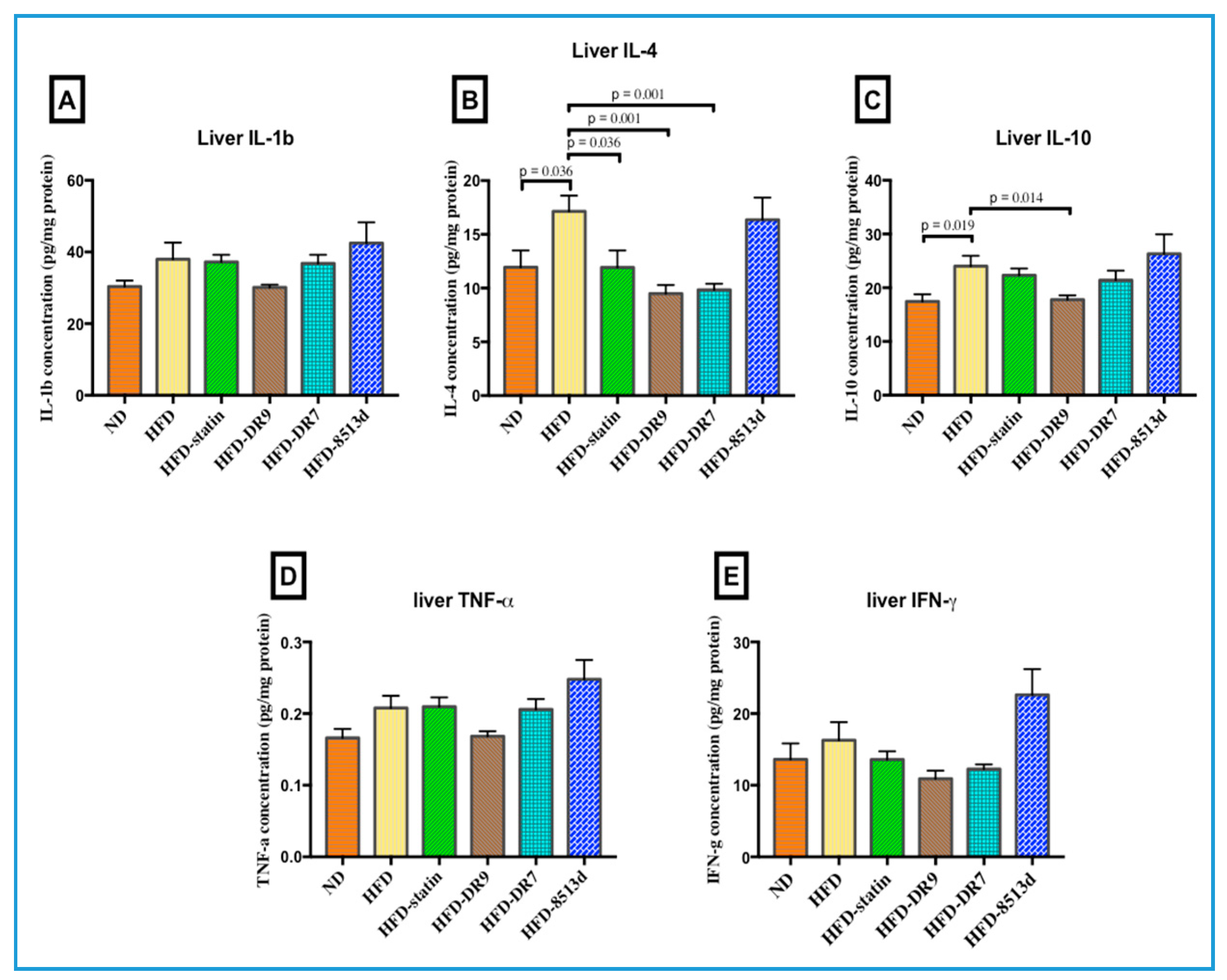
2.4. Liver Immune Response: IL-1β, IL-4, IL-10, TNF-α, and IFN-γ
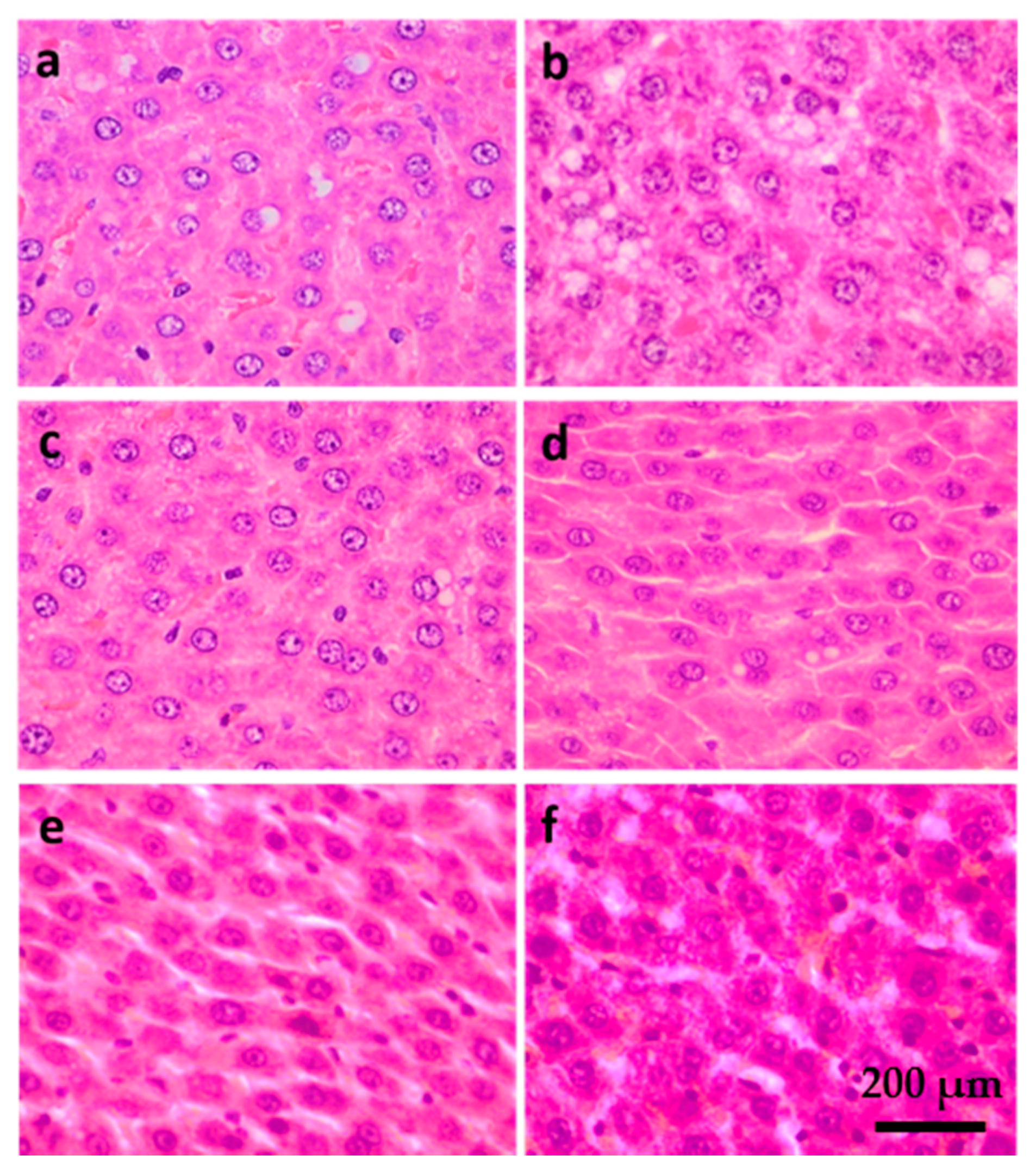
2.5. Liver Histology
3. Discussion
4. Materials and Methods
4.1. Bacterial Culture
4.2. Animal Experiments
4.3. Collection of Tissues
4.4. Serum Biochemical Analysis
4.5. Total RNA Isolation and cDNA Conversion
4.6. qRT-PCR
4.7. Multiplex ELISA
4.8. Liver Histology
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviation
| ABCA1 | ATP-binding cassette subfamily A member 1 |
| ABCG5 & ABCG8 | ATP-binding cassette subfamily G member 5 and 8 |
| AMPK | 5’ adenosine monophosphate-activated protein kinase |
| ALP | Alkaline phosphatase |
| ALT | Alanine aminotransferase |
| Apo A1 | Apolipoprotein A1 |
| AST | Aspartate aminotransferase |
| CVD | Cardiovascular disease |
| HDL | High-density lipoprotein |
| HFD | High-fat diet |
| IL-6 | Interleukin-6 |
| LDL-C | Low-density lipoprotein cholesterol |
| NAFLD | Non-alcoholic fatty liver disease |
| ND | Normal diet |
| SCD1 | Stearoyl-CoA desaturase 1 |
| SR-B1 | Scavenger Receptor B1 |
| TC | Total cholesterol |
| TG | Total triglyceride |
References
- Trapani, L.; Pallottini, V. Age-related hypercholesterolemia and HMG-CoA reductase dysregulation: Sex does matter (A Gender Perspective). Curr. Gerontol. Geriatr. Res. 2010, 2010, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.H. Hyperlipidemia as a risk factor for cardiovascular disease. Prim. Care 2012, 40, 195–211. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.V.; Shrivastava, A.J.; Chaturvedi, U.; Singh, S.C.; Shanker, K.; Saxena, J.K.; Bhatia, G.; Pal, A. A mechanism-based pharmacological evaluation of efficacy of Flacourtia indica in management of dyslipidemia and oxidative stress in hyperlipidemic rats. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 121–129. [Google Scholar] [CrossRef]
- Tomizawa, M.; Kawanabe, Y.; Shinozaki, F.; Sato, S.; Motoyoshi, Y.; Sugiyama, T.; Yamamoto, S.; Sueishi, M. Triglyceride is strongly associated with nonalcoholic fatty liver disease among markers of hyperlipidemia and diabetes. Biomed. Rep. 2014, 2, 633–636. [Google Scholar] [CrossRef]
- Wadhera, R.K.; Steen, D.L.; Khan, I.; Giugliano, R.P.; Foody, J.M. A review of low-density lipoprotein cholesterol, treatment strategies, and its impact on cardiovascular disease morbidity and mortality. J. Clin. Lipidol. 2016, 10, 472–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, B.C.; Staplin, N.; Haynes, R.; Reith, C.; Emberson, J.; Herrington, W.G.; Wheeler, D.C.; Walker, R.; Fellström, B.; Wanner, C.; et al. Lowering LDL cholesterol reduces cardiovascular risk independently of presence of inflammation. Kidney Int. 2017, 93, 1000–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Cholesterol Education Program Expert Panel on Detection Evaluation and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III) final report. Circulation 2002, 106, 3143–3421. [Google Scholar] [CrossRef]
- Seki, R.; Inoue, K.; Yamamoto, S.; Akimoto, K. Non-HDL cholesterol is better than Friedewald-Estimated LDL cholesterol to associate with cardiometabolic markers. Biomed. Res. Clin. Pract. 2017, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tham, C.S.-C.; Peh, K.-K.; Bhat, R.; Liong, M.-T. Probiotic properties of bifidobacteria and lactobacilli isolated from local dairy products. Ann. Microbiol. 2011, 62, 1079–1087. [Google Scholar] [CrossRef]
- Hor, Y.Y.; Liong, M.-T. Use of extracellular extracts of lactic acid bacteria and bifidobacteria for the inhibition of dermatological pathogen Staphylococcus aureus. Dermatol. Sin. 2014, 32, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Fung, W.-Y.; Liong, M.-T. Evaluation of proteolytic and ACE-inhibitory activity of Lactobacillus acidophilus in soy whey growth medium via response surface methodology. LWT Food Sci. Tehcnol. 2010, 43, 563–567. [Google Scholar] [CrossRef]
- Ben Salah, R.; Trabelsi, I.; Hamden, K.; Chouayekh, H.; Bejar, S. Lactobacillus plantarum TN8 exhibits protective effects on lipid, hepatic and renal profiles in obese rat. Anaerobe 2013, 23, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Cui, Y.; Yin, Y.-N.; Zhao, X.; Yang, J.-W.; Wang, Z.-G.; Fu, N.; Tang, Y.; Wang, X.-H.; Liu, X.-W.; et al. Effects of two Lactobacillus strains on lipid metabolism and intestinal microflora in rats fed a high-cholesterol diet. BMC Complement. Altern. Med. 2011, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, Q.; Ren, Y.; Ruan, Z. Effect of probiotic Lactobacillus on lipid profile: A systematic review and meta-analysis of randomized, controlled trials. PLoS ONE 2017, 12, e0178868. [Google Scholar] [CrossRef]
- Lew, L.C.; Choi, S.-B.; Khoo, B.-Y.; Sreenivasan, S.; Ong, K.-L.; Liong, M.-T. Lactobacillus plantarum DR7 reduces cholesterol via phosphorylation of AMPK that down-regulated the mRNA expression of HMG-CoA reductase. Korean J. Food Sci. Anim. 2018, 38, 350–361. [Google Scholar]
- Foretz, M.; Even, P.; Viollet, B. AMPK activation reduces hepatic lipid content by increasing fat oxidation in vivo. Int. J. Mol. Sci. 2018, 19, 2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Chen, F.; Zhou, B. Antioxidative, anti-inflammatory and anti-apoptotic effects of ellagic acid in liver and brain of rats treated by D-galactose. Sci. Rep. 2018, 8, 1465. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.; Su, X.; Liu, J.; Zhao, Y.; Li, Z.; Xu, X.; Li, H.; Nashun, B. Comparison of naturally aging and D-galactose induced aging model in beagle dogs. Exp. Ther. Med. 2017, 14, 5881–5888. [Google Scholar] [CrossRef] [Green Version]
- Bendali, F.; Kerdouche, K.; Hamma-Faradji, S.; Drider, D. In vitro and in vivo cholesterol lowering ability of Lactobacillus pentosus KF923750. Benef. Microbes 2017, 8, 271–280. [Google Scholar] [CrossRef]
- Hu, X.; Wang, T.; Li, W.; Jin, F.; Wang, L. Effects of NS lactobacillus strains on lipid metabolism of rats fed a high-cholesterol diet. Lipids Health Dis. 2013, 12, 67. [Google Scholar] [CrossRef] [Green Version]
- Rerksuppaphol, S.; Rerksuppaphol, L. A randomized double-blind controlled trial of Lactobacillus acidophilus plus Bifidobacterium bifidum versus placebo in patients with hypercholesterolemia. J. Clin. Diagn. Res. 2015, 9, KC01–KC04. [Google Scholar] [CrossRef] [PubMed]
- Lye, H.-S.; Rahmat-Ali, G.R.; Liong, M.-T. Mechanisms of cholesterol removal by lactobacilli under conditions that mimic the human gastrointestinal tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Lye, H.-S.; Kato, T.; Low, W.-Y.; Taylor, T.; Prakash, T.; Lew, L.C.; Ohno, H.; Liong, M.-T. Lactobacillus fermentum FTDC 8312 combats hypercholesterolemia via alteration of gut microbiota. J. Biotechnol. 2017, 262, 75–83. [Google Scholar] [CrossRef]
- Hong, Y.-F.; Kim, H.; Kim, H.S.; Park, W.J.; Kim, J.-Y.; Chung, D.K. Lactobacillus acidophilus K301 inhibits atherogenesis via induction of 24 (S), 25-epoxycholesterol-mediated ABCA1 and ABCG1 production and cholesterol efflux in macrophages. PLoS ONE 2016, 11, e0154302. [Google Scholar] [CrossRef] [Green Version]
- Michael, D.R.; Davies, T.S.; Moss, J.; Calvente, D.L.; Ramji, D.P.; Marchesi, J.R.; Pechlivanis, A.; Plummer, S.F.; Hughes, T.R. The anti-cholesterolaemic effect of a consortium of probiotics: An acute study in C57BL/6J mice. Sci. Rep. 2017, 7, 2883. [Google Scholar] [CrossRef] [Green Version]
- Enoch, H.G.; Catalá, A.; Strittmatter, P. Mechanism of rat liver microsomal stearyl-CoA desaturase. Studies of the substrate specificity, enzyme-substrate interactions, and the function of lipid. J. Biol. Chem. 1976, 251, 5095–5103. [Google Scholar]
- Miyazaki, M.; Kim, Y.-C.; Gray-Keller, M.P.; Attie, A.D.; Ntambi, J.M. The biosynthesis of hepatic cholesterol esters and triglycerides is impaired in mice with a disruption of the gene for stearoyl-CoA desaturase 1. J. Biol. Chem. 2000, 275, 30132–30138. [Google Scholar] [CrossRef] [Green Version]
- Attie, A.; Krauss, R.M.; Gray-Keller, M.P.; Brownlie, A.; Miyazaki, M.; Kastelein, J.J.; Lusis, A.J.; Stalenhoef, A.F.H.; Stoehr, J.P.; Hayden, M.R.; et al. Relationship between stearoyl-CoA desaturase activity and plasma triglycerides in human and mouse hypertriglyceridemia. J. Lipid Res. 2002, 43, 1899–1907. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Lee, J.-H.; Ntambi, J.M.; Hyun, C.-K. Inhibition of stearoyl-CoA desaturase1 activates AMPK and exhibits beneficial lipid metabolic effects in vitro. Eur. J. Pharmacol. 2011, 672, 38–44. [Google Scholar] [CrossRef]
- Dobrzyń, P.; Dobrzyń, A.; Miyazaki, M.; Cohen, P.; Asilmaz, E.; Hardie, D.G.; Friedman, J.M.; Ntambi, J.M. Stearoyl-CoA desaturase 1 deficiency increases fatty acid oxidation by activating AMP-activated protein kinase in liver. Proc. Natl. Acad. Sci. USA 2004, 101, 6409–6414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntambi, J.M.; Miyazaki, M.; Stoehr, J.P.; Lan, H.; Kendziorski, C.M.; Yandell, B.S.; Song, Y.; Cohen, P.; Friedman, J.M.; Attie, A.D. Loss of stearoyl-CoA desaturase-1 function protects mice against adiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Hawley, S.A.; Davison, M.; Woods, A.; Davies, S.P.; Beri, R.K.; Carling, D.; Hardie, D.G. Characterization of the AMP-activated protein kinase kinase from rat liver and identification of threonine 172 as the major site at which it phosphorylates AMP-activated protein kinase. J. Biol. Chem. 1996, 271, 27879–27887. [Google Scholar] [CrossRef] [Green Version]
- Towler, M.C.; Hardie, D.G.; Hardie, D.G. AMP-activated protein kinase in metabolic control and insulin signaling. Circ. Res. 2007, 100, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-S.; Cho, S.-M.; Lee, M.-H.; Lee, E.-O.; Kim, S.-H.; Lee, H.-J. Ethanol extract of Pinus koraiensis leaves containing lambertianic acid exerts anti-obesity and hypolipidemic effects by activating adenosine monophosphate-activated protein kinase (AMPK). BMC Complement. Altern. Med. 2016, 16, 51. [Google Scholar] [CrossRef] [Green Version]
- Lian, Z.; Li, Y.; Gao, J.; Qu, K.; Li, J.; Hao, L.; Wu, S.; Zhu, H. A novel AMPK activator, WS070117, improves lipid metabolism discords in hamsters and HepG2 cells. Lipids Health Dis. 2011, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- McGee, S.L.; Howlett, K.; Starkie, R.L.; Cameron-Smith, D.; Kemp, B.E.; Hargreaves, M. Exercise increases nuclear AMPK 2 in human skeletal muscle. Diabetes 2003, 52, 926–928. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.-J.; Zou, H.; Zheng, J.-N.; Zou, T.-T.; Vitale, A.; Miele, L.; Van Poucke, S.; Liu, W.-Y.; Shen, S.; Zhang, D.-C.; et al. Serum alkaline phosphatase, a risk factor for non-alcoholic fatty liver, but only for women in their 30s and 40s: Evidence from a large cohort study. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 1–8. [Google Scholar] [CrossRef]
- Matsushita, M.; Otani, K.; Sakamoto, Y.; Arai, T.; Yukimasa, N.; Muramoto, Y.; Komoda, T. Increase in alkaline phosphatase activity after high-fat meal ingestion is correlated to the amount of ABH substances in saliva. Rinsho Byori. Jpn. J. Clin. Pathol. 2015, 63, 543–547. [Google Scholar]
- Chen, S.C.-C.; Tsai, S.P.; Jhao, J.-Y.; Jiang, W.-K.; Tsao, C.K.; Chang, L.-Y. Liver fat, hepatic enzymes, alkaline phosphatase and the risk of incident Type 2 diabetes: A prospective study of 132,377 adults. Sci. Rep. 2017, 7, 4649. [Google Scholar] [CrossRef]
- Zavisic, G.; Petricevic, S.; Radulovic, Z.; Begovic, J.; Golic, N.; Topisirovic, L.; Strahinic, I. Probiotic features of two oral Lactobacillus isolates. Braz. J. Microbiol. 2012, 43, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. BBA Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gierens, H.; Nauck, M.; Roth, M.; Schinker, R.; Schürmann, C.; Scharnagl, H.; Neuhaus, G.; Wieland, H.; März, W. Interleukin-6 stimulates LDL receptor gene expression via activation of sterol-responsive and Sp1 binding elements. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Frisdal, E.; Lesnik, P.; Olivier, M.; Robillard, P.; Chapman, M.J.; Huby, T.; Guerin, M.; Le Goff, W. Interleukin-6 protects human macrophages from cellular cholesterol accumulation and attenuates the proinflammatory response. J. Biol. Chem. 2011, 286, 30926–30936. [Google Scholar] [CrossRef] [Green Version]
- Voloshyna, I.; Reiss, A.B. The ABC transporters in lipid flux and atherosclerosis. Prog. Lipid Res. 2011, 50, 213–224. [Google Scholar] [CrossRef]
- Duong, M.; Collins, H.L.; Jin, W.; Zanotti, I.; Favari, E.; Rothblat, G.H. Relative contributions of ABCA1 and SR-BI to cholesterol efflux to serum from fibroblasts and macrophages. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, L.P.; Mates, J.M.; Cheplowitz, A.M.; Avila, C.L.; Zimmerer, J.M.; Yao, Z.; Maiseyeu, A.; Rajaram, M.V.S.; Robinson, J.M.; Anderson, C.L. Scavenger receptor B1, the HDL receptor, is expressed abundantly in liver sinusoidal endothelial cells. Sci. Rep. 2016, 6, 20646. [Google Scholar] [CrossRef] [Green Version]
- Michael, D.; Moss, J.; Calvente, D.L.; Garaiová, I.; Plummer, S.; Ramji, D.P. Lactobacillus plantarum CUL66 can impact cholesterol homeostasis in Caco-2 enterocytes. Benef. Microbes 2016, 7, 443–451. [Google Scholar] [CrossRef]
- Yoon, H.-S.; Ju, J.-H.; Kim, H.; Lee, J.; Park, H.; Ji, Y.; Shin, H.-K.; Do, M.-S.; Lee, J.-M.; Holzapfel, W. Lactobacillus rhamnosus BFE 5264 and Lactobacillus plantarum NR74 promote cholesterol excretion through the up-regulation of ABCG5/8 in Caco-2 cells. Probiot. Antimicrob. Proteins 2011, 3, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Njoku, D.B. Suppressive and pro-inflammatory roles for IL-4 in the pathogenesis of experimental drug-induced liver injury: A review. Expert Opin. Drug Metab. Toxicol. 2010, 6, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.C.; Menezes-Garcia, Z.; Henriques, M.C.C.; Soriani, F.M.; Pinho, V.; Faria, A.C.; Santiago, A.F.; Cara, D.C.; Souza, D.G.; Teixeira, M.M.; et al. Acute and sustained inflammation and metabolic dysfunction induced by high refined carbohydrate-containing diet in mice. Obesity 2013, 21, E396–E406. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, R.A.; Sheedfar, F.; Morrison, M.C.; Hommelberg, P.P.H.; Kor, D.; Kloosterhuis, N.J.; Gruben, N.; Youssef, S.A.; De Bruin, A.; Hofker, M.H.; et al. High-Fat diet induced obesity primes inflammation in adipose tissue prior to liver in C57BL/6j mice. Aging 2015, 7, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, K.; Pontillo, A.; Di Palo, C.; Giugliano, G.; Masella, M.; Marfella, R.; Giugliano, D. Effect of weight loss and lifestyle changes on vascular inflammatory markers in obese women: A randomized trial. JAMA 2003, 289, 1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | Normal Diet (ND) | High-Fat Diet (HFD) | ||
|---|---|---|---|---|
| g/kg | % | g/kg | % | |
| Crude protein | 192.11 | 19.2 | 144.08 | 14.4 |
| Crude fat | 40.80 | 4.1 | 280.60 | 28.0 |
| Crude fiber | 60.74 | 6.1 | 45.56 | 4.6 |
| Crude ash | 58.55 | 5.9 | 43.91 | 4.4 |
| Moisture | 113.43 | 11.3 | 85.07 | 8.5 |
| Nitrogen-free extractives | 534.37 | 53.4 | 400.78 | 40.1 |
| Total | 1000.00 | 100.0 | 1000.00 | 100.0 |
| kcal/kg | % | kcal/kg | % | |
| Fat | 367 | 11.0 | 2525 | 54.0 |
| Protein | 768 | 24.0 | 576 | 12.3 |
| Carbohydrates | 2091 | 65.0 | 1575 | 33.7 |
| Total | 3226 | 100.0 | 4676 | 100.0 |
| Target Genes | Primer Sequences (5′–3′) | NCBI Accession No. |
|---|---|---|
| GAPDH | F: CCA TCC CAG ACC CCA TAA C | NM_017008.4 |
| R: GCA GCG AAC TTT ATT GAT GG | ||
| SCD-1 | F: CAC TGG TGC CCT GGT ACT GCT | NM_139192.2 |
| R: GGA TGT TCT CCC GAG ATT GAA | ||
| IL-6 | F: TAG TCC TTC CTA CCC CAA CTT C | NM_012589.2 |
| R: GCC GAG TAG ACC TCA TAG TGA C | ||
| ABCG5 | F: TGT GAC CCT GGC ATC TAT | NM_053754.2 |
| R: ATC ATT GGA CCA GTT CAG T | ||
| ABCG8 | F: GAT GCT GGC TAT CAT AGG GAG C | NM_130414.2 |
| R: TCT CTG CCT GTG ATA ACG TCG A | ||
| SR-B1 | F: CCC AGA AGA CAC CAC GA | NM_031541.1 |
| R: GTG TGG ACA GTG TGA CAT CT | ||
| LDL-R | F: TGG TGA CCG AGG ACA TCC AG | NM_175762.2 |
| R: GTG GAG TTT GGA ATC AAA CCC AAT AG | ||
| ABCA1 | F: CAG CAA CTA CAG TGG CGG TAA CA | NM_178095.2 |
| R: AAT GCT TAG GGC ACA ATT CCA CA | ||
| APO A1 | F: GGC AGA GAC TAT GTG TCC CAG TTT | NM_012738.1 |
| R: TTG AAC CCA GAG TGT CCC AGT T | ||
| AMPKα1 | F: GGG ATC CAT CAG CAA CTA TCG | NM_019142.2 |
| R: GGG AGG TCA CGG ATG AGG TA | ||
| AMPKα2 | F: CAT TTG TGC AAG GCC CCT AGT | NM_023991.1 |
| R: GAC TGT TGG TAT CTG CCT GTT TCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lew, L.-C.; Hor, Y.-Y.; Jaafar, M.-H.; Lau, A.-S.-Y.; Lee, B.-K.; Chuah, L.-O.; Yap, K.-P.; Azlan, A.; Azzam, G.; Choi, S.-B.; et al. Lactobacillus Strains Alleviated Hyperlipidemia and Liver Steatosis in Aging Rats via Activation of AMPK. Int. J. Mol. Sci. 2020, 21, 5872. https://doi.org/10.3390/ijms21165872
Lew L-C, Hor Y-Y, Jaafar M-H, Lau A-S-Y, Lee B-K, Chuah L-O, Yap K-P, Azlan A, Azzam G, Choi S-B, et al. Lactobacillus Strains Alleviated Hyperlipidemia and Liver Steatosis in Aging Rats via Activation of AMPK. International Journal of Molecular Sciences. 2020; 21(16):5872. https://doi.org/10.3390/ijms21165872
Chicago/Turabian StyleLew, Lee-Ching, Yan-Yan Hor, Mohamad-Hafis Jaafar, Amy-Sie-Yik Lau, Boon-Kiat Lee, Li-Oon Chuah, Kien-Pong Yap, Azali Azlan, Ghows Azzam, Sy-Bing Choi, and et al. 2020. "Lactobacillus Strains Alleviated Hyperlipidemia and Liver Steatosis in Aging Rats via Activation of AMPK" International Journal of Molecular Sciences 21, no. 16: 5872. https://doi.org/10.3390/ijms21165872