Neuroimmune Response in Natural Preclinical Scrapie after Dexamethasone Treatment


Abstract
:1. Introduction
2. Results
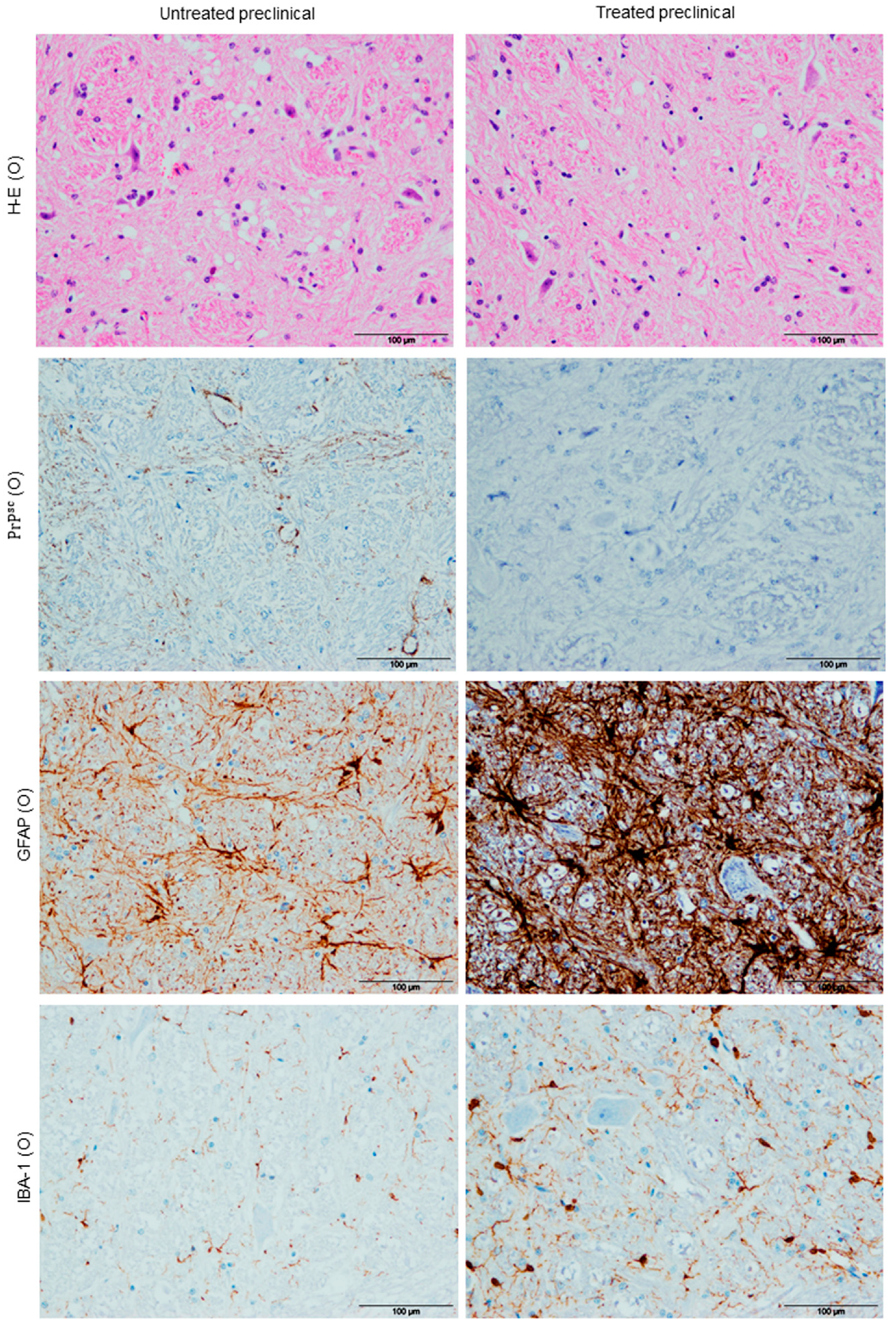
2.1. Histopathological Findings (H-E)
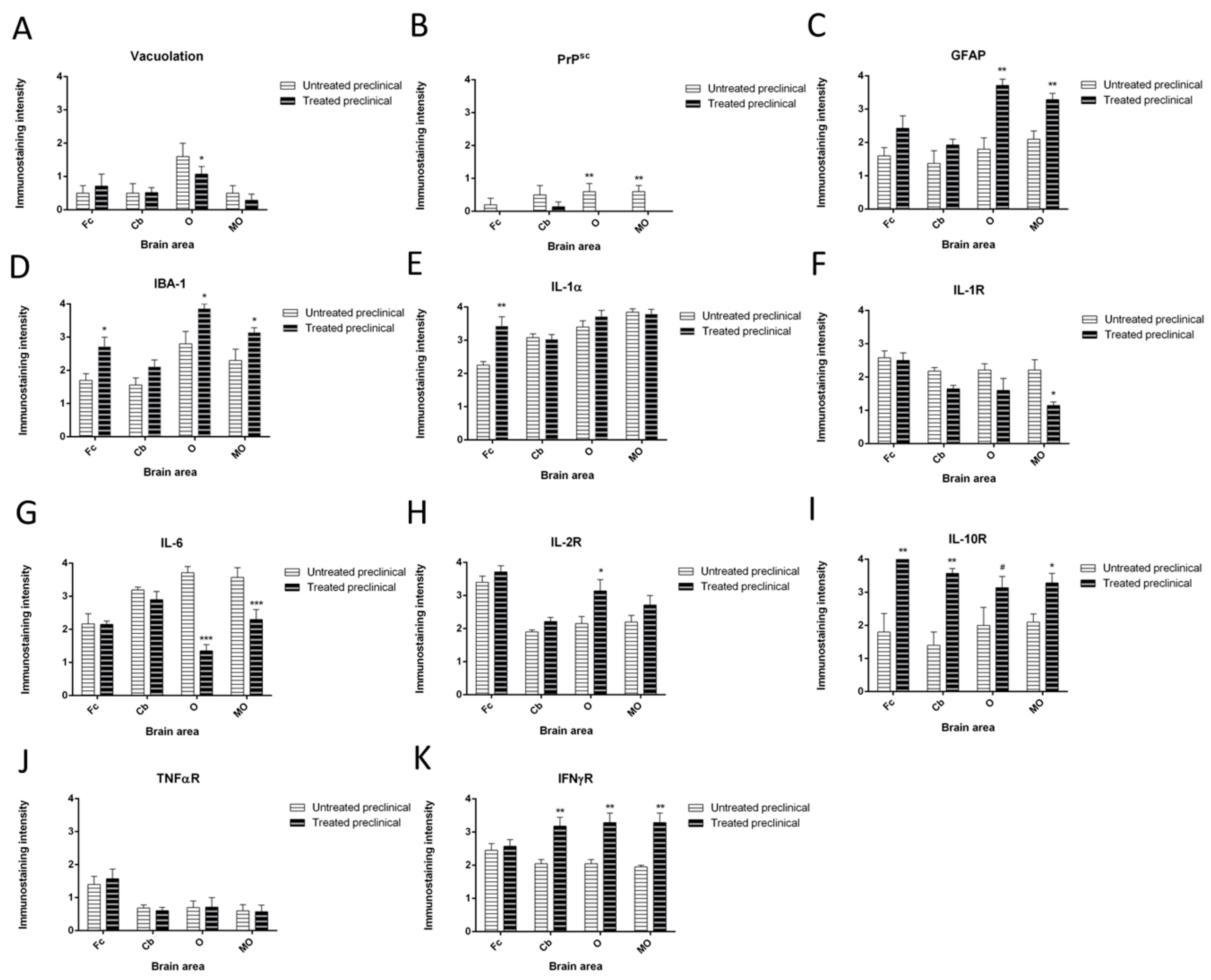
Vacuolation
2.2. Immunohistochemical Findings (Ihc)
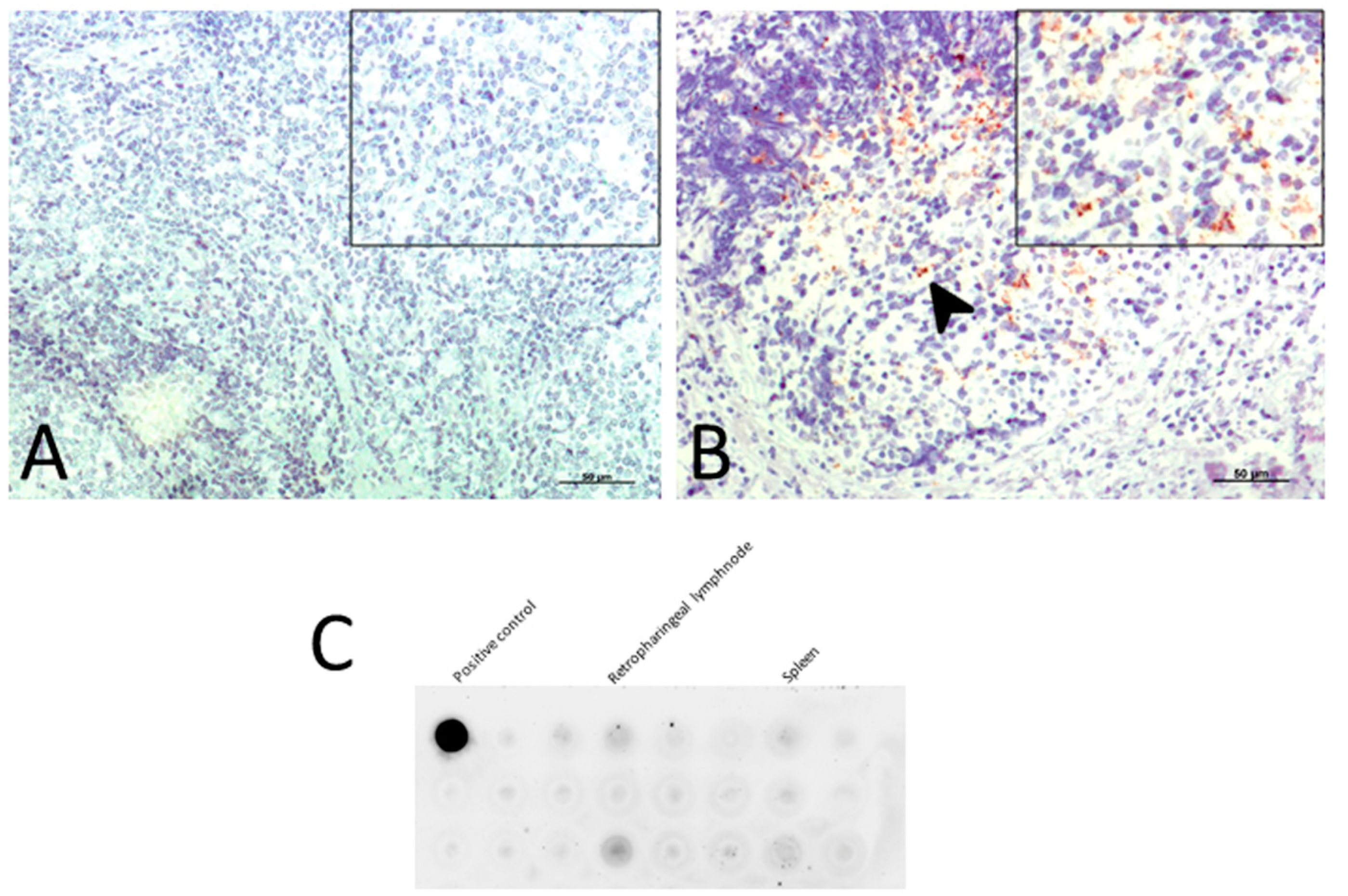
2.2.1. PrPsc Deposits
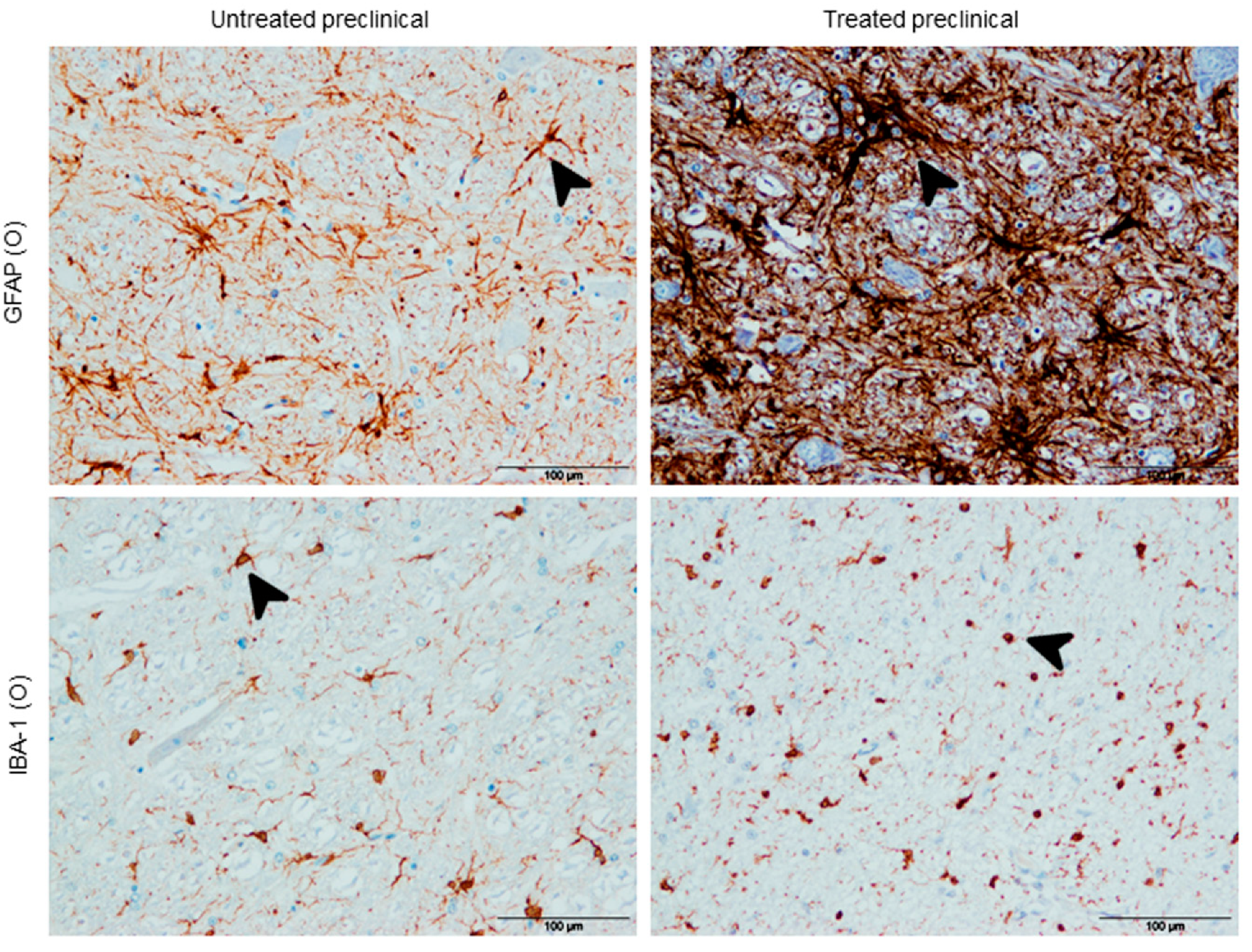
2.2.2. Astrogliois
2.2.3. Microgliosis
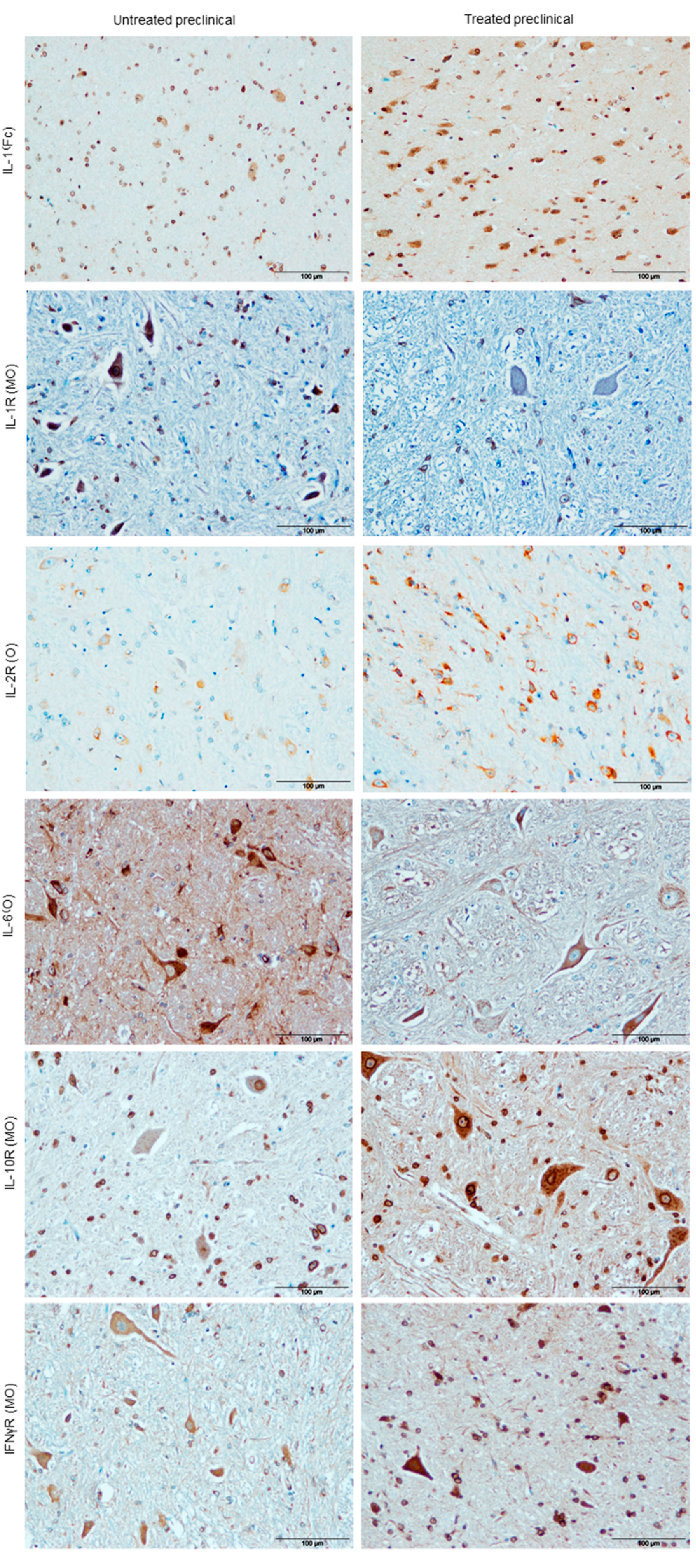
2.2.4. Cytokine Detection
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Methodology
4.3.1. Protein Misfolding Cyclic Amplification (PMCA)
4.3.2. Dot Blot (DB)
4.3.3. Histopathological (H-E) Studies
4.3.4. Immunohistochemical Techniques (IHC)
Pathological Prion Protein (PrPsc) Detection
Glial Fibrillary Acidic Protein (GFAP) Detection for Astrogliosis
Ionized Calcium-Binding Adaptor Molecule-1 (IBA-1) Detection for Microgliosis
IL-1α, IL-1R, IL-6 and IFNγR Detection
IL-2R, IL-10R and TNFαR Detection
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ligios, C.; Viglietti, A.; Carta, P.; Dexter, G.; Agrimi, U.; Simmons, M.M. Clinicopathological findings in sheep from Sardinia showing neurological signs of disease. Vet. Rec. 2004, 154, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Kummer, M.P.; Latz, E. Innate immune activation in neurodegenerative disease. Nat. Rev. Immunol. 2014, 14, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. How neuroinflammation contributes to neurodegeneration. Science 2016, 353, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Thangavel, R.; Natteru, P.A.; Selvakumar, G.P.; Saeed, D.; Zahoor, H.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Neuroinflammation Induces Neurodegeneration. J. Neurol. Neurosurg. Spine 2016, 1, 1003. [Google Scholar] [PubMed]
- Schwartz, M.; Deczkowska, A. Neurological Disease as a Failure of Brain-Immune Crosstalk: The Multiple Faces of Neuroinflammation. Trends Immunol. 2016, 37, 668–679. [Google Scholar] [CrossRef]
- Kordek, R.; Nerurkar, V.R.; Liberski, P.P.; Isaacson, S.; Yanagihara, R.; Gajdusek, D.C. Heightened expression of tumor necrosis factor alpha, interleukin 1 alpha, and glial fibrillary acidic protein in experimental Creutzfeldt-Jakob disease in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 9754–9758. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.E.; van Dam, A.M.; Man, A.H.W.K.; Berkenbosch, F.; Eikelenboom, P.; Fraser, H. Cytokines, prostaglandins and lipocortin-1 are present in the brains of scrapie-infected mice. Brain Res. 1994, 654, 200–206. [Google Scholar] [CrossRef]
- Williams, A.; Van Dam, A.M.; Ritchie, D.; Eikelenboom, P.; Fraser, H. Immunocytochemical appearance of cytokines, prostaglandin E2 and lipocortin-1 in the CNS during the incubation period of murine scrapie correlates with progressive PrP accumulations. Brain Res. 1997, 754, 171–180. [Google Scholar] [CrossRef]
- Gomez-Nicola, D.; Fransen, N.L.; Suzzi, S.; Perry, V.H. Regulation of microglial proliferation during chronic neurodegeneration. J. Neurosci. 2013, 33, 2481–2493. [Google Scholar] [CrossRef]
- Zhu, C.; Herrmann, U.S.; Falsig, J.; Abakumova, I.; Nuvolone, M.; Schwarz, P.; Frauenknecht, K.; Rushing, E.J.; Aguzzi, A. A neuroprotective role for microglia in prion diseases. J. Exp. Med. 2016, 213, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.A.; Race, B.; Williams, K.; Striebel, J.; Chesebro, B. Microglia Are Critical in Host Defense against Prion Disease. J. Virol. 2018, 92, e00549-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos-Carcavilla, A.; Calvo, J.H.; Gonzalez, C.; Moazami-Goudarzi, K.; Laurent, P.; Bertaud, M.; Hayes, H.; Beattie, A.E.; Serrano, C.; Lyahyai, J.; et al. IL-1 family members as candidate genes modulating scrapie susceptibility in sheep: Localization, partial characterization, and expression. Mamm. Genome 2007, 18, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Servida, F.; Ravasi, C.; Puricelli, M.; Formentin, E.A.; Dall’Ara, P.; Poli, G. Decrease in neuroinflammation after immunisation with synthetic prion peptides in an animal model of scrapie. Vet. Res. Commun. 2007, 31, 265–267. [Google Scholar] [CrossRef] [PubMed]
- Guijarro, I.M.; Garces, M.; Andres-Benito, P.; Marin, B.; Otero, A.; Barrio, T.; Carmona, M.; Ferrer, I.; Badiola, J.J.; Monzon, M. Assessment of Glial Activation Response in the Progress of Natural Scrapie after Chronic Dexamethasone Treatment. Int. J. Mol. Sci. 2020, 21, 3231. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.L.; Eddleston, M.; Kemper, P.; Oldstone, M.B.; Hobbs, M.V. Activation of cerebral cytokine gene expression and its correlation with onset of reactive astrocyte and acute-phase response gene expression in scrapie. J. Virol. 1994, 68, 2383–2387. [Google Scholar] [CrossRef] [Green Version]
- Veerhuis, R.; Hoozemans, J.J.; Janssen, I.; Boshuizen, R.S.; Langeveld, J.P.; Eikelenboom, P. Adult human microglia secrete cytokines when exposed to neurotoxic prion protein peptide: No intermediary role for prostaglandin E2. Brain Res. 2002, 925, 195–203. [Google Scholar] [CrossRef]
- Brown, A.R.; Webb, J.; Rebus, S.; Walker, R.; Williams, A.; Fazakerley, J.K. Inducible cytokine gene expression in the brain in the ME7/CV mouse model of scrapie is highly restricted, is at a strikingly low level relative to the degree of gliosis and occurs only late in disease. J. Gen. Virol. 2003, 84, 2605–2611. [Google Scholar] [CrossRef]
- Hernandez, R.S.; Sarasa, R.; Toledano, A.; Badiola, J.J.; Monzon, M. Morphological approach to assess the involvement of astrocytes in prion propagation. Cell Tissue Res. 2014, 358, 57–63. [Google Scholar] [CrossRef]
- Monzon, M.; Hernandez, R.S.; Garces, M.; Sarasa, R.; Badiola, J.J. Glial alterations in human prion diseases: A correlative study of astroglia, reactive microglia, protein deposition, and neuropathological lesions. Medicine (Baltim.) 2018, 97, e0320. [Google Scholar] [CrossRef]
- Garces, M.; Guijarro, M.I.; Vargas, A.; Badiola, J.J.; Monzon, M. Neuroglial patterns are shared by cerebella from prion and prion-like disorder affected patients. Mech. Ageing Dev. 2019, 184, 111176. [Google Scholar] [CrossRef]
- Perry, V.H.; Cunningham, C.; Boche, D. Atypical inflammation in the central nervous system in prion disease. Curr. Opin. Neurol. 2002, 15, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.A.; Striebel, J.F.; Race, B.; Phillips, K.; Chesebro, B. Prion infection of mouse brain reveals multiple new upregulated genes involved in neuroinflammation or signal transduction. J. Virol. 2015, 89, 2388–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkhratsky, A.; Rodrigues, J.J.; Pivoriunas, A.; Zorec, R.; Semyanov, A. Astroglial atrophy in Alzheimer’s disease. Pflug. Arch. 2019, 471, 1247–1261. [Google Scholar] [CrossRef] [PubMed]
- Llorens, F.; Lopez-Gonzalez, I.; Thune, K.; Carmona, M.; Zafar, S.; Andreoletti, O.; Zerr, I.; Ferrer, I. Subtype and regional-specific neuroinflammation in sporadic creutzfeldt-jakob disease. Front. Aging Neurosci. 2014, 6, 198. [Google Scholar] [CrossRef] [PubMed]
- Garwood, C.J.; Pooler, A.M.; Atherton, J.; Hanger, D.P.; Noble, W. Astrocytes are important mediators of Abeta-induced neurotoxicity and tau phosphorylation in primary culture. Cell Death Dis. 2011, 2, e167. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.W.; Gerlach, M.; Riederer, P.; Klein, M.A. Oxidative stress in the brain at early preclinical stages of mouse scrapie. Exp. Neurol. 2006, 201, 90–98. [Google Scholar] [CrossRef]
- Kretlow, A.; Wang, Q.; Beekes, M.; Naumann, D.; Miller, L.M. Changes in protein structure and distribution observed at pre-clinical stages of scrapie pathogenesis. Biochim. Biophys. Acta. 2008, 1782, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Manca, M.; Foutz, A.; Camacho, M.V.; Raymond, G.J.; Race, B.; Orru, C.D.; Yuan, J.; Shen, P.; Li, B.; et al. Early preclinical detection of prions in the skin of prion-infected animals. Nat. Commun. 2019, 10, 247. [Google Scholar] [CrossRef]
- Halliez, S.; Jaumain, E.; Huor, A.; Douet, J.Y.; Lugan, S.; Cassard, H.; Lacroux, C.; Beringue, V.; Andreoletti, O.; Vilette, D. White blood cell-based detection of asymptomatic scrapie infection by ex vivo assays. PLoS ONE 2014, 9, e104287. [Google Scholar] [CrossRef]
- Fernandez, A.P.; Serrano, J.; Rodrigo, J.; Monleon, E.; Monzon, M.; Vargas, A.; Badiola, J.J.; Martinez-Murillo, R.; Martinez, A. Changes in the expression pattern of the nitrergic system of ovine cerebellum affected by scrapie. J. Neuropathol. Exp. Neurol. 2007, 66, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Gough, K.C.; Baker, C.A.; Rees, H.C.; Terry, L.A.; Spiropoulos, J.; Thorne, L.; Maddison, B.C. The oral secretion of infectious scrapie prions occurs in preclinical sheep with a range of PRNP genotypes. J. Virol. 2012, 86, 566–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetterberg, H.; Bozzetta, E.; Favole, A.; Corona, C.; Cavarretta, M.C.; Ingravalle, F.; Blennow, K.; Pocchiari, M.; Meloni, D. Neurofilaments in blood is a new promising preclinical biomarker for the screening of natural scrapie in sheep. PLoS ONE 2019, 14, e0226697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyahyai, J.; Bolea, R.; Serrano, C.; Monleon, E.; Moreno, C.; Osta, R.; Zaragoza, P.; Badiola, J.J.; Martin-Burriel, I. Correlation between Bax overexpression and prion deposition in medulla oblongata from natural scrapie without evidence of apoptosis. Acta Neuropathol. 2006, 112, 451–460. [Google Scholar] [CrossRef]
- Imbimbo, B.P. An update on the efficacy of non-steroidal anti-inflammatory drugs in Alzheimer’s disease. Expert Opin. Investig. Drugs 2009, 18, 1147–1168. [Google Scholar] [CrossRef] [PubMed]
- Zeug, A.; Muller, F.E.; Anders, S.; Herde, M.K.; Minge, D.; Ponimaskin, E.; Henneberger, C. Control of astrocyte morphology by Rho GTPases. Brain Res. Bull. 2018, 136, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.H.; O’Connor, V. The role of microglia in synaptic stripping and synaptic degeneration: A revised perspective. ASN Neuro. 2010, 2, e00047. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.J.; Xue, Q.S.; Tischer, J.; Bechmann, I. Microglial pathology. Acta Neuropathol. Commun. 2014, 2, 142. [Google Scholar] [CrossRef]
- Kim, S.U.; de Vellis, J. Microglia in health and disease. J. Neurosci. Res. 2005, 81, 302–313. [Google Scholar] [CrossRef]
- Burwinkel, M.; Riemer, C.; Schwarz, A.; Schultz, J.; Neidhold, S.; Bamme, T.; Baier, M. Role of cytokines and chemokines in prion infections of the central nervous system. Int. J. Dev. Neurosci. 2004, 22, 497–505. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L.; et al. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Griffin, W.S.; Mrak, R.E. Interleukin-1 in the genesis and progression of and risk for development of neuronal degeneration in Alzheimer’s disease. J. Leukoc. Biol. 2002, 72, 233–238. [Google Scholar] [PubMed]
- Sastre, M.; Klockgether, T.; Heneka, M.T. Contribution of inflammatory processes to Alzheimer’s disease: Molecular mechanisms. Int. J. Dev. Neurosci. 2006, 24, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Schwarz, A.; Neidhold, S.; Burwinkel, M.; Riemer, C.; Simon, D.; Kopf, M.; Otto, M.; Baier, M. Role of interleukin-1 in prion disease-associated astrocyte activation. Am. J. Pathol. 2004, 165, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Bruttger, J.; Karram, K.; Wortge, S.; Regen, T.; Marini, F.; Hoppmann, N.; Klein, M.; Blank, T.; Yona, S.; Wolf, Y.; et al. Genetic Cell Ablation Reveals Clusters of Local Self-Renewing Microglia in the Mammalian Central Nervous System. Immunity 2015, 43, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Sarder, M.; Abe, K.; Saito, H.; Nishiyama, N. Comparative effect of IL-2 and IL-6 on morphology of cultured hippocampal neurons from fetal rat brain. Brain Res. 1996, 715, 9–16. [Google Scholar] [CrossRef]
- Dansokho, C.; Ait Ahmed, D.; Aid, S.; Toly-Ndour, C.; Chaigneau, T.; Calle, V.; Cagnard, N.; Holzenberger, M.; Piaggio, E.; Aucouturier, P.; et al. Regulatory T cells delay disease progression in Alzheimer-like pathology. Brain 2016, 139, 1237–1251. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Yoshizaki, K.; Kishimoto, T.; Ito, H. IL-6 is required for the development of Th1 cell-mediated murine colitis. J. Immnol. 2000, 164, 4878–4882. [Google Scholar] [CrossRef] [Green Version]
- Hafiz, F.B.; Brown, D.R. A model for the mechanism of astrogliosis in prion disease. Mol. Cell. Neurosci. 2000, 16, 221–232. [Google Scholar] [CrossRef]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8, S3. [Google Scholar] [CrossRef] [Green Version]
- Strauss, S.; Bauer, J.; Ganter, U.; Jonas, U.; Berger, M.; Volk, B. Detection of interleukin-6 and alpha 2-macroglobulin immunoreactivity in cortex and hippocampus of Alzheimer’s disease patients. Lab. Investig. 1992, 66, 223–230. [Google Scholar]
- Wood, J.A.; Wood, P.L.; Ryan, R.; Graff-Radford, N.R.; Pilapil, C.; Robitaille, Y.; Quirion, R. Cytokine indices in Alzheimer’s temporal cortex: No changes in mature IL-1 beta or IL-1RA but increases in the associated acute phase proteins IL-6, alpha 2-macroglobulin and C-reactive protein. Brain Res. 1993, 629, 245–252. [Google Scholar] [CrossRef]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Thackray, A.M.; McKenzie, A.N.; Klein, M.A.; Lauder, A.; Bujdoso, R. Accelerated prion disease in the absence of interleukin-10. J. Virol. 2004, 78, 13697–13707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamguney, G.; Giles, K.; Glidden, D.V.; Lessard, P.; Wille, H.; Tremblay, P.; Groth, D.F.; Yehiely, F.; Korth, C.; Moore, R.C.; et al. Genes contributing to prion pathogenesis. J. Gen. Virol. 2008, 89, 1777–1788. [Google Scholar] [CrossRef]
- Corbin, J.G.; Kelly, D.; Rath, E.M.; Baerwald, K.D.; Suzuki, K.; Popko, B. Targeted CNS expression of interferon-gamma in transgenic mice leads to hypomyelination, reactive gliosis, and abnormal cerebellar development. Mol. Cell. Neurosci. 1996, 7, 354–370. [Google Scholar] [CrossRef]
- Mabbott, N.A.; Williams, A.; Farquhar, C.F.; Pasparakis, M.; Kollias, G.; Bruce, M.E. Tumor necrosis factor alpha-deficient, but not interleukin-6-deficient, mice resist peripheral infection with scrapie. J. Virol. 2000, 74, 3338–3344. [Google Scholar] [CrossRef] [Green Version]
- Bowers, S.L.; Bilbo, S.D.; Dhabhar, F.S.; Nelson, R.J. Stressor-specific alterations in corticosterone and immune responses in mice. Brain Behav. Immun. 2008, 22, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Sorrells, S.F.; Caso, J.R.; Munhoz, C.D.; Sapolsky, R.M. The stressed CNS: When glucocorticoids aggravate inflammation. Neuron 2009, 64, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Zhang, Y.; Wu, W.; Yin, Y.; Huang, D.; Wang, Y.; Li, W.; Li, W. Chronic glucocorticoids exposure enhances neurodegeneration in the frontal cortex and hippocampus via NLRP-1 inflammasome activation in male mice. Brain Behav. Immun. 2016, 52, 58–70. [Google Scholar] [CrossRef]
- Tribouillard-Tanvier, D.; Striebel, J.F.; Peterson, K.E.; Chesebro, B. Analysis of protein levels of 24 cytokines in scrapie agent-infected brain and glial cell cultures from mice differing in prion protein expression levels. J. Virol. 2009, 83, 11244–11253. [Google Scholar] [CrossRef] [Green Version]
- Tribouillard-Tanvier, D.; Race, B.; Striebel, J.F.; Carroll, J.A.; Phillips, K.; Chesebro, B. Early cytokine elevation, PrPres deposition, and gliosis in mouse scrapie: No effect on disease by deletion of cytokine genes IL-12p40 and IL-12p35. J. Virol. 2012, 86, 10377–10383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacroux, C.; Comoy, E.; Moudjou, M.; Perret-Liaudet, A.; Lugan, S.; Litaise, C.; Simmons, H.; Jas-Duval, C.; Lantier, I.; Beringue, V.; et al. Preclinical detection of variant CJD and BSE prions in blood. PLoS Pathog. 2014, 10, e1004202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douet, J.Y.; Lacroux, C.; Aron, N.; Head, M.W.; Lugan, S.; Tillier, C.; Huor, A.; Cassard, H.; Arnold, M.; Beringue, V.; et al. Distribution and Quantitative Estimates of Variant Creutzfeldt-Jakob Disease Prions in Tissues of Clinical and Asymptomatic Patients. Emerg. Infect. Dis. 2017, 23, 946–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Trevejo, J.L.; Gomez-Villamandos, J.C.; Pedrera, M.; Blanco, A.; Bautista, M.J.; Sanchez-Cordon, P.J. Immunohistochemical study of macrophage and cytokine dynamics in the gut of scrapie-infected mice. Histol. Histopathol. 2010, 25, 1025–1038. [Google Scholar]
- Monleon, E.; Monzon, M.; Hortells, P.; Vargas, A.; Acin, C.; Badiola, J.J. Detection of PrPsc on lymphoid tissues from naturally affected scrapie animals: Comparison of three visualization systems. J. Histochem. Cytochem. 2004, 52, 145–151. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker Assessed | DEX Effect | Brain Area | Statistical Significance |
|---|---|---|---|
| Vacuolation | Decrease | O | * p ≤ 0.05 |
| Prion protein deposition | Decrease | O MO | ** p ≤ 0.01 ** p ≤ 0.01 |
| GFAP | Increase | O MO | ** p ≤ 0.01 ** p ≤ 0.01 |
| IBA-1 | Increase | Fc Cb O MO | * p ≤ 0.05 # p = 0.08 * p ≤ 0.05 * p ≤ 0.05 |
| IL-1α | Increase | Fc | ** p ≤ 0.01 |
| IL-1R | Decrease | MO | * p ≤ 0.05 |
| IL-2R | Increase | O | * p ≤ 0.05 |
| IL-6 | Decrease | O MO | *** p ≤ 0.001 *** p ≤ 0.001 |
| IL-10R | Increase | Fc Cb O MO | ** p ≤ 0.01 ** p ≤ 0.01 # p = 0.07 * p ≤ 0.05 |
| TNFαR | No changes | p > 0.05 | |
| IFNγR | Increase | Cb O MO | ** p ≤ 0.01 ** p ≤ 0.01 ** p ≤ 0.01 |
| Sheep No. | PRNP Genotype | Age (Years) | Group | Treatment and Duration |
|---|---|---|---|---|
| 1 | ARQ/ARQ | 3 | Preclinical | Untreated |
| 2 | ARQ/ARQ | 3 | Preclinical | Untreated |
| 3 | ARQ/ARQ | 6 | Preclinical | Untreated |
| 4 | ARQ/ARQ | 3 | Preclinical | Untreated |
| 5 | ARK/ARQ | 4 | Preclinical | Untreated |
| 6 | ARQ/ARQ | 8 | Preclinical | Treated (7 months) |
| 7 | ARQ/ARQ | 3 | Preclinical | Treated (11 months) |
| 8 | ARQ/ARQ | 7 | Preclinical | Treated (12 months) |
| 9 | ARQ/ARQ | 8 | Preclinical | Treated (13 months) |
| 10 | ARQ/ARQ | 5 | Preclinical | Treated (13 months) |
| 11 | ARQ/ARQ | 5 | Preclinical | Treated (14 months) |
| 12 | ARQ/ARQ | 6 | Preclinical | Treated (17 months) |
| Antibody | Antigen | Type | Dilution | Retrieval Method | Source |
|---|---|---|---|---|---|
| L42 | Prion protein | Monoclonal | 1:500 | Formic acid 15 min Proteinase K 15 min Heat treatment 20 min Peroxidase blocking | DAKO |
| Anti- GFAP | GFAP | Polyclonal | 1:500 | Peroxidase blocking | DAKO |
| Anti-IBA-1 | IBA-1 | Polyclonal | 1:1.000 | Heat treatment 20 min Peroxidase blocking | WAKO |
| IL-1 alpha | IL-1 | Polyclonal | 1:100 | Autoclave 121 °C (citrate buffer 10%) | ThermoFisher |
| Anti-IL-1RN | IL-1R | Polyclonal | 1:100 | Autoclave 121 °C (citrate buffer 10%) | Sigma |
| IL-2R.1 | IL-2R | Monoclonal | 1:1.000 | PTLink 96 °C | ThermoFisher |
| 8H12 | IL-6 | Monoclonal | 1:20 | Autoclave 121 °C (citrate buffer 10%) | ThermoFisher |
| OTI1D10 | IL-10R | Monoclonal | 1:250 | PTLink 96 °C | ThermoFisher |
| Ber-H2 | TNFαR | Monoclonal | Ready to use | PTLink 96 °C | DAKO |
| IFNGR1 | IFNγR | Polyclonal | 1:200 | Autoclave 121 °C (citrate buffer 10%) | ThermoFisher |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guijarro, I.M.; Garcés, M.; Marín, B.; Otero, A.; Barrio, T.; Badiola, J.J.; Monzón, M. Neuroimmune Response in Natural Preclinical Scrapie after Dexamethasone Treatment. Int. J. Mol. Sci. 2020, 21, 5779. https://doi.org/10.3390/ijms21165779
Guijarro IM, Garcés M, Marín B, Otero A, Barrio T, Badiola JJ, Monzón M. Neuroimmune Response in Natural Preclinical Scrapie after Dexamethasone Treatment. International Journal of Molecular Sciences. 2020; 21(16):5779. https://doi.org/10.3390/ijms21165779
Chicago/Turabian StyleGuijarro, Isabel M., Moisés Garcés, Belén Marín, Alicia Otero, Tomás Barrio, Juan J. Badiola, and Marta Monzón. 2020. "Neuroimmune Response in Natural Preclinical Scrapie after Dexamethasone Treatment" International Journal of Molecular Sciences 21, no. 16: 5779. https://doi.org/10.3390/ijms21165779