Multicopy Suppressor Analysis of Strains Lacking Cytoplasmic Peptidyl-Prolyl cis/trans Isomerases Identifies Three New PPIase Activities in Escherichia coli That Includes the DksA Transcription Factor

 and
and
Abstract
:1. Introduction
2. Results
2.1. Δ6ppi Mutant Bacteria Exhibit the Sensitivity towards Antibiotics, Membrane-Destabilizing Factors and the DNA-Damaging Agent Nalidixic Acid
2.2. Factors Limiting for the Viability of Δ6ppi Mutant Bacteria
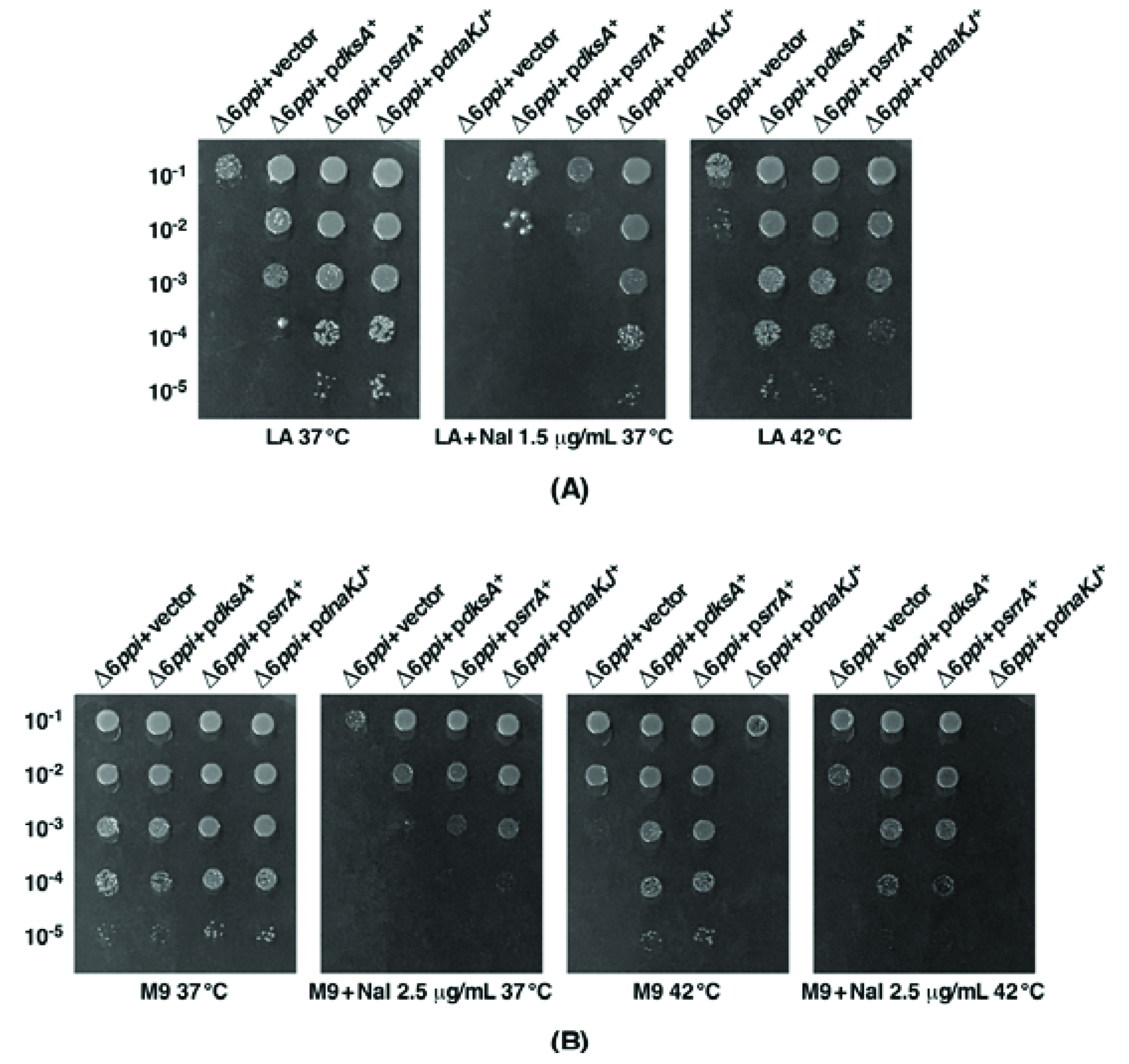
2.3. Overexpression of Either the srrA Gene or the dksA Gene Suppresses the Sensitivity of Δ6ppi Bacteria to Nal
2.4. The Essentiality of DksA, SrrA, MetL and Hda for the Viability of Δ6ppi Bacteria and Synthetic Growth Defects with Δcmk and ΔhchA
2.5. Overexpression of Either the dksA Gene or the cmk Gene or the metL Gene Also Restore the Growth of a Δ(dnaK dnaJ tig) Strain
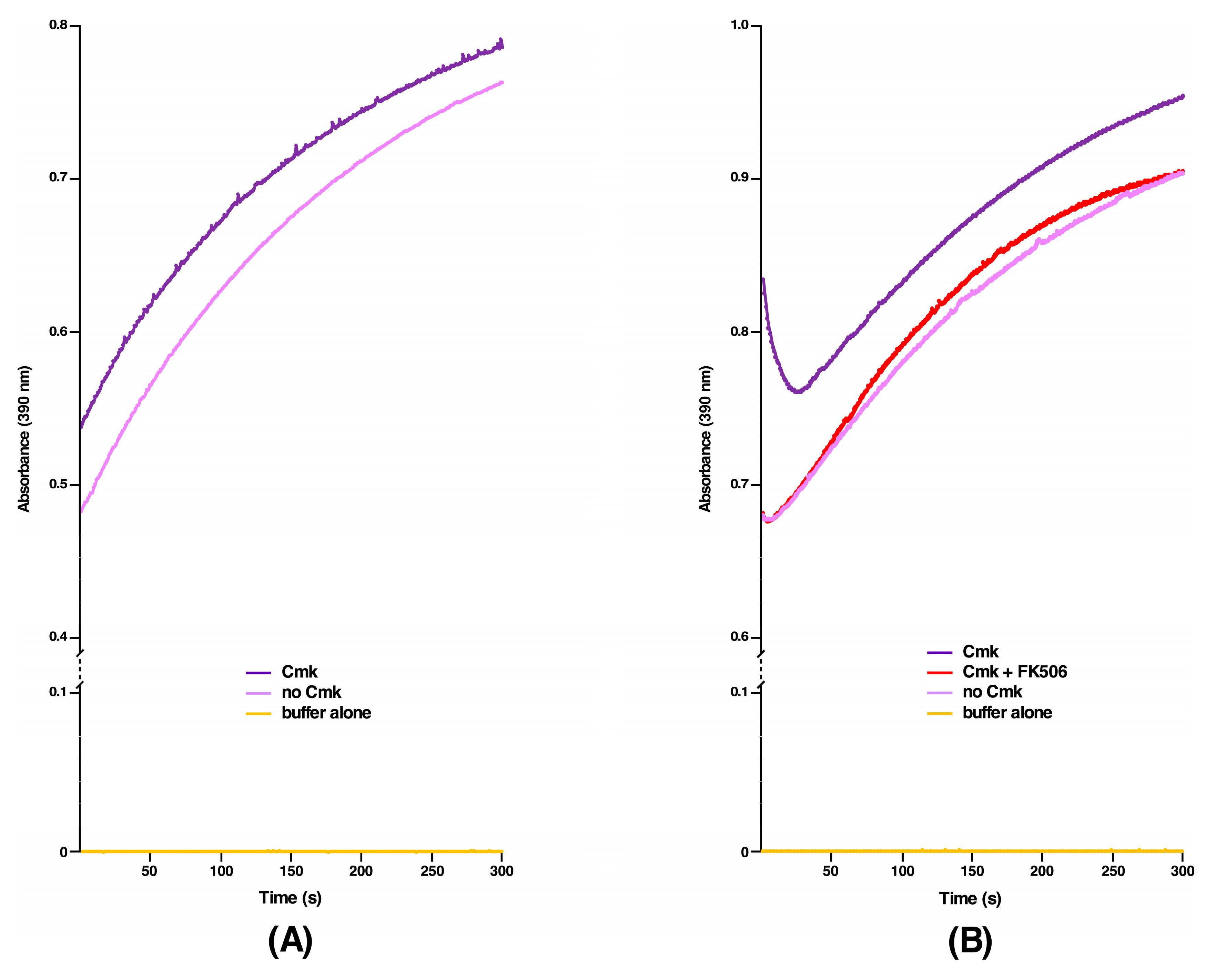
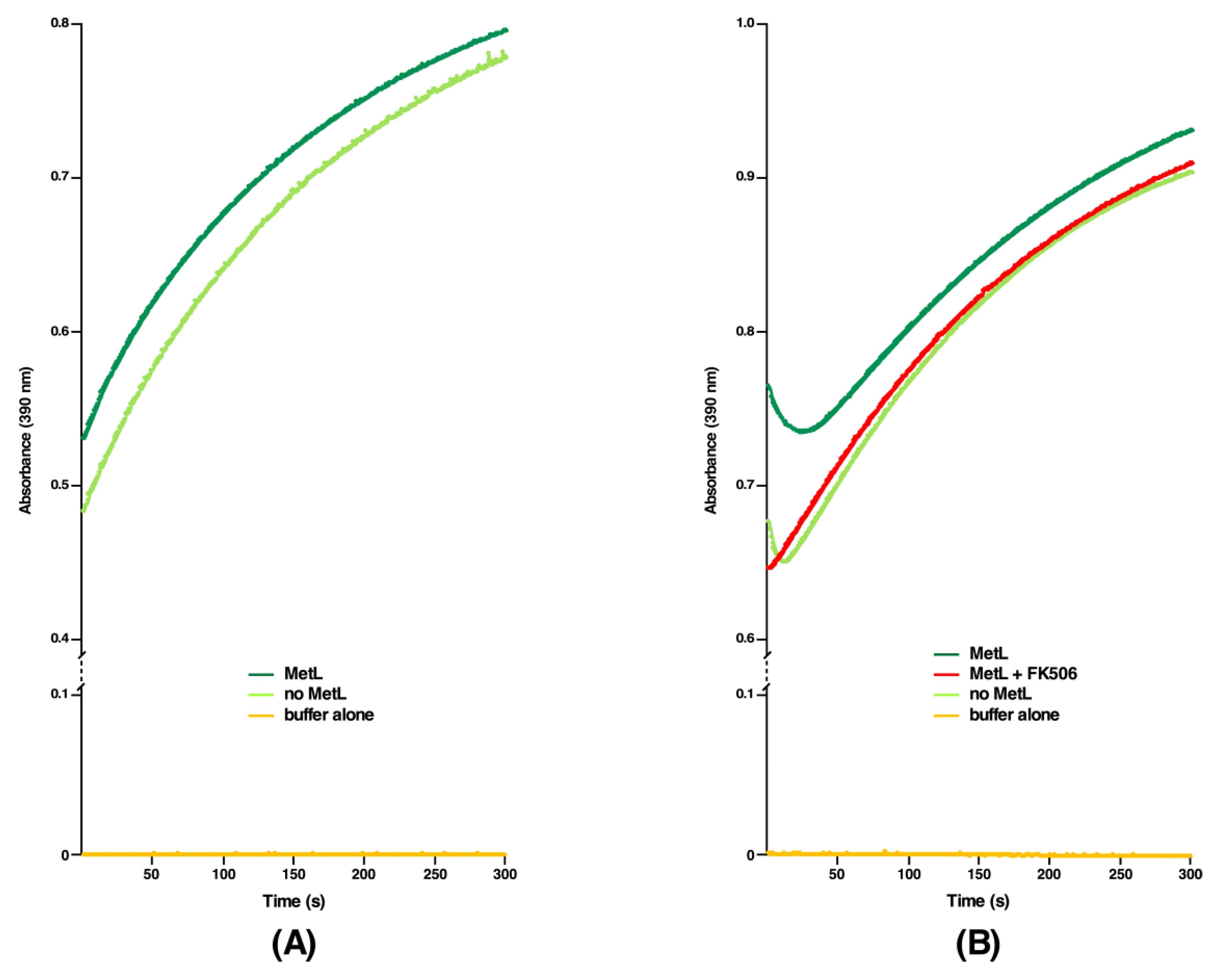
2.6. DksA, Cmk and MetL Exhibit the PPIase Activity, Explaining Their Mode of Suppression
2.7. The PPIase Activity of DksA, Cmk and MetL Is Inhibited by FK506
2.8. DksA, Cmk and MetL Can Catalyze the PPIase-Dependent Refolding of RNase T1
2.9. Identification of Amino Acid Residues That Are Critical for the PPIase Activity of DksA and Its Multicopy Suppression
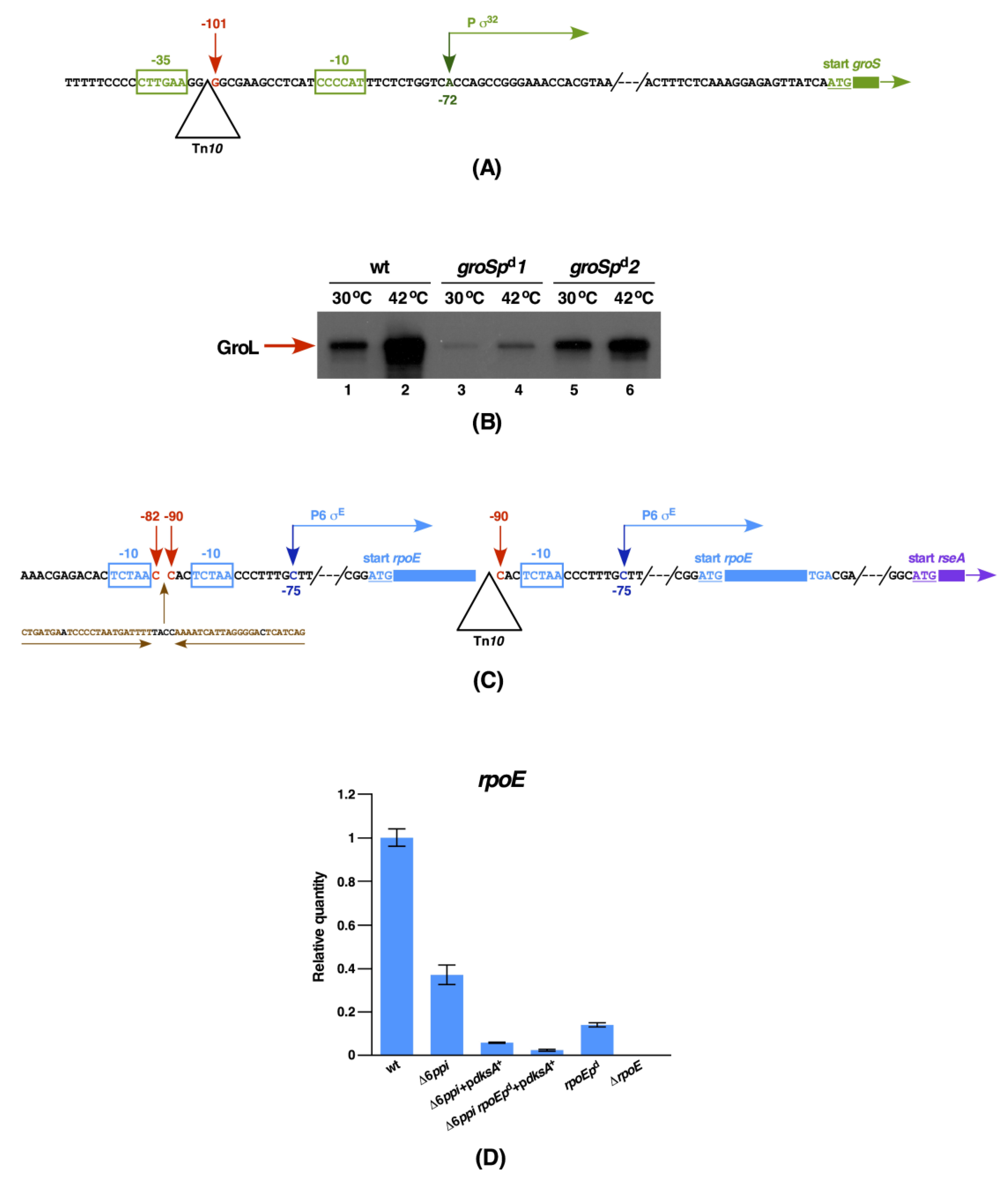
2.10. The DksA-Mediated Multicopy Suppression of Either Δ6ppi or ΔdnaK/J Mutant Bacteria Requires the Wild-Type Expression of GroL/S and RpoE Essential Proteins
2.11. The DksA-Mediated Multicopy Suppression Requires Cell Envelope Homeostasis, the Genome Integrity, the Ribosome Assembly, Translocation of Folded Proteins and Factors Combating Oxidative Stress
2.12. The ppk Gene Encoding Polyphosphate Kinase Is Not Required for the DksA-Mediated Multicopy Suppression of a Δ6ppi Strain and Only a Marginal Requirement for a Δ(dnaK/J) Derivative
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids and Media
4.2. Generation of Null Mutations in Various Genes, Whose Products in the High Dosage Suppress Growth Defects of Δ6ppi Strains and the Construction of Δ(dnaK/J tig) Deletion Derivatives
4.3. The Identification of Multicopy Suppressors, Whose Overexpression Restores the Growth of Either Δ6ppi or Δ(dnaK/J tig) Derivatives under Non-Permissive Growth Conditions
4.4. The Isolation of Chromosomal Transposon Insertion Mutations That Prevent the Multicopy Suppression by the dksA Gene
4.5. Cloning of Various Genes for Complementation Studies
4.6. RNA Purification and qRT-PCR Analysis
4.7. Protein Purification of Wild-Type and dksA Mutants
4.8. The PPIase Assay and the PPIase-Dependent Refolding of RNase T1
4.9. β-Galactosidase Activity Assay and Measurement of GroL Levels
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PPI | peptidyl-prolyl cis/trans isomerase |
| FKBP | FK506-binding protein |
| RNAP | RNA polymerase |
References
- Klein, G.; Wojtkiewicz, P.; Biernacka, D.; Stupak, A.; Gorzelak, P.; Raina, S. Identification of substrates of cytoplasmic peptidyl-prolyl cis/trans isomerases and their collective essentiality in Escherichia coli. Int. J. Mol. Sci. 2020, 21, 4212. [Google Scholar] [CrossRef] [PubMed]
- Okumura, M.; Kadokura, H.; Hashimoto, S.; Yutani, K.; Kanemura, S.; Hikima, T.; Hidaka, Y.; Ito, L.; Shiba, K.; Masui, S.; et al. Inhibition of the functional interplay between endoplasmic reticulum (ER) oxidoreduclin-1α (Ero1α) and protein-disulfide isomerase (PDI) by the endocrine disruptor bisphenol A. J. Biol. Chem. 2014, 289, 27004–27018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, G.; Aumüller, T. Regulation of peptide bond cis/trans isomerization by enzyme catalysis and its implication in physiological processes. Rev. Physiol. Biochem. Pharmacol. 2003, 148, 105–150. [Google Scholar] [CrossRef] [PubMed]
- Grathwohl, C.; Wüthrich, K. The X-Pro peptide bond as an NMR probe for conformational studies of flexible linear peptides. Biopolymers 1976, 15, 2025–2041. [Google Scholar] [CrossRef]
- Cheng, H.N.; Bovey, F.A. Cis-trans equilibrium and kinetic studies of acetyl-l-proline and glycyl-l-proline. Biopolymers 1977, 16, 1465–1472. [Google Scholar] [CrossRef]
- Stewart, D.E.; Sarkar, A.; Wampler, J.E. Occurrence and role of cis peptide bonds in protein structures. J. Mol. Biol. 1990, 214, 253–260. [Google Scholar] [CrossRef]
- Schmidpeter, P.A.M.; Koch, J.R.; Schmid, F.X. Control of protein function by prolyl isomerization. Biochim. Biophys. Acta 2015, 1850, 1973–1982. [Google Scholar] [CrossRef]
- Lang, K.; Schmid, F.X.; Fischer, G. Catalysis of protein folding by prolyl isomerase. Nature 1987, 329, 268–270. [Google Scholar] [CrossRef]
- Siekierka, J.J.; Hung, S.H.; Poe, M.; Lin, C.S.; Sigal, N.H. A cytosolic binding protein for the immunosuppressant FK506 has peptidyl-prolyl isomerase activity but is distinct from cyclophilin. Nature 1989, 341, 755–757. [Google Scholar] [CrossRef]
- Rahfeld, J.U.; Schierhorn, A.; Mann, K.; Fischer, G. A novel peptidyl-prolyl cis/trans isomerase from Escherichia coli. FEBS Lett. 1994, 343, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Myka, K.K.; Küsters, K.; Washburn, R.; Gottesman, M.E. DksA-RNA polymerase interactions support new origin formation and DNA repair in Escherichia coli. Mol. Microbiol. 2019, 111, 1382–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, M.; Zhao, X.; Drlica, K. Lethal fragmentation of bacterial chromosomes mediated by DNA gyrase and quinolones. Mol. Microbiol. 2006, 61, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, M.; Ara, T.; Arifuzzaman, M.; Ioka-Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Mori, H. Complete set of ORF clones of Escherichia coli ASKA library (a complete set of E. coli K-12 ORF archive): Unique resources for biological research. DNA Res. 2005, 12, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, G.; Kobylak, N.; Lindner, B.; Stupak, A.; Raina, S. Assembly of lipopolysaccharide in Escherichia coli requires the essential LapB heat shock protein. J. Biol. Chem. 2014, 289, 14829–14853. [Google Scholar] [CrossRef] [Green Version]
- Klein, G.; Stupak, A.; Biernacka, D.; Wojtkiewicz, P.; Lindner, B.; Raina, S. Multiple transcriptional factors regulate transcription of the rpoE gene in Escherichia coli under different growth conditions and when the lipopolysaccharide biosynthesis is defective. J. Biol. Chem. 2016, 291, 22999–23019. [Google Scholar] [CrossRef] [Green Version]
- Missiakas, D.; Georgopoulos, C.; Raina, S. Identification and characterization of the Escherichia coli gene dsbB, whose product is involved in the formation of disulfide bonds in vivo. Proc. Natl. Acad. Sci. USA 1993, 90, 7084–7088. [Google Scholar] [CrossRef] [Green Version]
- Klein, G.; Lindner, B.; Brade, H.; Raina, S. Molecular basis of lipopolysaccharide heterogeneity in Escherichia coli: Envelope stress-responsive regulators control the incorporation of glycoforms with a third 3-deoxy-α-D-manno-oct-2-ulosonic acid and rhamnose. J. Biol. Chem. 2011, 286, 42787–42807. [Google Scholar] [CrossRef] [Green Version]
- Raina, S.; Missiakas, D.; Baird, L.; Kumar, S.; Georgopoulos, C. Identification and transcriptional analysis of the Escherichia coli htrE operon which is homologous to Pap and related pilin operons. J. Bacteriol. 1993, 175, 5009–5021. [Google Scholar] [CrossRef] [Green Version]
- Teter, S.A.; Houry, W.A.; Ang, D.; Tradler, T.; Rockabrand, D.; Fischer, G.; Blum, P.; Georgopoulos, C.; Hartl, F.U. Polypeptide flux through bacterial Hsp70: DnaK cooperates with trigger factor in chaperoning nascent chains. Cell 1999, 97, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Deuerling, E.; Schulze-Specking, A.; Tomoyasu, T.; Mogk, A.; Bukau, B. Trigger factor and DnaK cooperate in folding of newly synthesized proteins. Nature 1999, 400, 693–696. [Google Scholar] [CrossRef]
- Geitner, A.J.; Weininger, U.; Paulsen, H.; Balbach, J.; Kovermann, M. Structure-based insights into the dynamics and function of two-domain SlpA from Escherichia coli. Biochemistry 2017, 56, 6533–6543. [Google Scholar] [CrossRef] [PubMed]
- Hennig, L.; Christner, C.; Kipping, M.; Schelbert, B.; Rücknagel, K.P.; Grabley, S.; Küllertz, G.; Fischer, G. Selective inactivation of parvulin-like peptidyl-prolyl cis/trans isomerases by juglone. Biochemistry 1998, 37, 5953–5960. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Wittmann-Liebold, B.; Lang, K.; Kiefhaber, T.; Schmid, F.X. Cyclophilin and peptidyl-prolyl cis-trans isomerase are probably identical proteins. Nature 1989, 337, 476–478. [Google Scholar] [CrossRef] [PubMed]
- Schmid, F.X.; Frech, C.; Scholz, C.; Walter, S. Catalyzed and assisted protein folding of ribonuclease T1. Biol. Chem. 1996, 377, 417–424. [Google Scholar] [PubMed]
- Gourse, R.L.; Chen, A.Y.; Gopalkrishnan, S.; Sanchez-Vazquez, P.; Myers, A.; Ross, W. Transcriptional responses to ppGpp and DksA. Annu. Rev. Microbiol. 2018, 72, 163–184. [Google Scholar] [CrossRef]
- Paul, B.J.; Barker, M.M.; Ross, W.; Schneider, D.A.; Webb, C.; Foster, J.W.; Gourse, R.L. DksA: A critical component of the transcription initiation machinery that potentiates the regulation of rRNA promoters by ppGpp and the initiating NTP. Cell 2004, 118, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Lennon, C.W.; Ross, W.; Gourse, R.L. Role of the coiled-coil tip of Escherichia coli DksA in promoter control. J. Mol. Biol. 2012, 416, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Parshin, A.; Shiver, A.L.; Lee, J.; Ozerova, M.; Schneidman-Duhovny, D.; Gross, C.A.; Borukhov, S. DksA regulates RNA polymerase in Escherichia coli through a network of interactions in the secondary channel that includes sequence insertion 1. Proc. Natl. Acad. Sci. USA 2015, 112, E6862–E6871. [Google Scholar] [CrossRef] [Green Version]
- Chandrangsu, P.; Wang, L.; Choi, S.H.; Gourse, R.L. Suppression of a dnaKJ deletion by multicopy dksA results from non-feedback-regulated transcripts that originate upstream of the major dksA promoter. J. Bacteriol. 2012, 194, 1437–1446. [Google Scholar] [CrossRef] [Green Version]
- Kang, P.J.; Craig, E.A. Identification and characterization of a new Escherichia coli gene that is a dosage-dependent suppressor for a dnaK deletion mutation. J. Bacteriol. 1990, 172, 2055–2064. [Google Scholar] [CrossRef] [Green Version]
- Lipinska, B.; Sharma, S.; Georgopoulos, C. Sequence analysis and regulation of the htrA gene of Escherichia coli: A σ32-independent mechanism of heat-inducible transcription. Nucleic Acids Res. 1988, 16, 10053–10067. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Lipinska, B.; Raina, S.; Georgopoulos, C. Identification of the Escherichia coli sohB gene, a multicopy suppressor of the HtrA (DegP) null phenotype. J. Bacteriol. 1991, 173, 5763–5770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dartigalongue, C.; Missiakas, D.; Raina, S. Characterization of the Escherichia coli σE regulon. J. Biol. Chem. 2001, 276, 20866–20875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, B.S.; Gabbert, K.K.; Kranz, R.G. The temperature-sensitive growth and survival phenotypes of Escherichia coli cydDC and cydAB strains are due to deficiencies in cytochrome bd and are corrected by exogenous catalase and reducing agents. J. Bacteriol. 1996, 178, 6348–6351. [Google Scholar] [CrossRef] [Green Version]
- Kusukawa, N.; Yura, T. Heat shock protein GroE of Escherichia coli: Key protective roles against thermal stress. Genes Dev. 1988, 2, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Kofron, J.L.; Kuzmič, P.; Kishore, V.; Colón-Bonilla, E.; Rich, D.H. Determination of kinetic constants for peptidyl prolyl cis-trans isomerases by an improved spectrophotometric assay. Biochemistry 1991, 30, 6127–6134. [Google Scholar] [CrossRef]
- Park, S.T.; Aldape, R.A.; Futer, O.; DeCenzo, M.T.; Livingston, D.J. PPIase catalysis by human FK506-binding protein proceeds through a conformational twist mechanism. J. Biol. Chem. 1992, 267, 3316–3324. [Google Scholar]
- Fujimitsu, K.; Su′etsugu, M.; Yamaguchi, Y.; Mazda, K.; Fu, N.; Kawakami, H.; Katayama, T. Modes of overinitiation, dnaA gene expression, and inhibition of cell division in a novel cold-sensitive hda mutant of Escherichia coli. J. Bacteriol. 2008, 190, 5368–5381. [Google Scholar] [CrossRef] [Green Version]
- Falcoz-Kelly, F.; van Rapenbusch, R.; Cohen, G.N. The methionine-repressible homoserine dehydrogenase and aspartokinase activities of Escherichia coli K12. Preparation of the homogeneous protein catalyzing the two activities. Molecular weight of the native enzyme and of its subunits. Eur. J. Biochem. 1969, 8, 146–152. [Google Scholar] [CrossRef]
- Anglès, F.; Castanié-Cornet, M.P.; Slama, N.; Dinclaux, M.; Cirinesi, A.M.; Portais, J.C.; Létisse, F.; Genevaux, P. Mutilevel interaction of the DnaK/DnaJ(HSP70/HSP40) stress-responsive chaperone machine with the central metabolism. Sci. Rep. 2017, 7, 41341. [Google Scholar] [CrossRef]
- Laptenko, O.; Lee, J.; Lomakin, I.; Borukhov, S. Transcript cleavage factors GreA and GreB act as transient catalytic components of RNA polymerase. EMBO J. 2003, 22, 6322–6334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opalka, N.; Chlenov, M.; Chacon, P.; Rice, W.J.; Wriggers, W.; Darst, S.A. Structure and function of the transcription elongation factor GreG bound to bacterial RNA polymerase. Cell 2003, 114, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Lamour, V.; Rutherford, S.T.; Kuznedelov, K.; Ramagopal, U.A.; Gourse, R.L.; Severinov, K.; Darst, S.A. Crystal structure of Escherichia coli Rnk, a new RNA polymerase-interacting protein. J. Mol. Biol. 2008, 383, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, G.; Raina, S. Small regulatory bacterial RNAs regulating the envelope stress response. Biochem. Soc. Trans. 2017, 45, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernadac, A.; Gavioli, M.; Lazzaroni, J.C.; Raina, S.; Lloubès, R. Escherichia coli tol-pal mutants form outer membrane vesicles. J. Bacteriol. 1998, 180, 4872–4878. [Google Scholar] [CrossRef] [Green Version]
- Vinés, E.D.; Marolda, C.L.; Balachandran, A.; Valvano, M.A. Defective O-antigen polymerization in tolA and pal mutants of Escherichia coli in response to extracytoplasmic stress. J. Bacteriol. 2005, 187, 3359–3368. [Google Scholar] [CrossRef] [Green Version]
- Missiakas, D.; Raina, S. Signal transduction pathways in response to protein misfolding in the extracytoplasmic compartments of E. coli: Role of two new phosphoprotein phosphatases PrpA and PrpB. EMBO J. 1997, 16, 1670–1685. [Google Scholar] [CrossRef] [Green Version]
- Dartigalongue, C.; Raina, S. A new heat-shock gene, ppiD, encodes a peptidyl-prolyl isomerase required for folding of outer membrane proteins in Escherichia coli. EMBO J. 1998, 17, 3968–3980. [Google Scholar] [CrossRef] [Green Version]
- Fayet, O.; Ziegelhoffer, T.; Georgopoulos, C. The groES and groEL heat shock gene products of Escherichia coli are essential for bacterial growth at all temperatures. J. Bacteriol. 1989, 171, 1379–1385. [Google Scholar] [CrossRef] [Green Version]
- Shewmaker, F.; Kerner, M.J.; Hayer-Hartl, M.; Klein, G.; Georgopoulos, C.; Landry, S.J. A mobile loop order-disorder transition modulates the speed of chaperonin cycling. Protein Sci. 2004, 13, 2139–2148. [Google Scholar] [CrossRef] [Green Version]
- Wimberly, H.; Shee, C.; Thornton, P.C.; Sivaramakrishnan, P.; Rosenberg, S.M.; Hastings, P.J. R-loops and nicks initiate DNA breakage and genome instability in non-growing Escherichia coli. Nat. Commun. 2013, 4, 2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cléon, F.; Habersetzer, J.; Alcock, F.; Kneuper, H.; Stansfeld, P.J.; Basit, H.; Wallace, M.I.; Berks, B.C.; Palmer, T. The TatC component of the twin-arginine protein translocase functions as an obligate oligomer. Mol. Microbiol. 2015, 98, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graubner, W.; Schierhorn, A.; Brüser, T. DnaK plays a pivotal role in Tat targeting of CueO and functions beside SlyD as a general Tat signal binding chaperone. J. Biol. Chem. 2007, 282, 7116–7124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Rodríguez, R.; Fisher, A.C.; Perlmutter, J.D.; Hicks, M.G.; Chanal, A.; Santini, C.L.; Wu, L.F.; Palmer, T.; DeLisa, M.P. An essential role for the DnaK molecular chaperone in stabilizing over-expressed substrate proteins of the bacterial twin-arginine translocation pathway. J. Mol. Biol. 2007, 367, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, R.; Oman, K.; Shoji, S.; Bundschuh, R.; Fredrick, K. The conserved GTPase LepA contributes mainly to translation initiation in Escherichia coli. Nucleic Acids Res. 2014, 42, 13370–13383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, M.R.; Fredrick, K. Roles of elusive translational GTPases come to light and inform on the process of ribosome biogenesis in bacteria. Mol. Microbiol. 2018, 107, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.J. Interactions between DksA and stress-responsive alternative sigma factors control inorganic polyphosphate accumulation in Escherichia coli. J. Bacteriol. 2020. [CrossRef]
- Morohoshi, T.; Maruo, T.; Shirai, Y.; Kato, J.; Ikeda, T.; Takiguchi, N.; Ohtake, H.; Kuroda, A. Accumulation of inorganic polyphosphate in phoU mutants of Escherichia coli and Synechocystis sp. strain PCC6803. Appl. Environ. Microbiol. 2002, 68, 4107–4110. [Google Scholar] [CrossRef] [Green Version]
- Bass, S.; Gu, Q.; Christen, A. Multicopy suppressors of prc mutant Escherichia coli include two HtrA (DegP) protease homologs (HhoAB), DksA, and a truncated R1pA. J. Bacteriol. 1996, 178, 1154–1161. [Google Scholar] [CrossRef] [Green Version]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [Green Version]
- Raina, S.; Missiakas, D.; Georgopoulos, C. The rpoE gene encoding the σE (σ24) heat shock sigma factor of Escherichia coli. EMBO J. 1995, 14, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Simons, R.W.; Houman, F.; Kleckner, N. Improved single and multicopy lac-based cloning vectors for protein and operon fusions. Gene 1987, 53, 85–96. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Ethanol 4% | SDS 1% | Deoxycholate 0.75% | Vancomycin 60 μg/mL | Erythromycin 12 μg/mL | Tetracycline 1.5 μg/mL | Kanamycin 2 μg/mL |
|---|---|---|---|---|---|---|---|
| wild type | 8 × 108 | 3 × 108 | 6 × 108 | 2 × 108 | 6 × 108 | 3 × 108 | 7 × 108 |
| Δ6ppi | 2 × 102 | 3 × 102 | - | - | - | 3 × 102 | 1 × 102 |
| Growth conditions | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene | LA 23 °C | LA 30 °C | LA 37 °C | LA 42 °C | LA 43.5 °C | V/Et/Er | M9 42 °C | Function |
| metL | + a | + | + | + | + | + V/Er | + sc | aspartokinase |
| dksA | - | - | ± b | + | + | - c | + | transcription |
| srrA | - | + sc | + | + | + | + Et | + | transcription |
| yjfN | - | + sc | + | + | + | + Er | + | proteolytic |
| cmk | + | + | + | + | ± | + Et/Er/V | + | DNA synthesis |
| hda | + | + | + sc | - | - | + Et/Er | + | replication |
| diaA | + | + | + | - | - | + Et/Er | + | replication |
| dnaK/J | + | + | + | + | ± | NT | - | chaperone |
| groL/S | - | ± sc | + | + | ± | NT | + | chaperonin |
| mqsA | - | + | + | + | ± | + Er | + | antitoxin |
| yceD | - | ± | + | - | - | + V | ± | stress response |
| ariR | + | + | + | + | + | + Er | + sc | stress response |
| pepA | + | + | + | + | + sc | - | ± | proteolytic |
| bcr | + | + | + | - | - | + Et/Er | - | peptide transport |
| cspC | - | ± | + | + sc | + sc | - | + | stress response |
| hha | - | - | - | - | - | + Er | + | transcription with mqs |
| mppA | + | + | ± sc | - | - | + V/Er | - | murein binding |
| ycaD | + | + | + sc | - | - | + V/Er | - | transport |
| murI | + sc | ± | + | - | - | + Et/Er/V | - | peptidoglycan |
| nudE | - | + | + | - | - | + Et/Er | - | nudix |
| nudJ | + | + | ± | - | - | - | - | nudix |
| yigB | + | + | - | - | - | - | - | riboflavin |
| ycjW | - | - | + | - | - | + Er | - | transcription |
| gadW | ± sc | ± | ± sc | - | - | + Er | - | acid resistance |
| ydgC | + | + | + | ± | - | + Er | - | GlpM family |
| yhaM | - | - | + | - | - | - | - | cysteine detoxification |
| preT | - | - | + | - | - | - | - | pyrimidine metabolism |
| Number of Transductants in Δ6ppi Strains Obtained Either in the Presence or Absence of Covering Wild-Type Plasmid-Born Gene | ||
|---|---|---|
| Gene | BW25113 LA/M9 + CAA 37 °C | MC4100 LA/M9 + CAA 37 °C |
| Δhda | 9 | 6 |
| Δhda +phda+ | 1234 | 1146 |
| ΔdksA | 8 * | small non-viable colonies |
| ΔdksA+pdksA+ | 1436 | 1598 |
| srrA | 7 | 455 small colonies after 48 h, cold sensitive at 23 °C and 30 °C |
| srrA+psrrA+ | 1255 | 1376 |
| metL | 11 * | 23 |
| metL+pmetL+ | 1830 | 1941 |
| cmk | 136 small colonies after 48 h | 254 small colonies after 48 h |
| cmk+pcmk+ | 1575 | 945 |
| hchA | 312 small colony size | 380 after 48 h (however, viable on M9) |
| hchA+phchA+ | 1174 | 1470 |
| Number of Transductants with Selection for Tetracycline Resistance | ||
|---|---|---|
| Donor | Recipient | |
| BW25113 | Δ6ppi | |
| htrE::tet | 873 tetR | 912 tetR |
| htrE::tet dksA::cm | 940 tetR | 922 tetR |
| (870 cmR) | (0 cmR) | |
| Strain | Auxotrophy | rrnBP1 Activity | ΔdnaK/J | Δ(dnaK/J tig) | Δ6ppi | PPIase Activity |
|---|---|---|---|---|---|---|
| wild type DksA | +a | repressed | + | + | + | + |
| DksA D74N | -b | not repressed | -e | -e | - | reduced |
| DksA F82Y | ± f sc | repressed | -e | -e | - | highly reduced |
| DksA S83A | -c | weakly repressed | -e | -g | - | reduced |
| DksA L84A | -b | not repressed | -e | -e | - | reduced |
| DksA E85A | -c | weakly repressed | - | -g | -d | marginal reduction |
| Gene | Tn10 Position | ΔdnaK/J+pdksA+ | wt | Δ6ppi | ΔdksA | Function | |
|---|---|---|---|---|---|---|---|
| 40 °C | 42 °C | 43 °C | M9 37 °C | 43 °C | |||
| rpoEpd | −90 | - | - | +a sc | + | + sc | sigma factor |
| groSpd1 | −101 | - | - | - | + sc | - | chaperone |
| groSpd2 | −101 | - | - | + sc | + | + | chaperone |
| degP | 12/112 * | - | - | - | + | - Ts >39 °C | periplasmic protease |
| lepA | 702 | ± sc | - | ±b sc | + sc | -c | translational GTPase |
| ahpC | 94 | - | - | + | + sc | + | oxidative stress |
| rnhA | 59 | - | - | + | + sc | ± sc | ribonuclease HI |
| tatC | 23 | - | - | + | + | - sc | transport of folded proteins |
| tolA | 299 | ± sc | - | + | ± sc | ± vsc | cell envelope integrity |
| mrcB | 1186 | ± sc | - | + | + | + | peptidoglycan synthesis |
| oxyR | 518 | ± sc | - | + | + | + | oxidative stress regulator |
| cpxR | 474 | + | ± sc | + | + | + sc | envelope stress regulator |
| cydA | 20 | - | - | - | + | - | cytochrome d terminal oxidase |
| clsA | 533 | ± sc | - | + | + | + | cardiolipin synthase |
| ftsX | 166 | - | - | ± sc | ± sc | - | cell division |
| nudB | 369 | + | ± | + | + | ± sc | folate biosynthesis |
| envZ | 291 | ± sc | - | + | + | + | regulation of ompF/C expression |
| phoU | 655 | - | - | +sc | +sc | - vsc | Pi signalling |
| Δppk | deletion | + | - | + | + | + | polyphosphate kinase |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtkiewicz, P.; Biernacka, D.; Gorzelak, P.; Stupak, A.; Klein, G.; Raina, S. Multicopy Suppressor Analysis of Strains Lacking Cytoplasmic Peptidyl-Prolyl cis/trans Isomerases Identifies Three New PPIase Activities in Escherichia coli That Includes the DksA Transcription Factor. Int. J. Mol. Sci. 2020, 21, 5843. https://doi.org/10.3390/ijms21165843
Wojtkiewicz P, Biernacka D, Gorzelak P, Stupak A, Klein G, Raina S. Multicopy Suppressor Analysis of Strains Lacking Cytoplasmic Peptidyl-Prolyl cis/trans Isomerases Identifies Three New PPIase Activities in Escherichia coli That Includes the DksA Transcription Factor. International Journal of Molecular Sciences. 2020; 21(16):5843. https://doi.org/10.3390/ijms21165843
Chicago/Turabian StyleWojtkiewicz, Pawel, Daria Biernacka, Patrycja Gorzelak, Anna Stupak, Gracjana Klein, and Satish Raina. 2020. "Multicopy Suppressor Analysis of Strains Lacking Cytoplasmic Peptidyl-Prolyl cis/trans Isomerases Identifies Three New PPIase Activities in Escherichia coli That Includes the DksA Transcription Factor" International Journal of Molecular Sciences 21, no. 16: 5843. https://doi.org/10.3390/ijms21165843