Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite—Urolithin A—In the Brain

 , ,
, ,  , , , and
, , , and 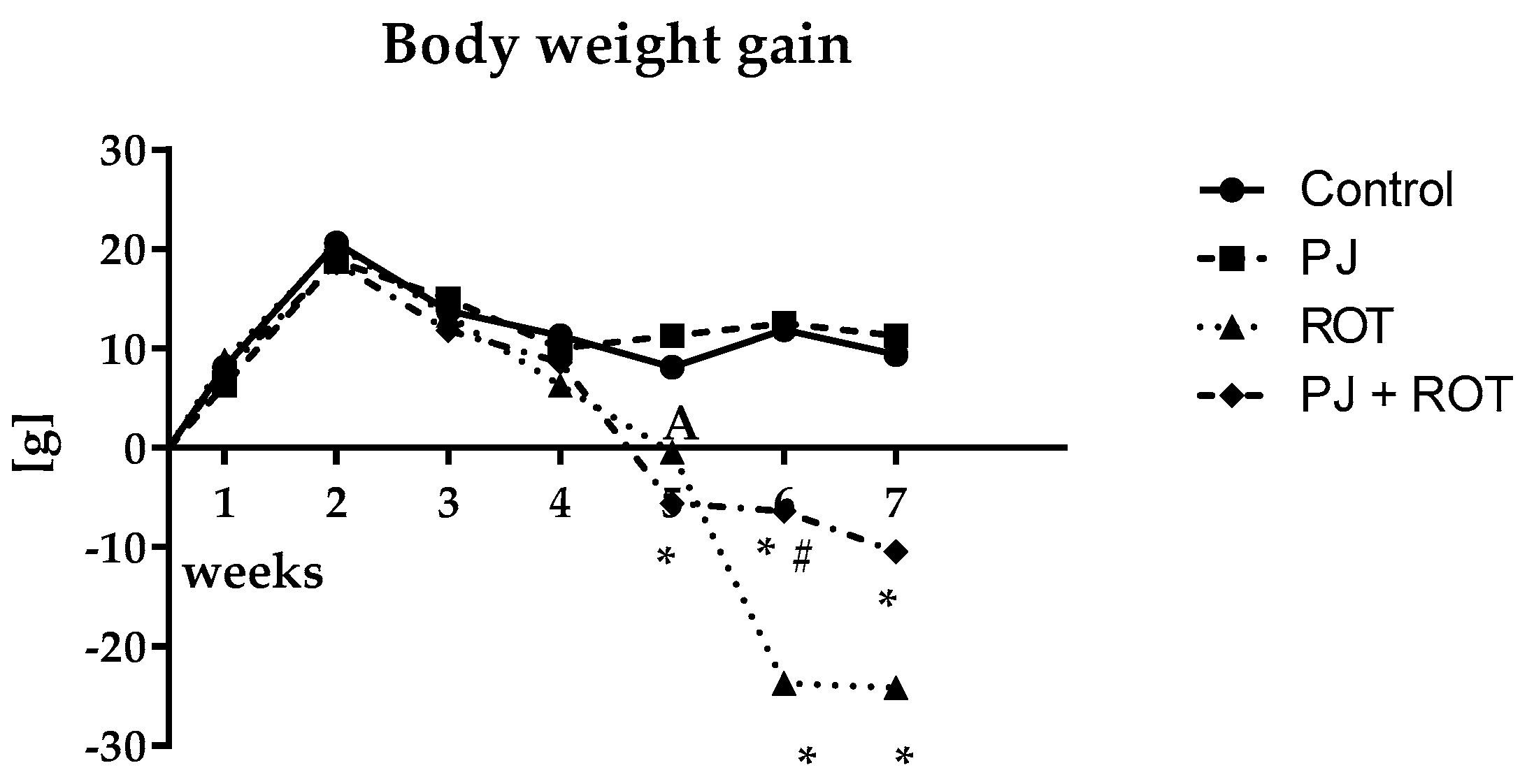{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
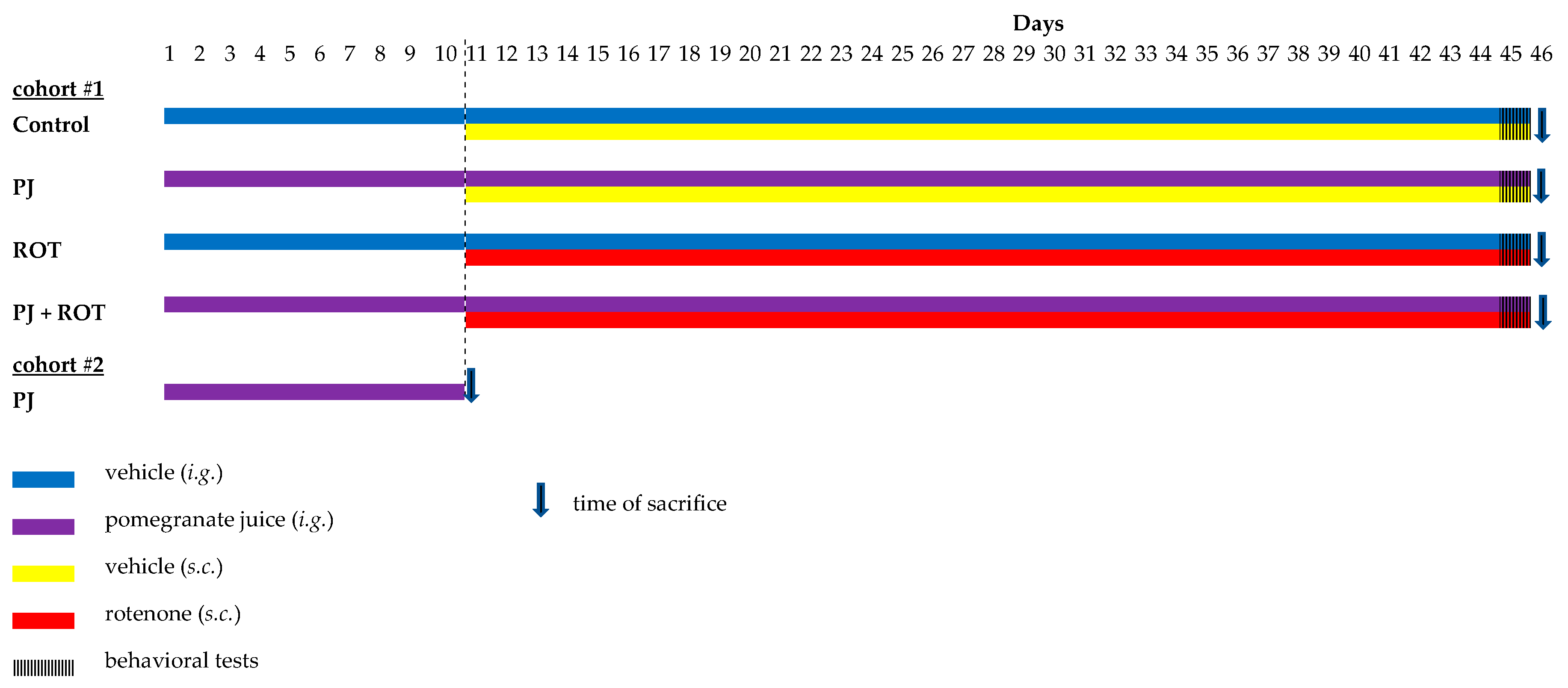{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bodyweight Gain
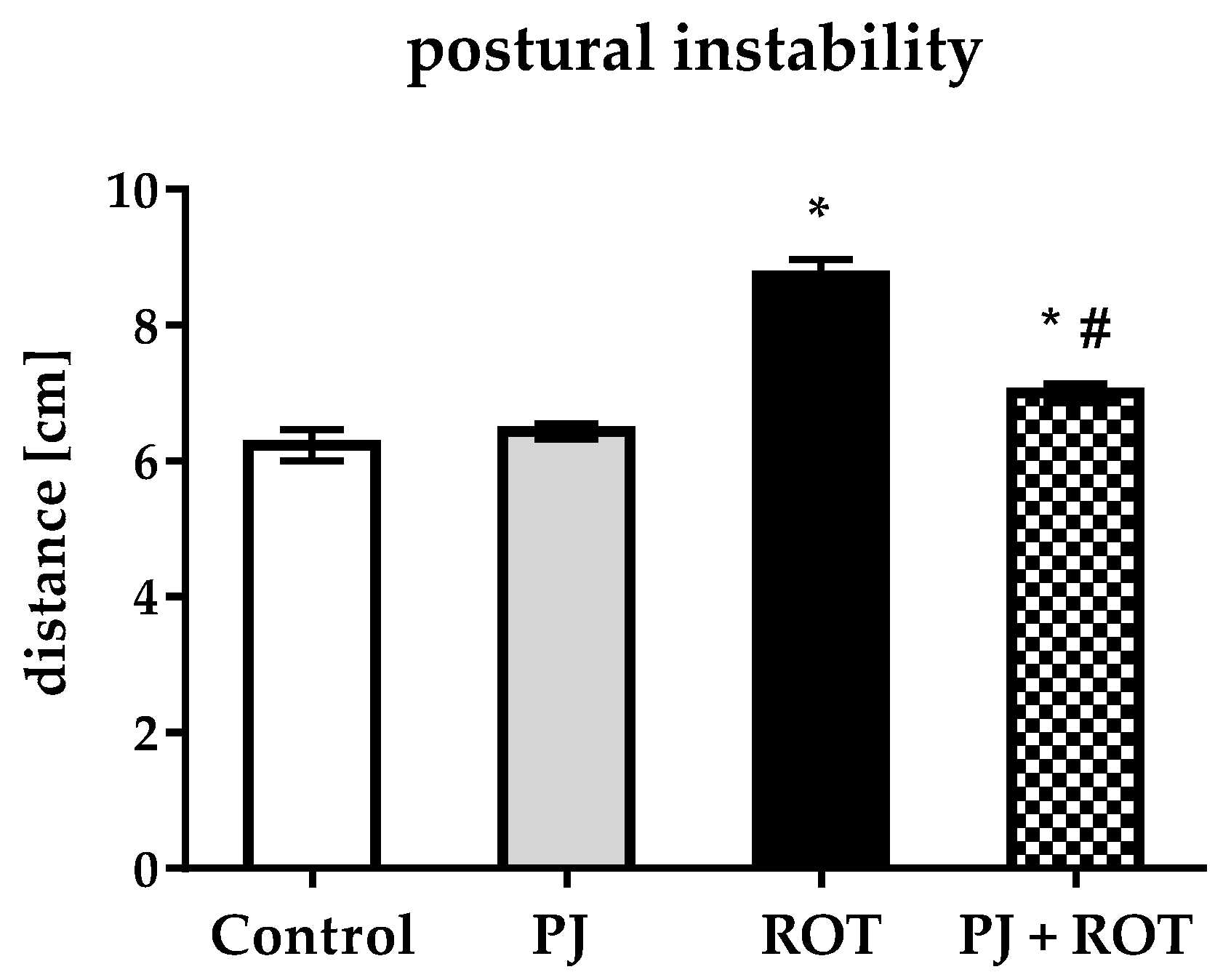
2.2. Postural Instability
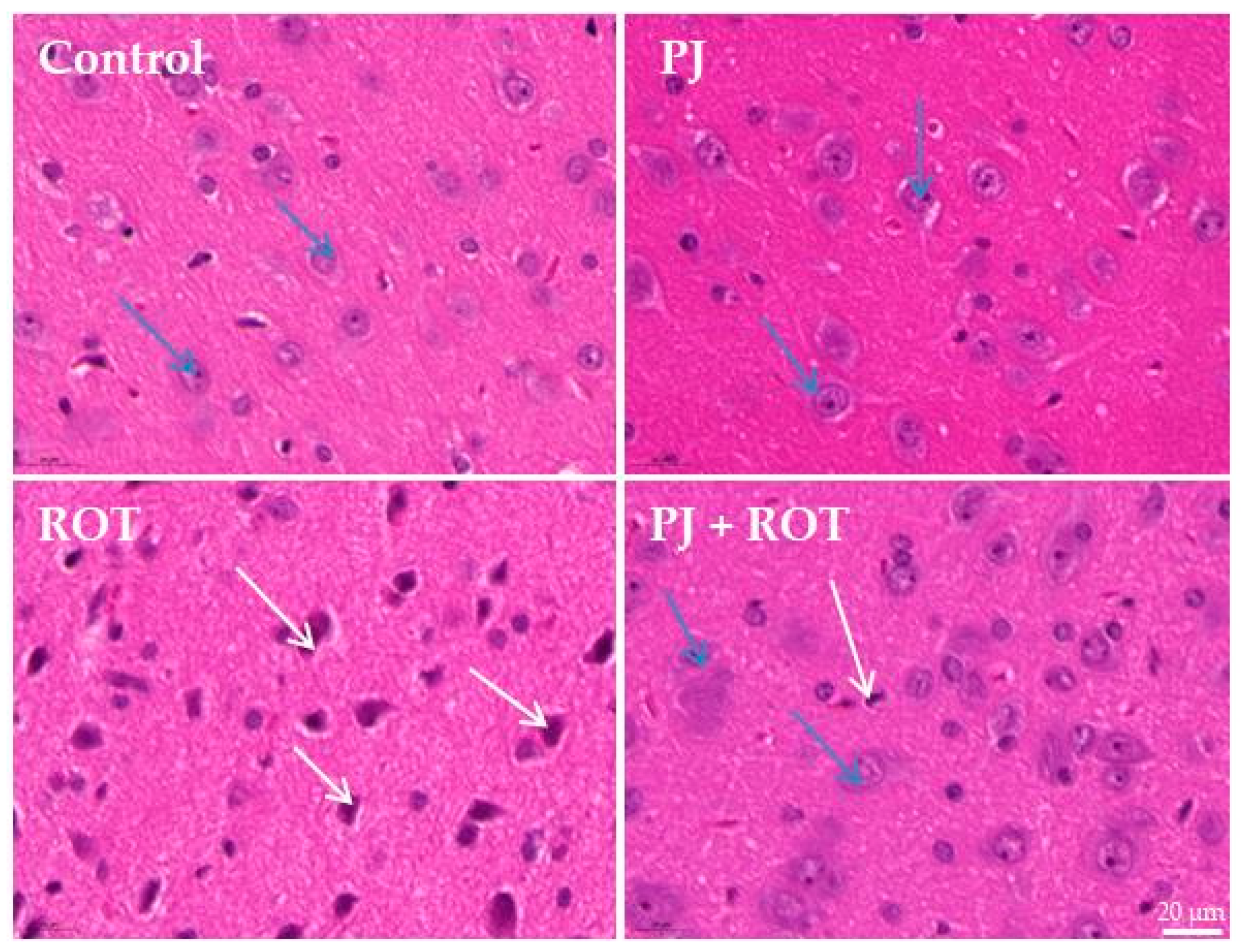
2.3. Microscopic Examination
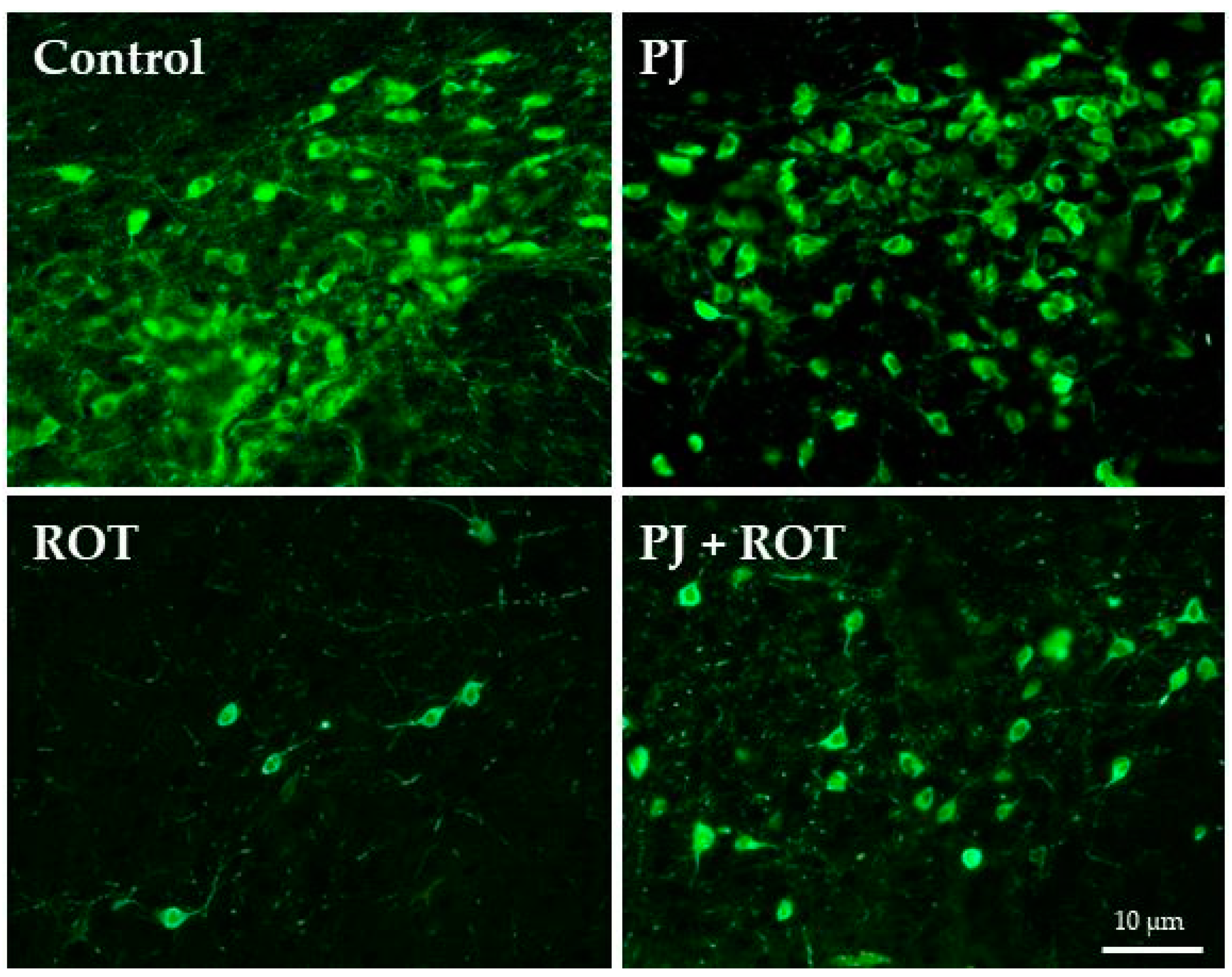
2.4. Immunofluorescence Staining of TH Positive (TH+) Neurons in the Region of SN
2.5. Biochemical Examinations
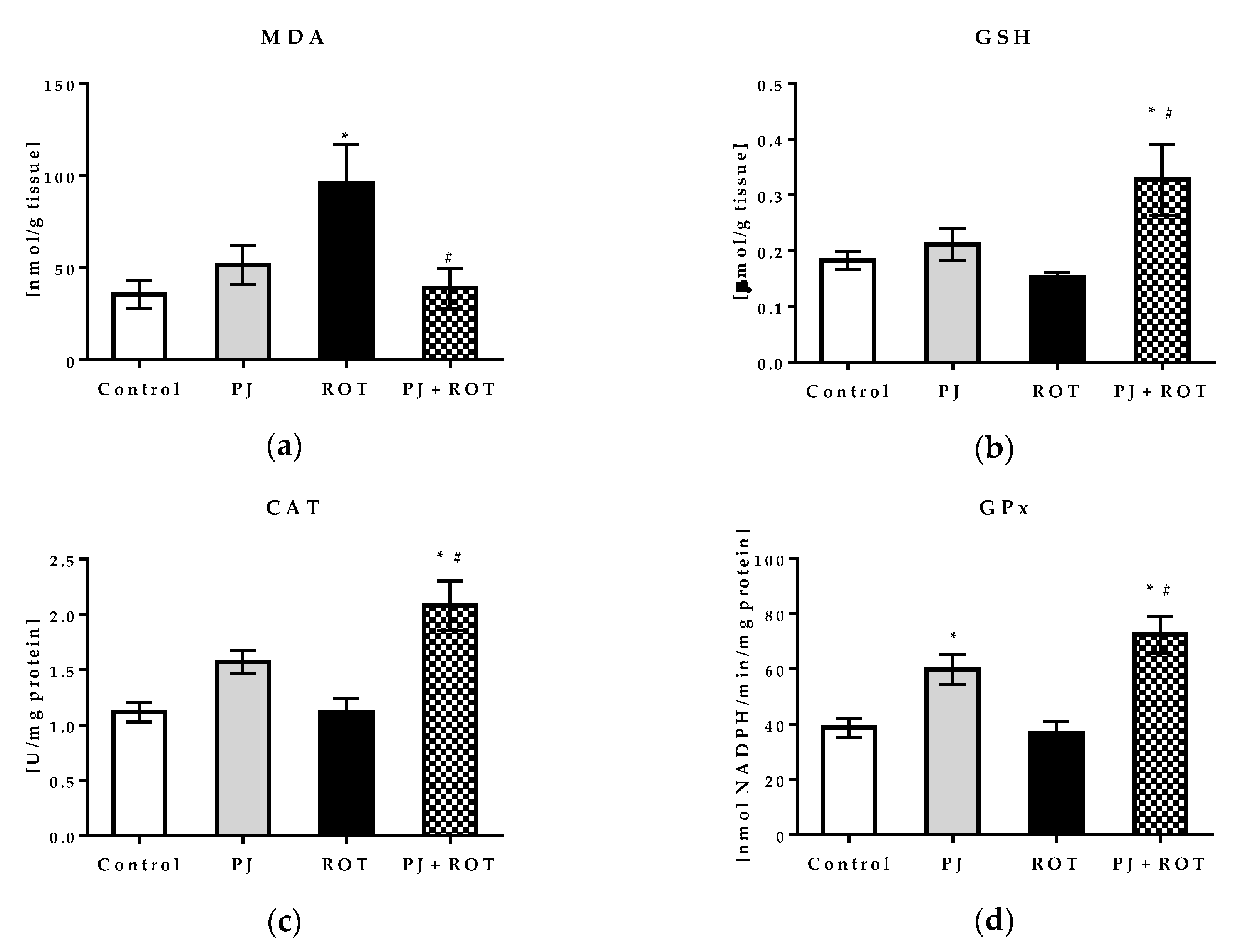
2.5.1. Oxidative Stress Markers and Mitochondrial Aldehyde Dehydrogenase Activity
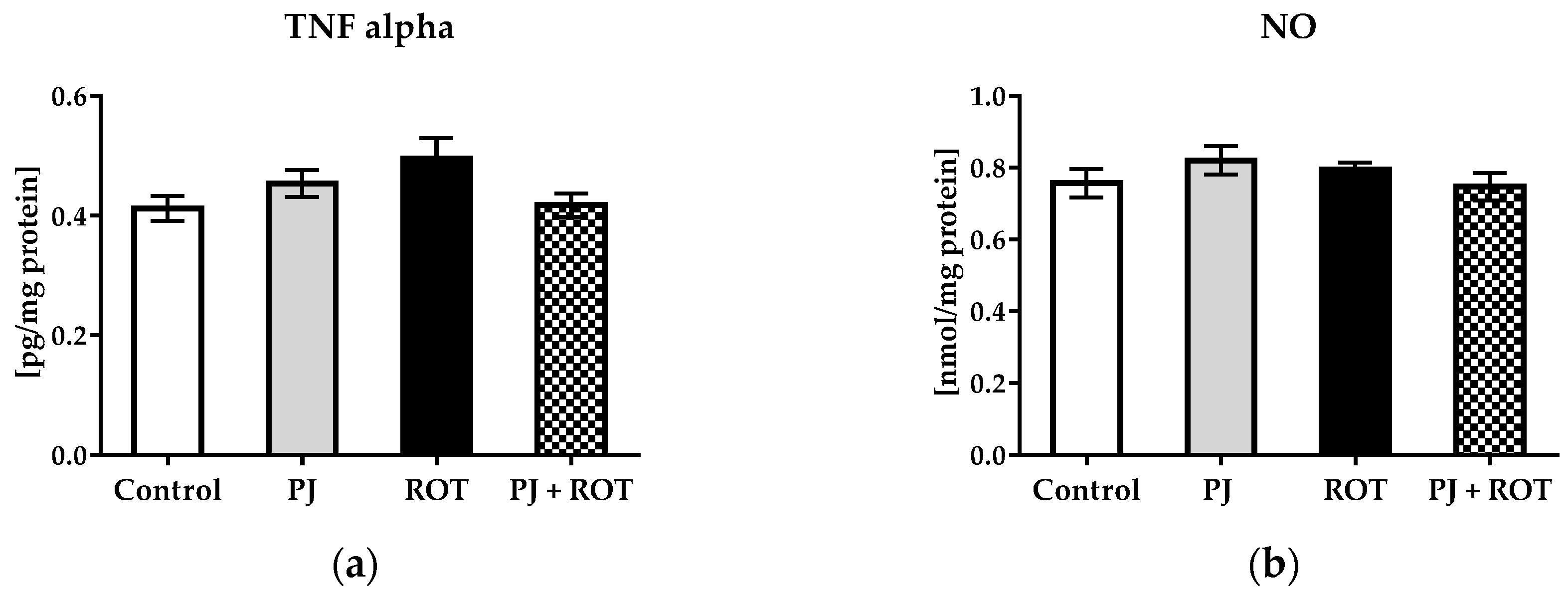
2.5.2. Inflammation Markers
2.6. Apoptosis Markers
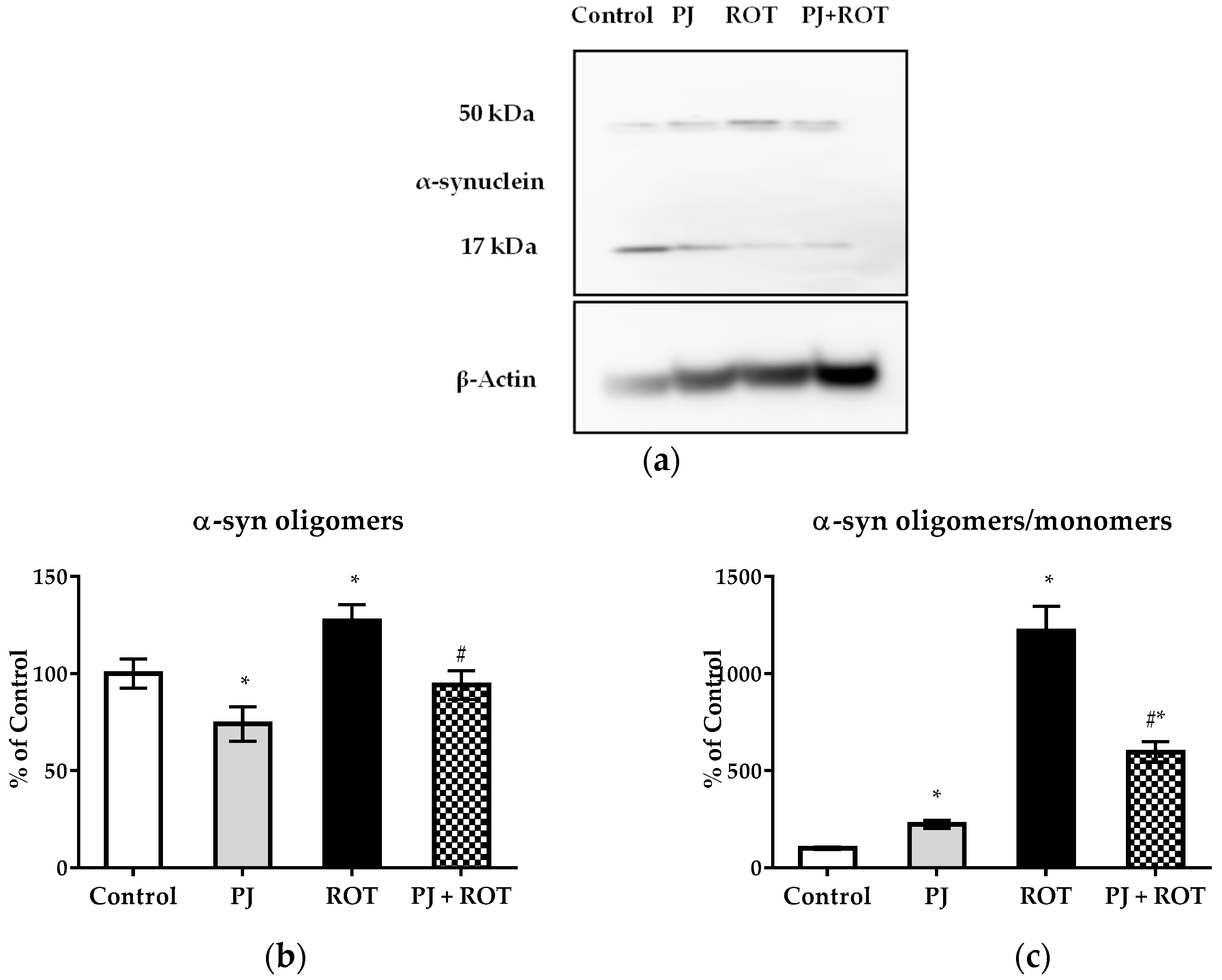
2.7. α-Synuclein Expression
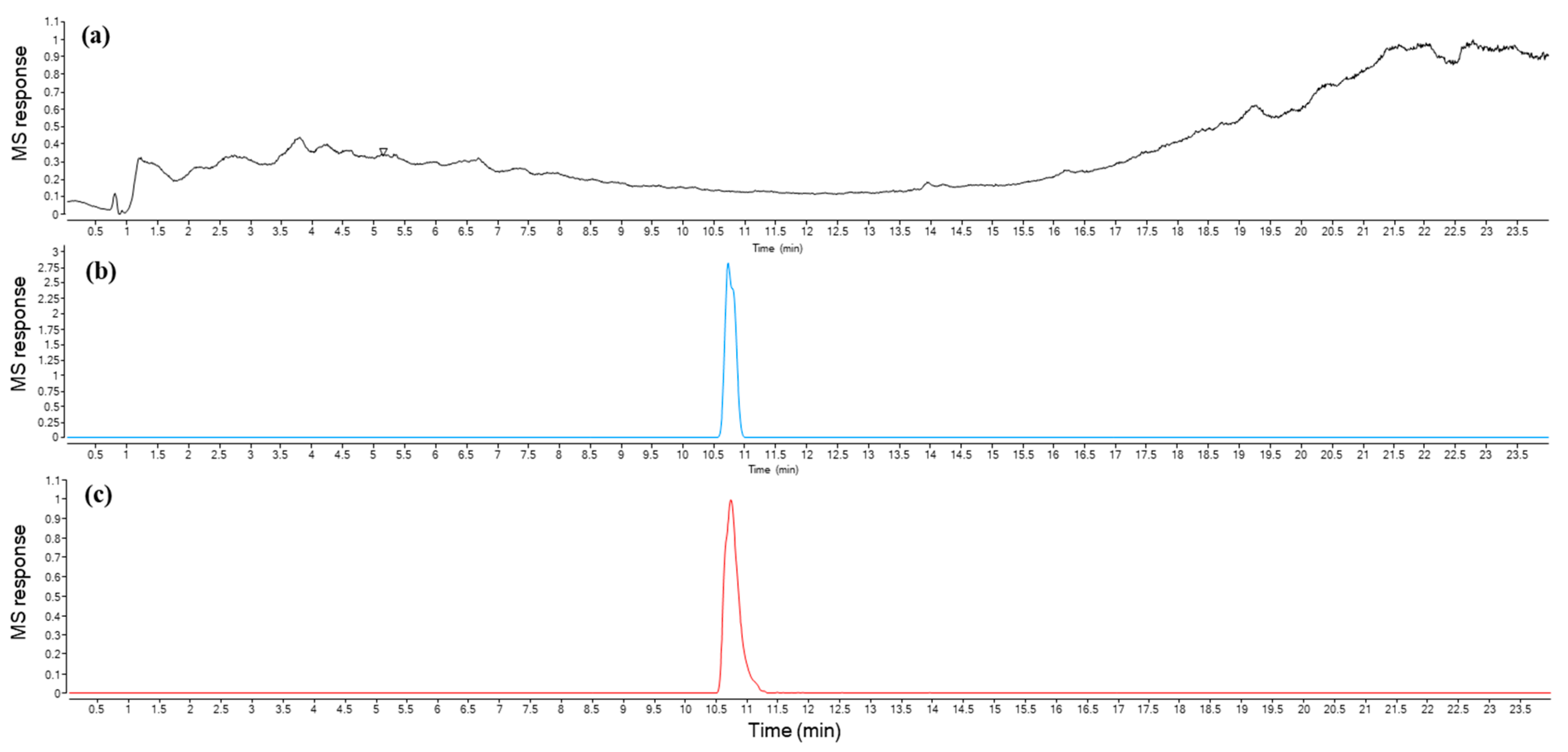
2.8. Urolithin a Determination
3. Discussion
4. Materials and Methods
4.1. Pomegranate Juice
4.2. Animals
4.3. Experimental Design
4.4. Postural Instability Test
4.5. Hematoxylin and Eosin (H&E) Staining
4.6. Immunofluorescent Staining of TH+ Neurons
4.7. Biochemical Examinations
4.7.1. Lipid Peroxidation
4.7.2. Endogenous Antioxidants
4.7.3. Mitochondrial Aldehyde Dehydrogenase (ALDH2) Activity
4.7.4. Tumor Necrosis Factor-Alpha (TNF-α)
4.7.5. Nitrite Concentration
4.7.6. Protein Determination
4.8. Western Blotting
4.9. Urolithin a Determination
4.9.1. Isolation
4.9.2. UPLC-ESI-QTOF-MS Analysis
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer disease |
| ALDH2 | mitochondrial aldehyde dehydrogenase |
| APP | amyloid precursor protein |
| Bax | Bcl-2 family member - apoptosis regulator |
| Bcl-xl | B-cell lymphoma-extra large |
| CAT | catalase |
| EA | ellagic acid |
| ESI | electrospray ionization |
| H&E | hematoxylin and eosin |
| ETs | ellagitannins |
| H-I | hypoxic-ischemic |
| Iba1 | ionized calcium-binding adapter molecule 1 |
| I/R | ischemia-reperfusion |
| GSH | reduced glutathione |
| GST | glutathione S-transferase |
| HFS | high-fat/sucrose |
| LPO | lipid peroxidation |
| MDA | malondialdehyde |
| MPP+ | 1-methyl-4-phenylpyridinium |
| MPTP | 4-phenyl-1, 2, 3, 6-tetrahydropyridine |
| NO | nitric oxide |
| NPS | normal Pig Serum |
| PD | Parkinson’s disease |
| PJ | pomegranate juice |
| ROS | reactive oxygen species |
| ROT | rotenone |
| SNpc | substantia nigra pars compacta |
| TH | tyrosine hydroxylase |
| TNF-α | tumor necrosis factor-alpha |
| UA | urolithin A |
References
- Kujawska, M.; Jodynis-Liebert, J. Polyphenols in Parkinson’s Disease: A Systematic Review of In Vivo Studies. Nutrients 2018, 10, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.L.; Kirk, R.D.; DaSilva, N.A.; Ma, H.; Seeram, N.P.; Bertin, M.J. Polyphenol Microbial Metabolites Exhibit Gut and Blood–Brain Barrier Permeability and Protect Murine Microglia against LPS-Induced Inflammation. Metabolites 2019, 9, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vegara, S.; Martí, N.; Lorente, J.; Coll, L.; Streitenberger, S.; Valero, M.; Saura, D. Chemical guide parameters for Punica granatum cv. ‘Mollar’ fruit juices processed at industrial scale. Food Chem. 2014, 147, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, Y.; Khatib, N.; Saadi, N.; Ross, M.G.; Weiner, Z.; Beloosesky, R. Maternal pomegranate juice attenuates maternal inflammation-induced fetal brain injury by inhibition of apoptosis, neuronal nitric oxide synthase, and NF-κB in a rat model. Am. J. Obstet. Gynecol. 2018, 219, 113.e1–113.e9. [Google Scholar] [CrossRef]
- West, T.; Atzeva, M.; Holtzman, D.M. Pomegranate polyphenols and resveratrol protect the neonatal brain against hypoxic-ischemic injury. Dev. Neurosci. 2007, 29, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Loren, D.J.; Seeram, N.P.; Schulman, R.N.; Holtzman, D.M. Maternal dietary supplementation with pomegranate juice is neuroprotective in an animal model of neonatal hypoxic-ischemic brain injury. Pediatr. Res. 2005, 57, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.A.; El Morsy, E.M.; Ahmed, A.A. Pomegranate extract protects against cerebral ischemia/reperfusion injury and preserves brain DNA integrity in rats. Life Sci. 2014, 110, 61–69. [Google Scholar] [CrossRef]
- Yuan, T.; Ma, H.; Liu, W.; Niesen, D.B.; Shah, N.; Crews, R.; Rose, K.N.; Vattem, D.A.; Seeram, N.P. Pomegranate’s Neuroprotective Effects against Alzheimer’s Disease Are Mediated by Urolithins, Its Ellagitannin-Gut Microbial Derived Metabolites. ACS Chem. Neurosci. 2016, 7, 26–33. [Google Scholar] [CrossRef]
- Rojanathammanee, L.; Puig, K.L.; Combs, C.K. Pomegranate polyphenols and extract inhibit nuclear factor of activated T-cell activity and microglial activation in vitro and in a transgenic mouse model of Alzheimer disease. J. Nutr. 2013, 143, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Essa, M.M.; Subash, S.; Akbar, M.; Al-Adawi, S.; Guillemin, G.J. Long-term dietary supplementation of pomegranates, figs and dates alleviate neuroinflammation in a transgenic mouse model of Alzheimer’s disease. PLoS ONE 2015, 10, 0120964. [Google Scholar] [CrossRef] [Green Version]
- Subash, S.; Braidy, N.; Essa, M.M.; Zayana, A.B.; Ragini, V.; Al-Adawi, S.; Al-Asmi, A.; Guillemin, G.J. Long-term (15 mo) dietary supplementation with pomegranates from Oman attenuates cognitive and behavioral deficits in a transgenic mice model of Alzheimer’s disease. Nutrition 2015, 31, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Subash, S.; Essa, M.M.; Al-Asmi, A.; Al-Adawi, S.; Vaishnav, R.; Braidy, N.; Manivasagam, T.; Guillemin, G.J. Pomegranate from Oman Alleviates the Brain Oxidative Damage in Transgenic Mouse Model of Alzheimer’s disease. J. Tradit. Complement. Med. 2014, 4, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.H.; Subaiea, G.M.; Eid, A.; Li, L.; Seeram, N.P.; Zawia, N.H. Pomegranate extract modulates processing of amyloid-β precursor protein in an aged Alzheimer’s disease animal model. Curr. Alzheimer Res. 2014, 11, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Hartman, R.E.; Shah, A.; Fagan, A.M.; Schwetye, K.E.; Parsadanian, M.; Schulman, R.N.; Finn, M.B.; Holtzman, D.M. Pomegranate juice decreases amyloid load and improves behavior in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2006, 24, 506–515. [Google Scholar] [CrossRef]
- Bookheimer, S.Y.; Renner, B.A.; Ekstrom, A.; Li, Z.; Henning, S.M.; Brown, J.A.; Jones, M.; Moody, T.; Small, G.W. Pomegranate juice augments memory and FMRI activity in middle-aged and older adults with mild memory complaints. Evid. Based Complement. Altern. Med. 2013, 2013, 946298. [Google Scholar] [CrossRef]
- Verzelloni, E.; Pellacani, C.; Tagliazucchi, D.; Tagliaferri, S.; Calani, L.; Costa, L.G.; Brighenti, F.; Borges, G.; Crozier, A.; Conte, A.; et al. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol. Nutr. Food Res. 2011, 55 (Suppl. 1), S35–S43. [Google Scholar] [CrossRef]
- Liu, W.; Ma, H.; Frost, L.; Yuan, T.; Dain, J.A.; Seeram, N.P. Pomegranate phenolics inhibit formation of advanced glycation endproducts by scavenging reactive carbonyl species. Food Funct. 2014, 5, 2996–3004. [Google Scholar] [CrossRef]
- Lee, G.; Park, J.S.; Lee, E.J.; Ahn, J.H.; Kim, H.S. Anti-inflammatory and antioxidant mechanisms of urolithin B in activated microglia. Phytomedicine 2019, 55, 50–57. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Núñez-Sánchez, M.; Tomás-Barberán, F.A.; Espín, J.C. Neuroprotective Effects of Bioavailable Polyphenol-Derived Metabolites against Oxidative Stress-Induced Cytotoxicity in Human Neuroblastoma SH-SY5Y Cells. J. Agric. Food Chem. 2017, 65, 752–758. [Google Scholar] [CrossRef] [Green Version]
- Velagapudi, R.; Lepiarz, I.; ElBakoush, A.; Katola, F.O.; Bhatia, H.; Fiebich, B.L.; Olajide, O. Induction of Autophagy and Activation of SIRT-1 Deacetylation Mechanisms Mediate Neuroprotection by the Pomegranate Metabolite Urolithin A in BV2 Microglia and Differentiated 3D Human Neural Progenitor Cells. Mol. Nutr. Food Res. 2019, 1801237. [Google Scholar] [CrossRef] [Green Version]
- Hajipour, S.; Sarkaki, A.; Mohammad, S.; Mansouri, T.; Pilevarian, A.; RafieiRad, M. Motor and cognitive deficits due to permanent cerebral hypoperfusion/ischemia improve by pomegranate seed extract in rats. Pak. J. Biol. Sci. 2014, 17, 991–998. [Google Scholar] [PubMed] [Green Version]
- Tapias, V.; Cannon, J.R.; Greenamyre, J.T. Pomegranate juice exacerbates oxidative stress and nigrostriatal degeneration in Parkinson’s disease. Neurobiol. Aging 2014, 35, 1162–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaee, S.; Jahromy, M.H. Potential Effects of Pomegranate Juice in Attenuating LID in Mice Model of Parkinson Disease. Pharmacogn. J. 2018, 10, 700–704. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.D.; Zhao, X.; Li, Y.; Li, G.R.; Liu, X.L. Damage to dopaminergic neurons by oxidative stress in Parkinson’s disease (Review). Int. J. Mol. Med. 2018, 41, 1817–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Xia, Y.; Wan, F.; Ma, K.; Guo, X.; Kou, L.; Yin, S.; Han, C.; Liu, L.; Huang, J.; et al. New Perspectives on Roles of Alpha-Synuclein in Parkinson’s Disease. Front. Aging Neurosci. 2018, 10, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, S.; Freyberger, A.; Lawrenz, B.; Schladt, L.; Schmuck, G.; Ellinger-Ziegelbauer, H. Mechanistic Investigations of the Mitochondrial Complex I Inhibitor Rotenone in the Context of Pharmacological and Safety Evaluation. Sci. Rep. 2017, 7, 45465. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.C.; Yeh, T.H.; Lai, S.C.; Wu-Chou, Y.H.; Chen, C.H.; Mochly-Rosen, D.; Huang, Y.C.; Chen, Y.J.; Chen, C.L.; Chang, Y.M.; et al. Neuroprotective effects of aldehyde dehydrogenase 2 activation in rotenone-induced cellular and animal models of parkinsonism. Exp. Neurol. 2015, 263, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Manna, J.; Dunbar, G.L. Current understanding of the molecular mechanisms in Parkinson’s disease: Targets for potential treatments. Transl. Neurodegener. 2017, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Curtis, J.A.; Molfenter, S.; Troche, M.S. Predictors of Residue and Airway Invasion in Parkinson’s Disease. Dysphagia 2019. [Google Scholar] [CrossRef]
- Cannon, J.R.; Tapias, V.; Na, H.M.; Honick, A.S.; Drolet, R.E.; Greenamyre, J.T. A highly reproducible rotenone model of Parkinson’s disease. Neurobiol. Dis. 2009, 34, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Thakur, P.; Nehru, B. Modulatory effects of sodium salicylate on the factors affecting protein aggregation during rotenone induced Parkinson’s disease pathology. Neurochem. Int. 2014, 75, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Nehru, B. Beneficial Effect of Vitamin E in Rotenone Induced Model of PD: Behavioural, Neurochemical and Biochemical Study. Exp. Neurobiol. 2013, 22, 214–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binienda, Z.K.; Sarkar, S.; Mohammed-Saeed, L.; Gough, B.; Beaudoin, M.A.; Ali, S.F.; Paule, M.G.; Imam, S.Z. Chronic exposure to rotenone, a dopaminergic toxin, results in peripheral neuropathy associated with dopaminergic damage. Neurosci. Lett. 2013, 541, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Li, X.; Zhu, H. Curcumin ameliorates dopaminergic neuronal oxidative damage via activation of the Akt/Nrf2 pathway. Mol. Med. Rep. 2016, 13, 1381–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhang, D.Q.; Liao, Z.; Wang, B.; Gong, S.; Wang, C.; Zhang, M.Z.; Wang, G.H.; Cai, H.; Liao, F.F.; et al. Anti-oxidant polydatin (piceid) protects against substantia nigral motor degeneration in multiple rodent models of Parkinson’s disease. Mol. Neurodegener. 2015, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Kim, J. Degeneration of Dopaminergic Neurons Due to Metabolic Alterations and Parkinson’s Disease. Front. Aging Neurosci. 2016, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Kreiner, G.; Rafa-Zabłocka, K.; Barut, J.; Chmielarz, P.; Kot, M.; Bagińska, M.; Parlato, R.; Daniel, W.A.; Nalepa, I. Stimulation of noradrenergic transmission by reboxetine is beneficial for a mouse model of progressive parkinsonism. Sci. Rep. 2019, 9, 5262. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, B.; Sun, H.; Zhou, Y.; Liu, M.; Ding, J.; Fang, F.; Fan, Y.; Hu, G. Aquaporin-4 deficiency diminishes the differential degeneration of midbrain dopaminergic neurons in experimental Parkinson’s disease. Neurosci. Lett. 2016, 614, 7–15. [Google Scholar] [CrossRef]
- Khadrawy, Y.A.; Salem, A.M.; El-Shamy, K.A.; Ahmed, E.K.; Fadl, N.N.; Hosny, E.N. Neuroprotective and Therapeutic Effect of Caffeine on the Rat Model of Parkinson’s Disease Induced by Rotenone. J. Diet. Suppl. 2017, 14, 553–572. [Google Scholar] [CrossRef]
- Khurana, N.; Gajbhiye, A. Ameliorative effect of Sida cordifolia in rotenone induced oxidative stress model of Parkinson’s disease. Neurotoxicology 2013, 39, 57–64. [Google Scholar] [CrossRef]
- Javed, H.; Azimullah, S.; Meeran, M.F.N.; Ansari, S.A.; Ojha, S. Neuroprotective Effects of Thymol, a Dietary Monoterpene Against Dopaminergic Neurodegeneration in Rotenone-Induced Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraj, R.L.; Beiram, R.; Azimullah, S.; Meeran, M.F.N.; Ojha, S.K.; Adem, A.; Jalal, F.Y. Lycopodium Attenuates Loss of Dopaminergic Neurons by Suppressing Oxidative Stress and Neuroinflammation in a Rat Model of Parkinson’s Disease. Molecules 2019, 24, 2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaidikar, L.; Byna, B.; Thakur, S.R. Neuroprotective effect of punicalagin against cerebral ischemia reperfusion-induced oxidative brain injury in rats. J. Stroke Cerebrovasc. Dis. 2014, 23, 2869–2878. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, A.; Zheng, Y.R.; Wu, X.L.; Tang, W.D.; Liu, M.R.; Ma, S.J.; Jiang, L.; Hu, W.W.; Zhang, X.N.; Chen, Z. Urolithin A-activated autophagy but not mitophagy protects against ischemic neuronal injury by inhibiting ER stress in vitro and in vivo. CNS Neurosci. Ther. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.H.; Zhang, H.F.; Yang, Z.B.; Li, T.B.; Liu, B.; Lou, Z.; Ma, Q.L.; Luo, X.J.; Peng, J. Alda-1 reduces cerebral ischemia/reperfusion injury in rat through clearance of reactive aldehydes. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Bernier, M.; Wahl, D.; Ali, A.; Allard, J.; Faulkner, S.; Wnorowski, A.; Sanghvi, M.; Moaddel, R.; Alfaras, I.; Mattison, J.A.; et al. Resveratrol supplementation confers neuroprotection in cortical brain tissue of nonhuman primates fed a high-fat/sucrose diet. Aging 2016, 8, 899–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Reus, M.I.; Gómez del Rio, M.A.; Iglesias, I.; Elorza, M.; Slowing, K.; Benedí, J. Standardized Hypericum perforatum reduces oxidative stress and increases gene expression of antioxidant enzymes on rotenone-exposed rats. Neuropharmacology 2007, 52, 606–616. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Abul Khair, S.B.; Haque, M.E. Neuroprotective potential of ferulic acid in the rotenone model of Parkinson’s disease. Drug Des. Dev. Ther. 2015, 9, 5499–5510. [Google Scholar] [CrossRef] [Green Version]
- Palle, S.; Neerati, P. Improved neuroprotective effect of resveratrol nanoparticles as evinced by abrogation of rotenone-induced behavioral deficits and oxidative and mitochondrial dysfunctions in rat model of Parkinson’s disease. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 445–453. [Google Scholar] [CrossRef]
- El-Sayed, E.K.; Ahmed, A.; Morsy, E.E.; Nofal, S. Neuroprotective effect of agmatine (decarboxylated l-arginine) against oxidative stress and neuroinflammation in rotenone model of Parkinson’s disease. Hum. Exp. Toxicol. 2019, 38, 173–184. [Google Scholar] [CrossRef]
- Manjunath, M.J. Effect of Withania somnifera supplementation on rotenone-induced oxidative damage in cerebellum and striatum of the male mice brain. Cent. Nerv. Syst. Agents Med. Chem. 2013, 13, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amri, Z.; Ghorbel, A.; Turki, M.; Akrout, F.M.; Ayadi, F.; Elfeki, A.; Hammami, M. Effect of pomegranate extracts on brain antioxidant markers and cholinesterase activity in high fat-high fructose diet induced obesity in rat model. BMC Complement. Altern. Med. 2017, 17, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morzelle, M.C.; Salgado, J.M.; Telles, M.; Mourelle, D.; Bachiega, P.; Buck, H.S.; Viel, T.A. Neuroprotective Effects of Pomegranate Peel Extract after Chronic Infusion with Amyloid-β Peptide in Mice. PLoS ONE 2016, 11, 0166123. [Google Scholar] [CrossRef] [PubMed]
- El-Missiry, M.A.; ElKomy, M.A.; Othman, A.I.; AbouEl-Ezz, A.M. Punicalagin ameliorates the elevation of plasma homocysteine, amyloid-β, TNF-α and apoptosis by advocating antioxidants and modulating apoptotic mediator proteins in brain. Biomed. Pharmacother. 2018, 102, 472–480. [Google Scholar] [CrossRef]
- Hassimotto, N.M.; Lajolo, F.M. Antioxidant status in rats after long-term intake of anthocyanins and ellagitannins from blackberries. J. Sci. Food Agric. 2011, 91, 523–531. [Google Scholar] [CrossRef]
- Sun, W.; Yan, C.; Frost, B.; Wang, X.; Hou, C.; Zeng, M.; Gao, H.; Kang, Y.; Liu, J. Pomegranate extract decreases oxidative stress and alleviates mitochondrial impairment by activating AMPK-Nrf2 in hypothalamic paraventricular nucleus of spontaneously hypertensive rats. Sci. Rep. 2016, 6, 34246. [Google Scholar] [CrossRef] [Green Version]
- Zaitone, S.A.; Abo-Elmatty, D.M.; Shaalan, A.A. Acetyl-L-carnitine and α-lipoic acid affect rotenone-induced damage in nigral dopaminergic neurons of rat brain, implication for Parkinson’s disease therapy. Pharmacol. Biochem. Behav. 2012, 100, 347–360. [Google Scholar] [CrossRef]
- Ethell, D.W.; Fei, Q. Parkinson-linked genes and toxins that affect neuronal cell death through the Bcl-2 family. Antioxid. Redox Signal. 2009, 11, 529–540. [Google Scholar] [CrossRef]
- Horowitz, J.M.; Pastor, D.M.; Goyal, A.; Kar, S.; Ramdeen, N.; Hallas, B.H.; Torres, G. BAX protein-immunoreactivity in midbrain neurons of Parkinson’s disease patients. Brain Res. Bull. 2003, 62, 55–61. [Google Scholar] [CrossRef]
- Dhanalakshmi, C.; Janakiraman, U.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M.; Kalandar, A.; Khan, M.A.; Guillemin, G.J. Vanillin Attenuated Behavioural Impairments, Neurochemical Deficts, Oxidative Stress and Apoptosis Against Rotenone Induced Rat Model of Parkinson’s Disease. Neurochem. Res. 2016, 41, 1899–1910. [Google Scholar] [CrossRef]
- Kandil, E.A.; Sayed, R.H.; Ahmed, L.A.; Abd El Fattah, M.A.; El-Sayeh, B.M. Hypoxia-inducible factor 1 alpha and nuclear-related receptor 1 as targets for neuroprotection by albendazole in a rat rotenone model of Parkinson’s disease. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Zhou, Q.; Zhang, W.; Fu, X.; Wu, Q.; Lu, Y.; Shi, J.; Zhou, S. Icariin-mediated activation of autophagy confers protective effect on rotenone induced neurotoxicity in vivo and in vitro. Toxicol. Rep. 2019, 6, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Aronson, W.J.; Zhang, Y.; Henning, S.M.; Moro, A.; Lee, R.P.; Sartippour, M.; Harris, D.M.; Rettig, M.; Suchard, M.A.; et al. Pomegranate ellagitannin-derived metabolites inhibit prostate cancer growth and localize to the mouse prostate gland. J. Agric. Food Chem. 2007, 55, 7732–7737. [Google Scholar] [CrossRef] [PubMed]
- Oszmiański, J.; Wojdyło, A.; Nowicka, P.; Teleszko, M.; Cebulak, T.; Wolanin, M. Determination of phenolic compounds and antioxidant activity in leaves from wild Rubus, L. species. Molecules 2015, 20, 4951–4966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Salam, O.M.E.; Youness, E.R.; Ahmed, N.A.; El-Toumy, S.A.; Souleman, A.M.A.; Shaffie, N.; Abouelfadl, D.M. Bougainvillea spectabilis flowers extract protects against the rotenone-induced toxicity. Asian Pac. J. Trop. Med. 2017, 10, 478–490. [Google Scholar] [CrossRef]
- Kujawska, M.; Ignatowicz, E.; Ewertowska, M.; Adamska, T.; Markowski, J.; Jodynis-Liebert, J. Attenuation of KBrO3-induced renal and hepatic toxicity by cloudy apple juice in rat. Phytother. Res. 2013, 27, 1214–1219. [Google Scholar] [CrossRef]
- Gilchrist, M.; Savoie, M.; Nohara, O.; Wills, F.L.; Wallace, J.L.; Befus, A.D. Nitric oxide synthase and nitric oxide production in in vivo-derived mast cells. J. Leukoc. Biol. 2002, 71, 618–624. [Google Scholar]
- Nuñez-Sánchez, M.A.; García-Villalba, R.; Monedero-Saiz, T.; García-Talavera, N.V.; Gómez-Sánchez, M.B.; Sánchez-Álvarez, C.; García-Albert, A.M.; Rodríguez-Gil, F.J.; Ruiz-Marín, M.; Pastor-Quirante, F.A.; et al. Targeted metabolic profiling of pomegranate polyphenols and urolithins in plasma, urine and colon tissues from colorectal cancer patients. Mol. Nutr. Food Res. 2014, 58, 1199–1211. [Google Scholar] [CrossRef]
- Seeram, N.P.; Henning, S.M.; Zhang, Y.; Suchard, M.; Li, Z.; Heber, D. Pomegranate juice ellagitannin metabolites are present in human plasma and some persist in urine for up to 48 hours. J. Nutr. 2006, 136, 2481–2485. [Google Scholar] [CrossRef] [Green Version]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kujawska, M.; Jourdes, M.; Kurpik, M.; Szulc, M.; Szaefer, H.; Chmielarz, P.; Kreiner, G.; Krajka-Kuźniak, V.; Mikołajczak, P.Ł.; Teissedre, P.-L.; et al. Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite—Urolithin A—In the Brain. Int. J. Mol. Sci. 2020, 21, 202. https://doi.org/10.3390/ijms21010202
Kujawska M, Jourdes M, Kurpik M, Szulc M, Szaefer H, Chmielarz P, Kreiner G, Krajka-Kuźniak V, Mikołajczak PŁ, Teissedre P-L, et al. Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite—Urolithin A—In the Brain. International Journal of Molecular Sciences. 2020; 21(1):202. https://doi.org/10.3390/ijms21010202
Chicago/Turabian StyleKujawska, Małgorzata, Michael Jourdes, Monika Kurpik, Michał Szulc, Hanna Szaefer, Piotr Chmielarz, Grzegorz Kreiner, Violetta Krajka-Kuźniak, Przemyslaw Łukasz Mikołajczak, Pierre-Louis Teissedre, and et al. 2020. "Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite—Urolithin A—In the Brain" International Journal of Molecular Sciences 21, no. 1: 202. https://doi.org/10.3390/ijms21010202