Perinatal Exposure to Triclosan Results in Abnormal Brain Development and Behavior in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. TCS Impairs to Growth and Development of Primary Cortical Neuron
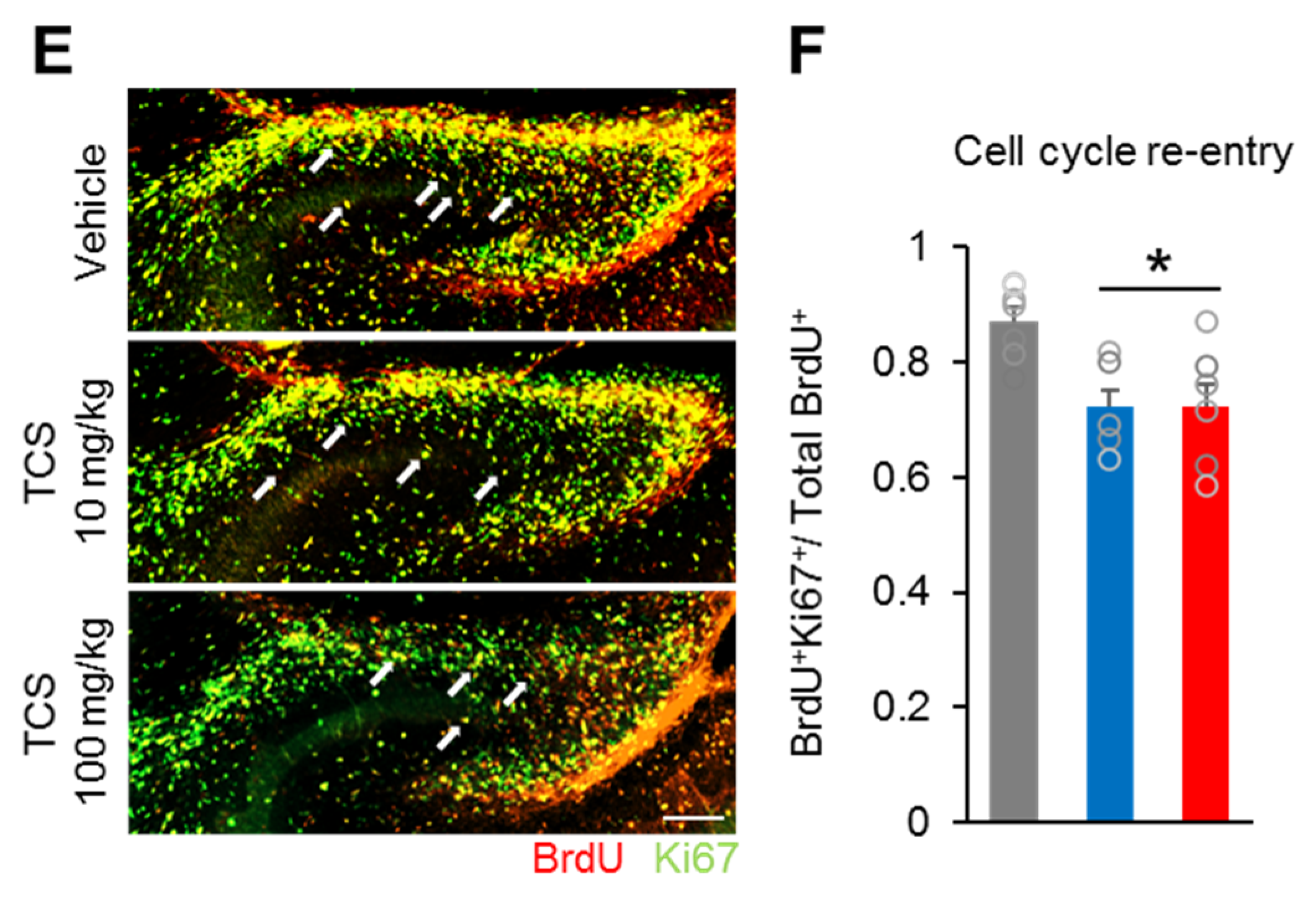
2.2. TCS Impairs Neuronal Progenitor Cell Proliferation and Cell Death
2.3. TCS-Exposure Result in Growth and Developmental Delay
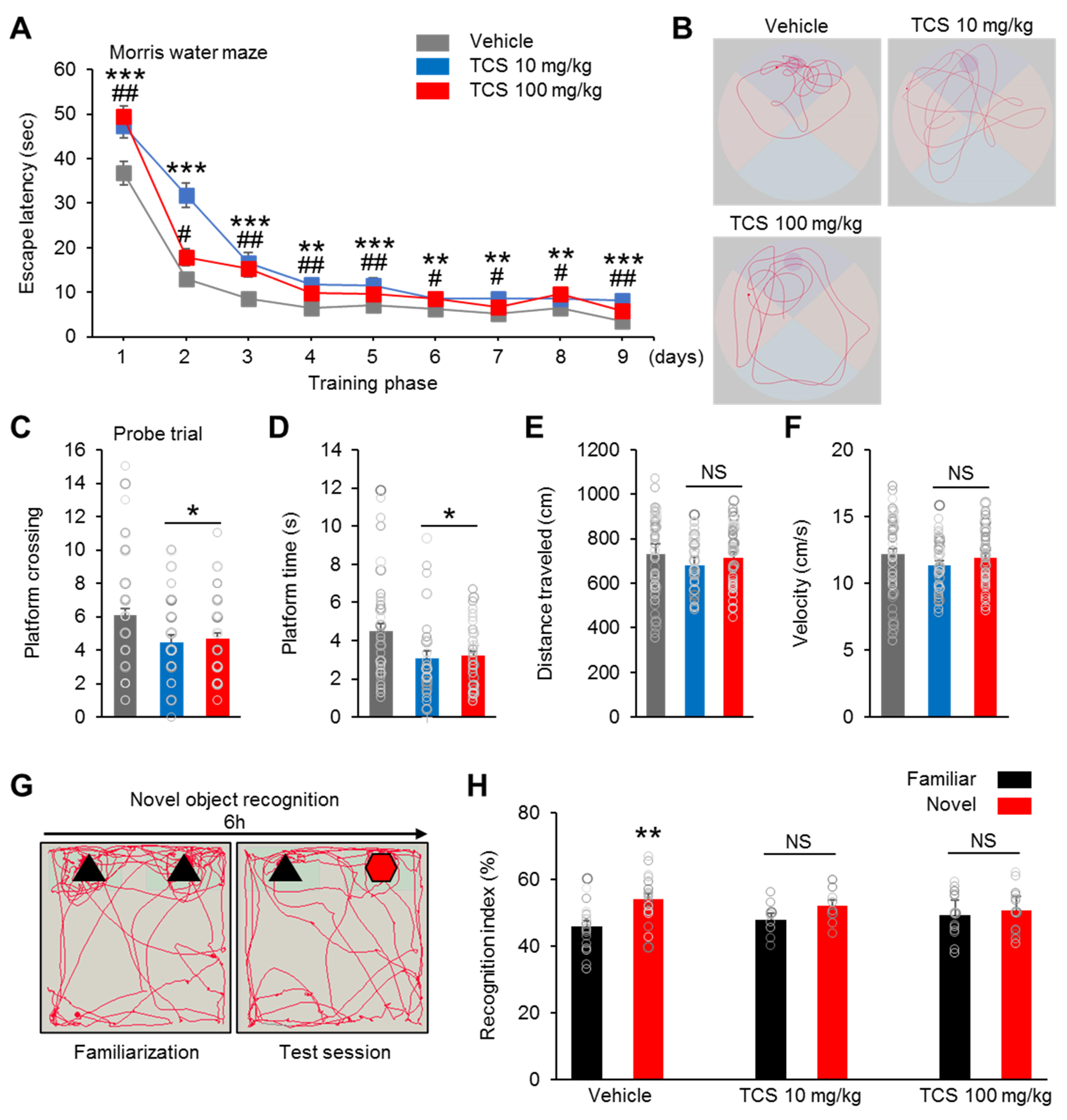
2.4. TCS-Exposure Induce Cognition Dysfunction in Offspring Mice
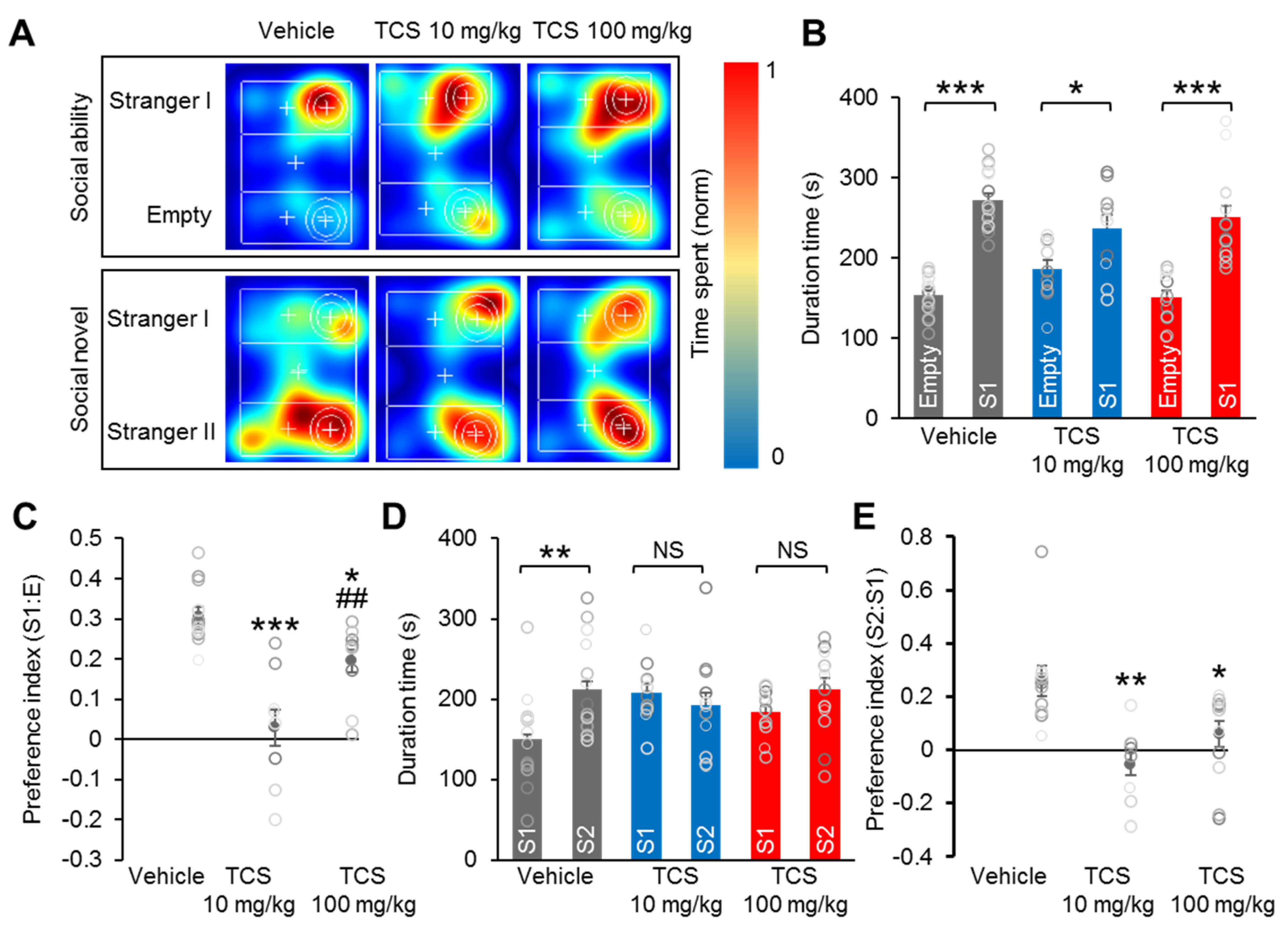
2.5. TCS Induces Social Deficiency in Mice
2.6. TCS-Exposure Induces Anxiety-Like Behavior, but does not Affect Depression-Like Behavior in Mice
2.7. TCS-Exposure Reduces Nesting-Behavior in Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Treatments
4.2. Primary Cortical Neuron Culture
Neuronal Morphology
4.3. Immunofluorescence
4.3.1. Staining without BrdU
4.3.2. BrdU Staining
4.4. RNA Extraction and Quantitative Real-Time PCR
4.5. Western-Blot Analysis
4.6. Behavioral Analysis
4.6.1. Experimental design.
4.6.2. Open Field Test
4.6.3. Morris Water Maze
4.6.4. Novel Object Recognition
4.6.5. Tail Suspension Test
4.6.6. Forced Swimming Test
4.6.7. Three-Chamber Social Test
4.6.8. Nest-Building Test
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ginsberg, G.L.; Balk, S.J. Consumer products as sources of chemical exposures to children: Case study of triclosan. Curr. Opin. Pediatr. 2016, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Gee, R.; Charles, A.; Taylor, N.; Darbre, P. Oestrogenic and androgenic activity of triclosan in breast cancer cells. J. Appl. Toxicol. Int. J. 2008, 28, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.-M.; An, B.-S.; Choi, K.-C.; Jeung, E.-B. Potential estrogenic activity of triclosan in the uterus of immature rats and rat pituitary GH3 cells. Toxicol. Lett. 2012, 208, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Geens, T.; Neels, H.; Covaci, A. Distribution of bisphenol-A, triclosan and n-nonylphenol in human adipose tissue, liver and brain. Chemosphere 2012, 87, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Ye, X.; Wong, L.-Y.; Reidy, J.A.; Needham, L.L. Urinary concentrations of triclosan in the US population: 2003–2004. Environ. Health Perspect. 2007, 116, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.-W.; Wu, Q.; Green, B.; Nolen, G.; Shi, L.; LoSurdo, J.; Deng, H.; Bauer, S.; Fang, J.-L.; Ning, B. Cytotoxicity and inhibitory effects of low-concentration triclosan on adipogenic differentiation of human mesenchymal stem cells. Toxicol. Appl. Pharmacol. 2012, 262, 117–123. [Google Scholar] [CrossRef]
- Lankester, J.; Patel, C.; Cullen, M.R.; Ley, C.; Parsonnet, J. Urinary triclosan is associated with elevated body mass index in NHANES. PLoS ONE 2013, 8, e80057. [Google Scholar] [CrossRef]
- Wang, C.; Chen, L.; Zhao, S.; Hu, Y.; Zhou, Y.; Gao, Y.; Wang, W.; Zhang, J.; Tian, Y. Impacts of prenatal triclosan exposure on fetal reproductive hormones and its potential mechanism. Environ. Int. 2018, 111, 279–286. [Google Scholar] [CrossRef]
- Etzel, T.M.; Calafat, A.M.; Ye, X.; Chen, A.; Lanphear, B.P.; Savitz, D.A.; Yolton, K.; Braun, J.M. Urinary triclosan concentrations during pregnancy and birth outcomes. Environ. Res. 2017, 156, 505–511. [Google Scholar] [CrossRef]
- Rodríguez, P.E.; Sanchez, M.S. Maternal exposure to triclosan impairs thyroid homeostasis and female pubertal development in Wistar rat offspring. J. Toxicol. Environ. Healthpart A 2010, 73, 1678–1688. [Google Scholar]
- Cao, X.; Hua, X.; Wang, X.; Chen, L. Exposure of pregnant mice to triclosan impairs placental development and nutrient transport. Sci. Rep. 2017, 7, 44803. [Google Scholar] [CrossRef] [PubMed]
- Ruszkiewicz, J.A.; Li, S.; Rodriguez, M.B.; Aschner, M. Is triclosan a neurotoxic agent? J. Toxicol. Environ. Healthpart B 2017, 20, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Arias-Cavieres, A.; More, J.; Vicente, J.M.; Adasme, T.; Hidalgo, J.; Valdés, J.L.; Humeres, A.; Valdés-Undurraga, I.; Sanchez, G.; Hidalgo, C. Triclosan Impairs Hippocampal Synaptic Plasticity and Spatial Memory in Male Rats. Front. Mol. Neurosci. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabari, S.A.; Esfahani, M.L.; Hosseini, S.M.; Rahimi, A. Neurobehavioral toxicity of triclosan in mice. Food Chem. Toxicol. 2019, 130, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Park, B.K.; Gonzales, E.L.T.; Yang, S.M.; Bang, M.; Choi, C.S.; Shin, C.Y. Effects of triclosan on neural stem cell viability and survival. Biomol. Ther. (Seoul) 2016, 24, 99. [Google Scholar] [CrossRef] [Green Version]
- Szychowski, K.A.; Wnuk, A.; Rzemieniec, J.; Kajta, M.; Leszczyńska, T.; Wójtowicz, A.K. Triclosan-evoked neurotoxicity involves NMDAR subunits with the specific role of GluN2A in caspase-3-dependent apoptosis. Mol. Neurobiol. 2019, 56, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Muth-Köhne, E.; Wichmann, A.; Delov, V.; Fenske, M. The classification of motor neuron defects in the zebrafish embryo toxicity test (ZFET) as an animal alternative approach to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2012, 34, 413–424. [Google Scholar]
- Rabaglino, M.B.; Chang, E.I.; Richards, E.M.; James, M.O.; Keller-Wood, M.; Wood, C.E. Genomic effect of triclosan on the fetal hypothalamus: Evidence for altered neuropeptide regulation. Endocrinology 2016, 157, 2686–2697. [Google Scholar] [CrossRef] [Green Version]
- Shonkoff, J.P.; Boyce, W.T.; McEwen, B.S. Neuroscience, molecular biology, and the childhood roots of health disparities: Building a new framework for health promotion and disease prevention. JAMA 2009, 301, 2252–2259. [Google Scholar] [CrossRef]
- Dezonne, R.; Lima, F.; Trentin, A.; Gomes, F. Thyroid hormone and astroglia: Endocrine control of the neural environment. J. Neuroendocrinol. 2015, 27, 435–445. [Google Scholar] [CrossRef]
- Henrichs, J.; Bongers-Schokking, J.J.; Schenk, J.J.; Ghassabian, A.; Schmidt, H.G.; Visser, T.J.; Hooijkaas, H.; de Muinck Keizer-Schrama, S.M.; Hofman, A.; Jaddoe, V.V. Maternal thyroid function during early pregnancy and cognitive functioning in early childhood: The generation R study. J. Clin. Endocrinol. Metab. 2010, 95, 4227–4234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dann, A.B.; Hontela, A. Triclosan: Environmental exposure, toxicity and mechanisms of action. J. Appl. Toxicol. 2011, 31, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, O.; Oishi, S. Disposition of orally administered 2, 2-Bis (4-hydroxyphenyl) propane (Bisphenol A) in pregnant rats and the placental transfer to fetuses. Environ. Health Perspect. 2000, 108, 931–935. [Google Scholar] [CrossRef]
- Tran, D.; Jung, E.-M.; Ahn, C.; Lee, J.-H.; Yoo, Y.-M.; Jeung, E.-B. Effects of bisphenol A and 4-tert-octylphenol on embryo implantation failure in mouse. Int. J. Environ. Res. Public Health 2018, 15, 1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schug, T.T.; Blawas, A.M.; Gray, K.; Heindel, J.J.; Lawler, C.P. Elucidating the links between endocrine disruptors and neurodevelopment. Endocrinology 2015, 156, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Eom, S.; Kim, H.-J.; Lee, J.J.; Choi, G.; Choi, S.; Kim, S.; Kim, S.Y.; Cho, G.; Kim, Y.D. Association between maternal exposure to major phthalates, heavy metals, and persistent organic pollutants, and the neurodevelopmental performances of their children at 1 to 2 years of age-CHECK cohort study. Sci. Total Environ. 2018, 624, 377–384. [Google Scholar] [CrossRef]
- Allmyr, M.; Adolfsson-Erici, M.; McLachlan, M.S.; Sandborgh-Englund, G. Triclosan in plasma and milk from Swedish nursing mothers and their exposure via personal care products. Sci. Total Environ. 2006, 372, 87–93. [Google Scholar] [CrossRef]
- Allmyr, M.; Harden, F.; Toms, L.-M.L.; Mueller, J.F.; McLachlan, M.S.; Adolfsson-Erici, M.; Sandborgh-Englund, G. The influence of age and gender on triclosan concentrations in Australian human blood serum. Sci. Total Environ. 2008, 393, 162–167. [Google Scholar] [CrossRef]
- Pycke, B.F.; Geer, L.A.; Dalloul, M.; Abulafia, O.; Jenck, A.M.; Halden, R.U. Human fetal exposure to triclosan and triclocarban in an urban population from Brooklyn, New York. Environ. Sci. Technol. 2014, 48, 8831–8838. [Google Scholar] [CrossRef]
- Farkas, L.M.; Huttner, W.B. The cell biology of neural stem and progenitor cells and its significance for their proliferation versus differentiation during mammalian brain development. Curr. Opin. Cell Biol. 2008, 20, 707–715. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Gotoh, H.; Ono, K.; Nomura, T. Differential potentials of neural progenitors for the generation of neurons and non-neuronal cells in the developing amniote brain. Sci. Rep. 2019, 9, 4514. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.A.; Firestein, B.L. The dendritic tree and brain disorders. Mol. Cell. Neurosci. 2012, 50, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Beyer, C. Estrogen and the developing mammalian brain. Anat. Embryol. (Berl.) 1999, 199, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Ghassabian, A.; El Marroun, H.; Peeters, R.P.; Jaddoe, V.W.; Hofman, A.; Verhulst, F.C.; Tiemeier, H.; White, T. Downstream effects of maternal hypothyroxinemia in early pregnancy: Nonverbal IQ and brain morphology in school-age children. J. Clin. Endocrinol. Metab. 2014, 99, 2383–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Escobar, G.M.; Obregón, M.J.; Del Rey, F.E. Role of thyroid hormone during early brain development. Eur. J. Endocrinol. 2004, 151, U25–U37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putcha, G.V.; Harris, C.A.; Moulder, K.L.; Easton, R.M.; Thompson, C.B.; Johnson, E.M. Intrinsic and extrinsic pathway signaling during neuronal apoptosis: Lessons from the analysis of mutant mice. J. Cell Biol. 2002, 157, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Anilkumar, U.; Prehn, J.H. Anti-apoptotic BCL-2 family proteins in acute neural injury. Front. Cell. Neurosci. 2014, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochman, A.; Sternin, H.; Gorodin, S.; Korsmeyer, S.; Ziv, I.; Melamed, E.; Offen, D. Enhanced oxidative stress and altered antioxidants in brains of Bcl-2-deficient mice. J. Neurochem. 1998, 71, 741–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, Y.; Rocchi, E.; Lazaro, J.; Clos, J. Thyroid hormone promotes BCL-2 expression and prevents apoptosis of early differentiating cerebellar granule neurons. Int. J. Dev. Neurosci. 1995, 13, 871–885. [Google Scholar] [CrossRef]
- Singh, R.; Upadhyay, G.; Kumar, S.; Kapoor, A.; Kumar, A.; Tiwari, M.; Godbole, M. Hypothyroidism alters the expression of Bcl-2 family genes to induce enhanced apoptosis in the developing cerebellum. J. Endocrinol. 2003, 176, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Louis, G.W.; Hallinger, D.R.; Braxton, M.J.; Kamel, A.; Stoker, T.E. Effects of chronic exposure to triclosan on reproductive and thyroid endpoints in the adult Wistar female rat. J. Toxicol. Environ. Healthpart A 2017, 80, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Beland, F.A.; Fang, J.-L. Effect of triclosan, triclocarban, 2, 2′, 4, 4′-tetrabromodiphenyl ether, and bisphenol A on the iodide uptake, thyroid peroxidase activity, and expression of genes involved in thyroid hormone synthesis. Toxicol. Vitr. 2016, 32, 310–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiamolera, M.I.; Wondisford, F.E. Thyrotropin-releasing hormone and the thyroid hormone feedback mechanism. Endocrinology 2009, 150, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, J.; Suzuki, A.; Kimura, T.; Tateyama, M.; Tanaka, T.; Yazawa, T.; Arimasu, Y.; Chen, I.-S.; Aoyama, K.; Kubo, Y. Congenital goitrous hypothyroidism is caused by dysfunction of the iodide transporter SLC26A7. Commun. Biol. 2019, 2, 270. [Google Scholar] [CrossRef] [Green Version]
- Dezonne, R.S.; Stipursky, J.; Gomes, F.C.A. Effect of thyroid hormone depletion on cultured murine cerebral cortex astrocytes. Neurosci. Lett. 2009, 467, 58–62. [Google Scholar] [CrossRef]
- Mayerl, S.; Müller, J.; Bauer, R.; Richert, S.; Kassmann, C.M.; Darras, V.M.; Buder, K.; Boelen, A.; Visser, T.J.; Heuer, H. Transporters MCT8 and OATP1C1 maintain murine brain thyroid hormone homeostasis. J. Clin. Investig. 2014, 124, 1987–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Nascimento, C.; Salgueiro, R.B.; Pantaleão, T.; da Costa, V.M.C.; Nunes, M.T. Maternal exposure to iodine excess throughout pregnancy and lactation induces hypothyroidism in adult male rat offspring. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Thompson, C.C.; Potter, G.B. Thyroid hormone action in neural development. Cereb. Cortex 2000, 10, 939–945. [Google Scholar] [CrossRef]
- Ganguli, M.; Burmeister, L.A.; Seaberg, E.C.; Belle, S.; DeKosky, S.T. Association between dementia and elevated TSH: A community-based study. Biol. Psychiatry 1996, 40, 714–725. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Feng, X.; Chang, F.; Chen, M.; Xia, Y.; Chen, L. Triclosan causes spontaneous abortion accompanied by decline of estrogen sulfotransferase activity in humans and mice. Sci. Rep. 2015, 5, 18252. [Google Scholar] [CrossRef] [Green Version]
- Jung, E.M.; Moffat, J.J.; Liu, J.; Dravid, S.M.; Gurumurthy, C.B.; Kim, W.Y. Arid1b haploinsufficiency disrupts cortical interneuron development and mouse behavior. Nat. Neurosci. 2017, 20, 1694–1707. [Google Scholar] [CrossRef] [PubMed]
- Weatherly, L.M.; Gosse, J.A. Triclosan exposure, transformation, and human health effects. J. Toxicol. Environ. Healthpart B 2017, 20, 447–469. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.L.; Vanlandingham, M.; da Costa, G.G.; Beland, F.A. Absorption and metabolism of triclosan after application to the skin of B 6 C 3 F 1 mice. Environ. Toxicol. 2016, 31, 609–623. [Google Scholar] [PubMed]
- Jung, E.M.; Ka, M.; Kim, W.Y. Loss of GSK-3 Causes Abnormal Astrogenesis and Behavior in Mice. Mol. Neurobiol. 2016, 53, 3954–3966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, D.N.; Jung, E.-M.; Yoo, Y.-M.; Ahn, C.; Kang, H.Y.; Choi, K.-C.; Hyun, S.-H.; Dang, V.H.; Pham, T.N.; Jeung, E.-B. Depletion of follicles accelerated by combined exposure to phthalates and 4-vinylcyclohexene diepoxide, leading to premature ovarian failure in rats. Reprod. Toxicol. 2018, 80, 60–67. [Google Scholar] [CrossRef]
- Edfawy, M.; Guedes, J.R.; Pereira, M.I.; Laranjo, M.; Carvalho, M.J.; Gao, X.; Ferreira, P.A.; Caldeira, G.; Franco, L.O.; Wang, D. Abnormal mGluR-mediated synaptic plasticity and autism-like behaviours in Gprasp2 mutant mice. Nat. Commun. 2019, 10, 1431. [Google Scholar] [CrossRef]
- Deacon, R.M. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, D.N.; Jung, E.-M.; Yoo, Y.-M.; Lee, J.-H.; Jeung, E.-B. Perinatal Exposure to Triclosan Results in Abnormal Brain Development and Behavior in Mice. Int. J. Mol. Sci. 2020, 21, 4009. https://doi.org/10.3390/ijms21114009
Tran DN, Jung E-M, Yoo Y-M, Lee J-H, Jeung E-B. Perinatal Exposure to Triclosan Results in Abnormal Brain Development and Behavior in Mice. International Journal of Molecular Sciences. 2020; 21(11):4009. https://doi.org/10.3390/ijms21114009
Chicago/Turabian StyleTran, Dinh Nam, Eui-Man Jung, Yeong-Min Yoo, Jae-Hwan Lee, and Eui-Bae Jeung. 2020. "Perinatal Exposure to Triclosan Results in Abnormal Brain Development and Behavior in Mice" International Journal of Molecular Sciences 21, no. 11: 4009. https://doi.org/10.3390/ijms21114009