Comparative Physiological Analysis Reveals the Role of NR-Derived Nitric Oxide in the Cold Tolerance of Forage Legumes


Abstract
:1. Introduction
2. Results
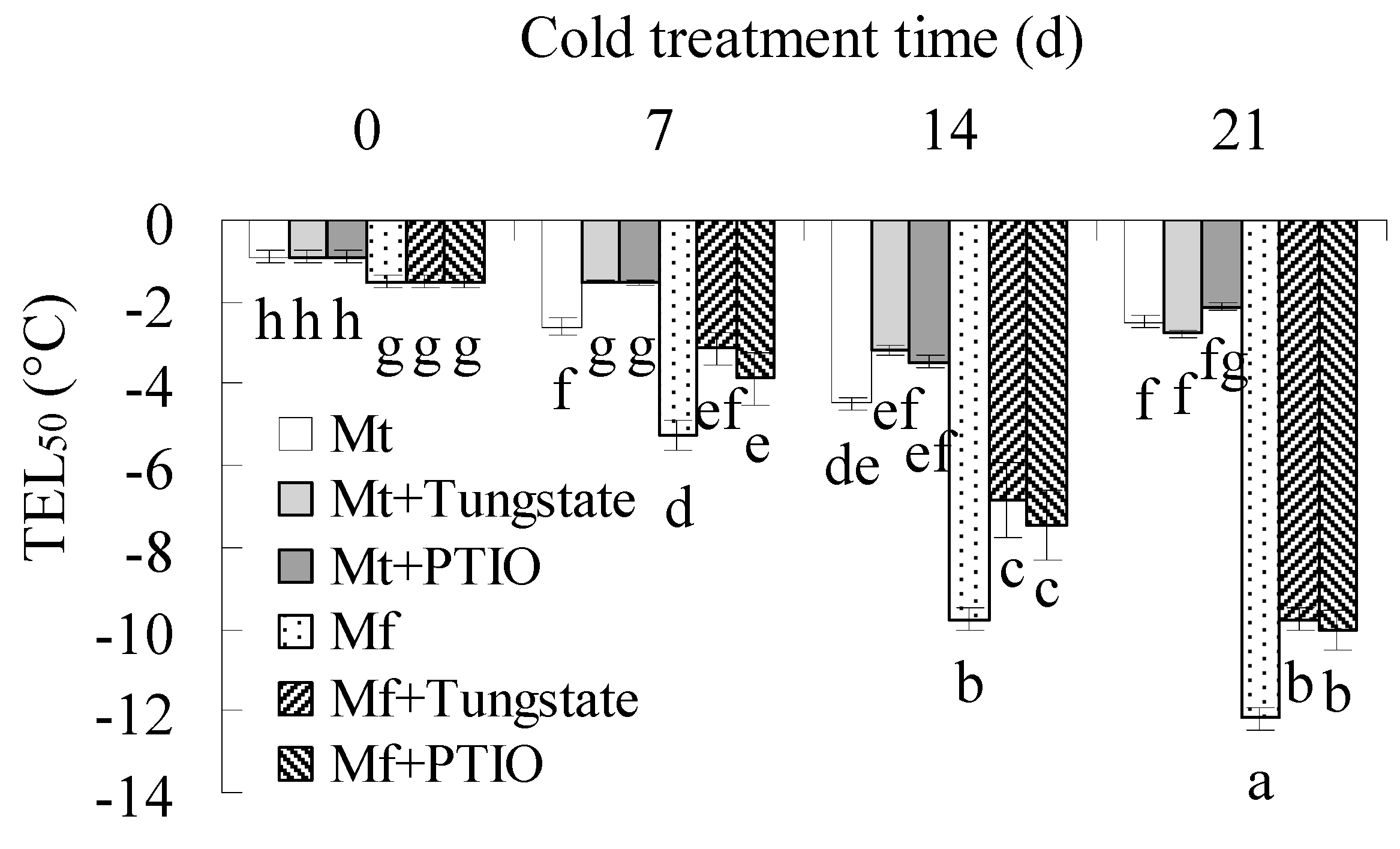
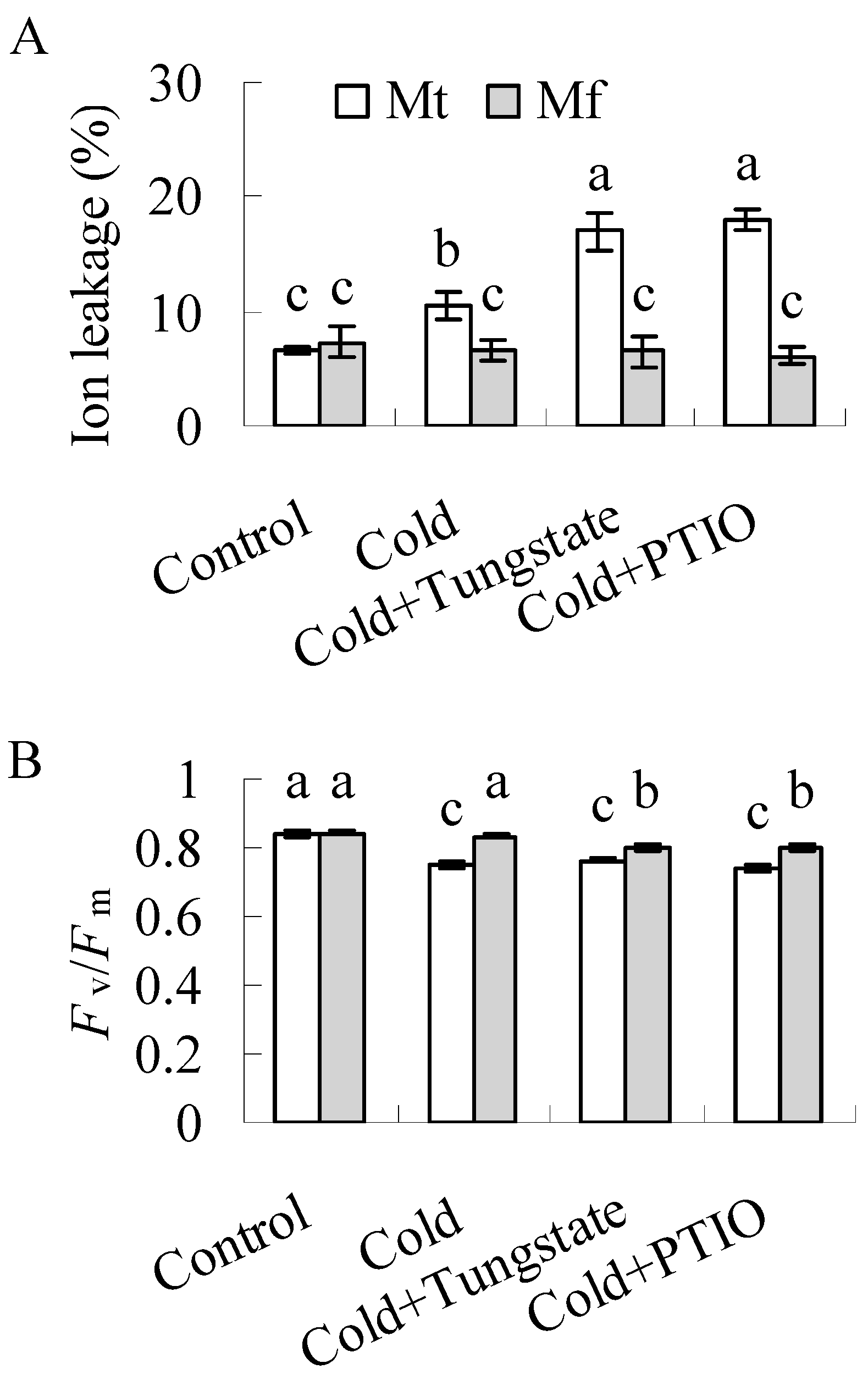
2.1. Differential Cold Tolerance in M. falcata and M. truncatula
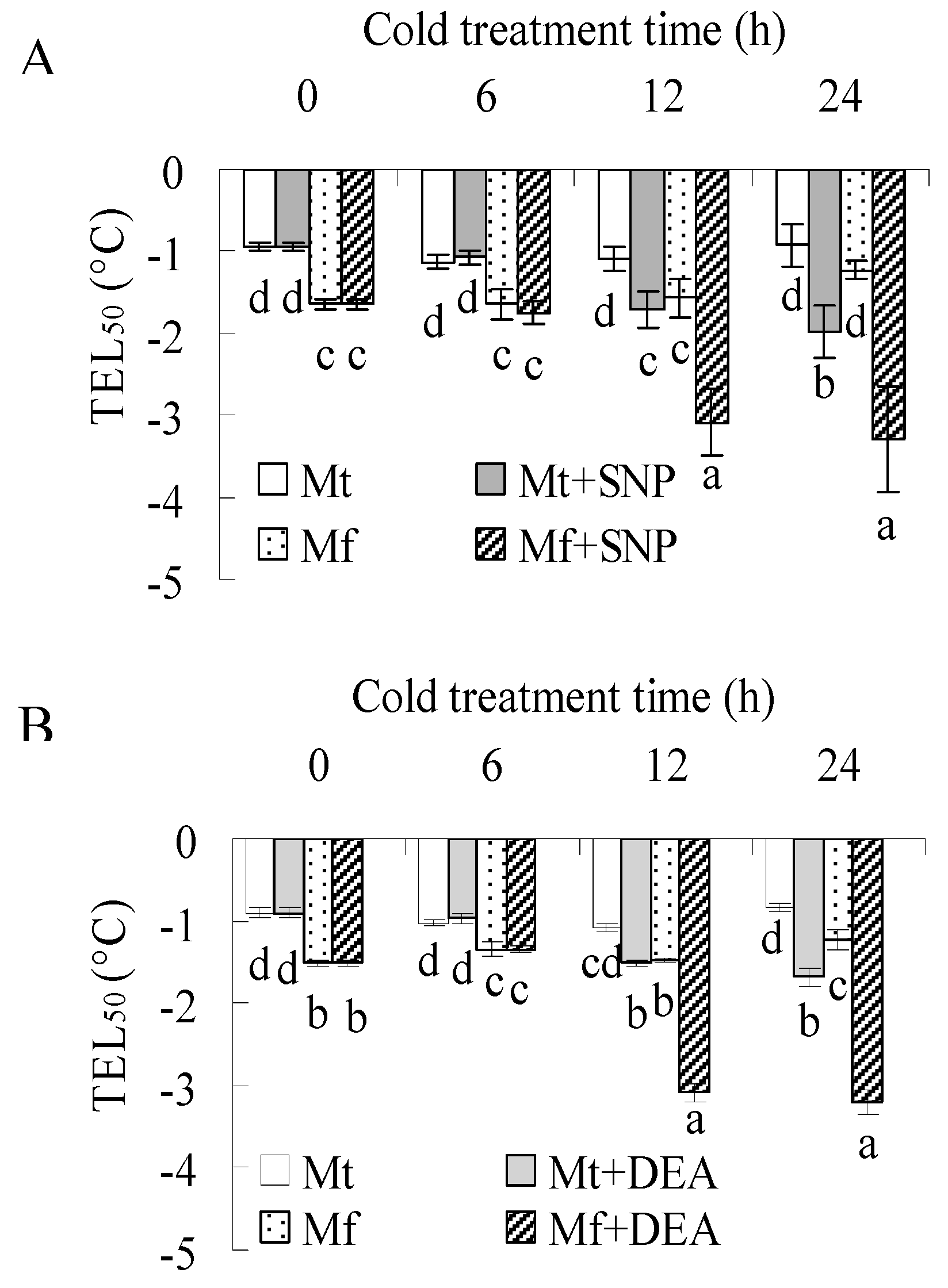
2.2. NO is Involved in Cold Acclimation of M. falcata and M. truncatula
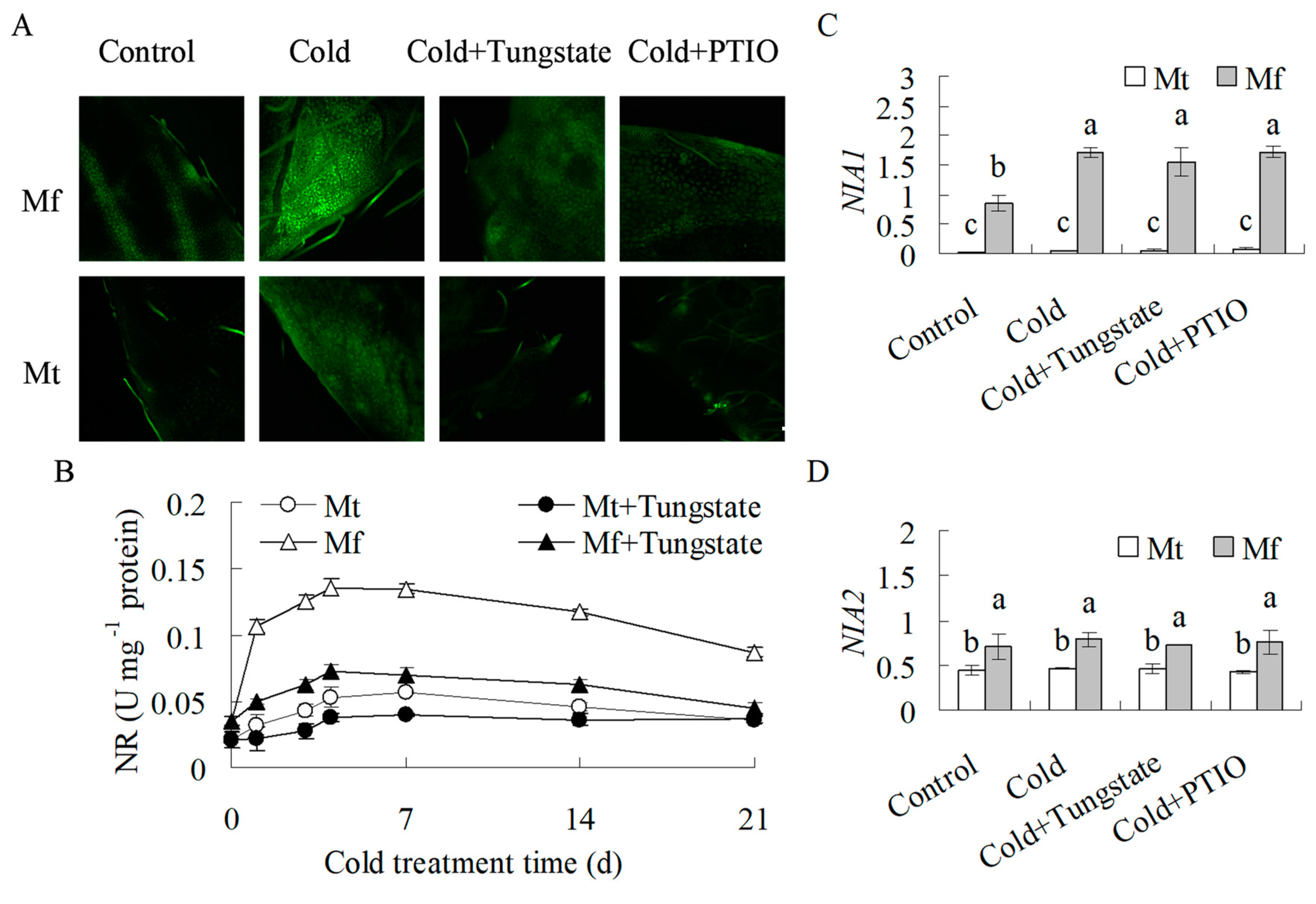
2.3. NO Production was Induced during Cold Treatment
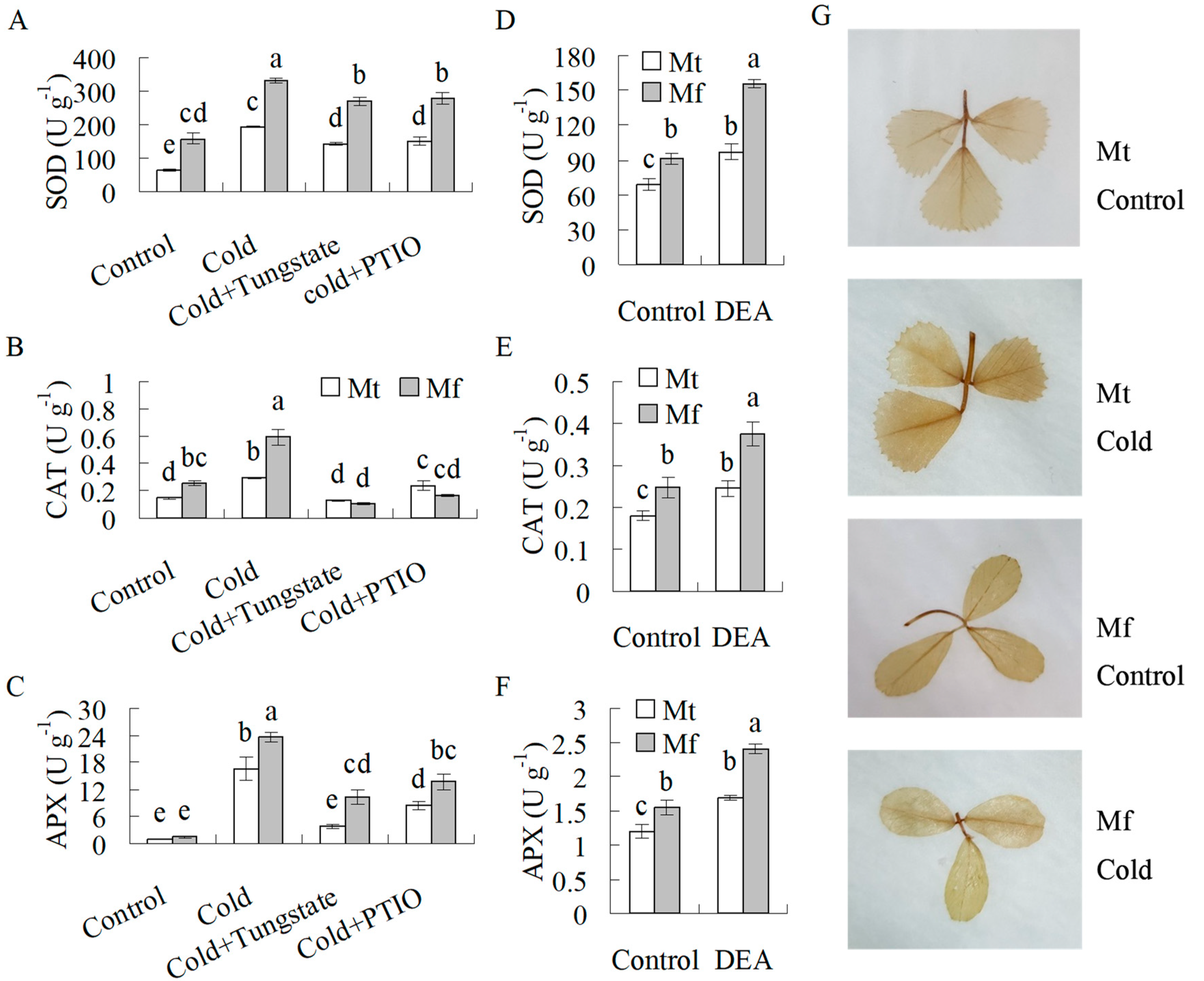
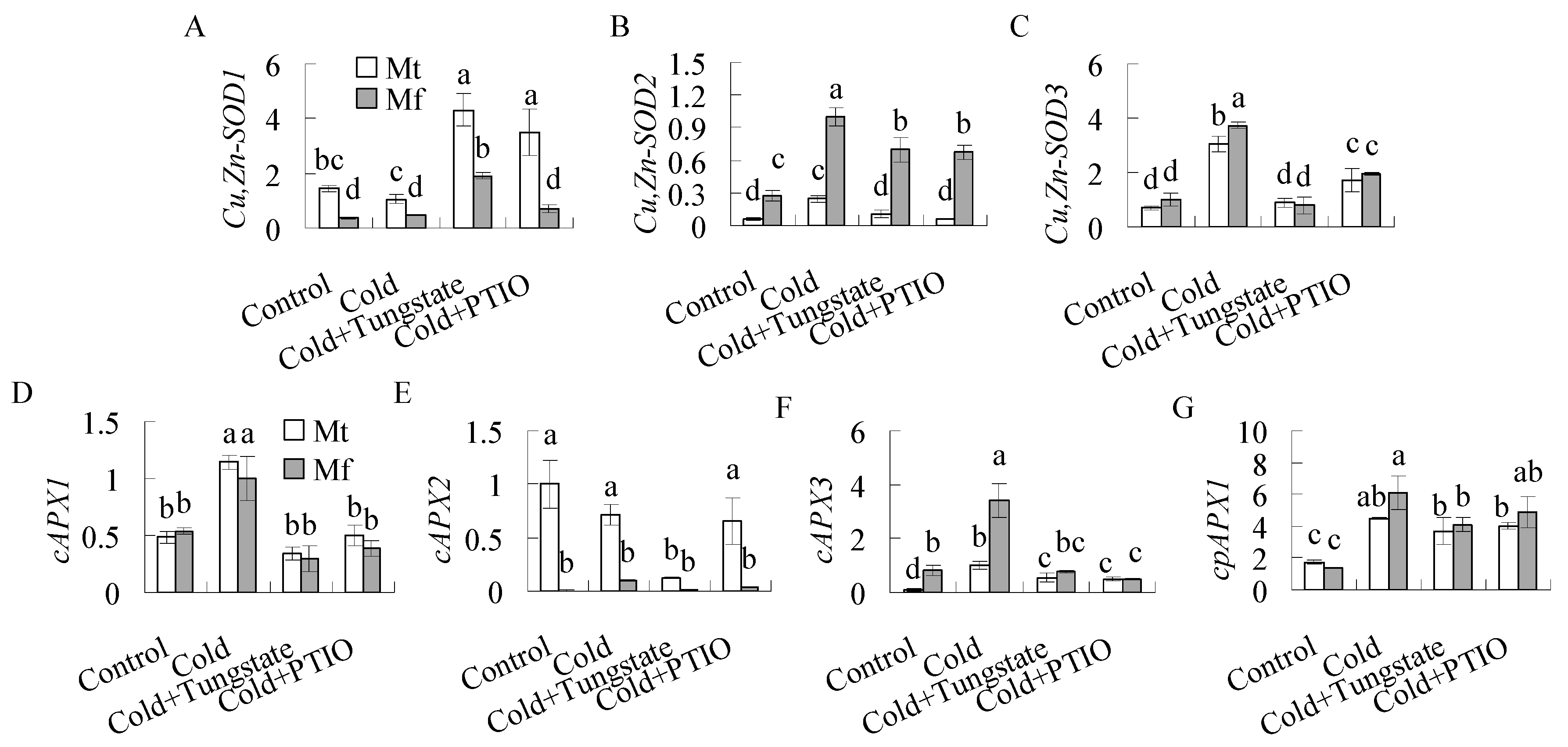
2.4. Antioxidant Enzyme Activities were Induced by Cold and NO
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Treatments
4.2. Isolation of RNA and Real-Time Quantitative PCR (qPCR) Analysis
4.3. Evaluation of Cold Tolerance
4.4. Detection of NO in Leaves
4.5. Determination of NR, SOD, CAT, and APX Activities
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Hua, J. From freezing to scorching, transcriptional responses to temperature variations in plants. Curr. Opin. Plant Biol. 2009, 12, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Ding, Y.; Yang, S. Molecular regulation of CBF signaling in cold acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van den Ende, W.; Cuypers, A. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.J.; Ort, D.R. Impact of chilling temperatures on photosynthesis in warm climate plants. Trends Plant Sci. 2001, 6, 36–42. [Google Scholar] [CrossRef]
- Puyaubert, J.; Baudouin, E. New clues for a cold case: Nitric oxide response to low temperature. Plant Cell Environ. 2014, 37, 2623–2630. [Google Scholar] [CrossRef]
- Cantrel, C.; Vazquez, T.; Puyaubert, J.; Reze, N.; Lesch, M.; Kaiser, W.M.; Dutilleul, C.; Guillas, I.; Zachowski, A.; Baudouin, E. Nitric oxide participates in cold-responsive phosphosphingolipid formation and gene expression in Arabidopsis thaliana. New Phytol. 2011, 189, 415–427. [Google Scholar] [CrossRef]
- Corpas, F.J.; Chaki, M.; Fernandez-Ocana, A.; Valderrama, R.; Palma, J.M.; Carreras, A.; Begara-Morales, J.C.; Airaki, M.; del Rio, L.A.; Barroso, J.B. Metabolism of reactive nitrogen species in pea plants under abiotic stress conditions. Plant Cell Physiol. 2008, 49, 1711–1722. [Google Scholar] [CrossRef]
- Ziogas, V.; Tanou, G.; Filippou, P.; Diamantidis, G.; Vasilakakis, M.; Fotopoulos, V.; Molassiotis, A. Nitrosative responses in citrus plants exposed to six abiotic stress conditions. Plant Physiol. Biochem. 2013, 68, 118–126. [Google Scholar] [CrossRef]
- Sehrawat, A.; Abat, J.K.; Deswal, R. RuBisCO depletion improved proteome coverage of cold responsive S-nitrosylated targets in Brassica juncea. Front. Plant Sci. 2013, 4, 342. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Esim, N.; Atici, O. Nitric oxide improves chilling tolerance of maize by affecting apoplastic antioxidative enzymes in leaves. Plant Growth Regul. 2014, 72, 29–38. [Google Scholar] [CrossRef]
- Mur, L.A.; Mandon, J.; Persijn, S.; Cristescu, S.M.; Moshkov, I.E.; Novikova, G.V.; Hall, M.A.; Harren, F.J.; Hebelstrup, K.H.; Gupta, K.J. Nitric oxide in plants: An assessment of the current state of knowledge. AoB Plants 2013, 5, pls052. [Google Scholar] [CrossRef] [PubMed]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fermandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Costa-Broseta, A.; Perea-Resa, C.; Castillo, M.C.; Ruiz, M.F.; Salinas, J.; Leon, J. Nitric oxide controls constitutive freezing tolerance in Arabidopsis by attenuating the levels of osmoprotectants, stress-related hormones and anthocyanins. Sci. Rep. 2018, 8, 9268. [Google Scholar] [CrossRef]
- Lu, S.; Zhuo, C.; Wang, X.; Guo, Z. Nitrate reductase (NR)-dependent NO production mediates ABA- and H2O2-induced antioxidant enzymes. Plant Physiol. Biochem. 2014, 74, 9–15. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, Ã.; Ocanã-Calahorro, F.; Mariscal, V.; Carreras, A.; Barroso, J.B.; Galvã, A.; Fernãndez, E. A dual system formed by the ARC and NR molybdoenzymes mediates nitrite-dependent NO production in Chlamydomonas. Plant Cell Environ. 2016, 39, 2097–2107. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
- Wilson, I.D.; Ribeiro, D.M.; Bright, J.; Confraria, A.; Harrison, J.; Barros, R.S.; Desikan, R.; Neill, S.J.; Hancock, J.T. Role of nitric oxide in regulating stomatal apertures. Plant Signal. Behav. 2009, 4, 467–469. [Google Scholar] [CrossRef] [Green Version]
- Horchani, F.; Prevot, M.; Boscari, A.; Evangelisti, E.; Meilhoc, E.; Bruand, C.; Raymond, P.; Boncompagni, E.; Aschi-Smiti, S.; Puppo, A.; et al. Both plant and bacterial nitrate reductases contribute to nitric oxide production in Medicago truncatula nitrogen-fixing nodules. Plant Physiol. 2011, 155, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Riday, H.; Brummer, E.C. Forage yield heterosis in alfalfa. Crop Sci. 2002, 42, 716–723. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Cheng, H.; Stockinger, E.J. Comparative genomic sequence and expression analyses of Medicago truncatula and alfalfa subspecies falcata COLD-ACCLIMATION-SPECIFIC genes. Plant Physiol. 2008, 146, 1242–1254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Zhao, M.G.; Tian, Q.Y.; Zhang, W.H. Comparative studies on tolerance of Medicago truncatula and Medicago falcata to freezing. Planta 2011, 234, 445–457. [Google Scholar] [CrossRef]
- Tan, J.; Wang, C.; Xiang, B.; Han, R.; Guo, Z. Hydrogen peroxide and nitric oxide mediated cold- and dehydration-induced myo-inositol phosphate synthase that confers multiple resistances to abiotic stresses. Plant Cell Environ. 2013, 36, 288–299. [Google Scholar] [CrossRef]
- Tan, J.; Zhuo, C.; Guo, Z. Nitric oxide mediates cold- and dehydration-induced expression of a novel MfHyPRP that confers tolerance to abiotic stress. Physiol. Plant. 2013, 149, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, C.; Wang, T.; Lu, S.; Zhao, Y.; Li, X.; Guo, Z. A cold responsive galactinol synthase gene from Medicago falcata (MfGolS1) is induced by myo-inositol and confers multiple tolerances to abiotic stresses. Physiol. Plant. 2013, 149, 67–78. [Google Scholar] [CrossRef]
- Sambe, M.A.; He, X.; Tu, Q.; Guo, Z. A cold-induced myo-inositol transporter-like gene (MfINT-like) confers tolerance to multiple abiotic stresses in transgenic tobacco plants. Physiol. Plant. 2015, 153, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Tan, J.; Zhuo, C.; Wang, C.; Xiang, B.; Wang, Z. Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. 2014, 12, 601–612. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Sambe, M.A.; Zhuo, C.; Tu, Q.; Guo, Z. A temperature induced lipocalin gene from Medicago falcata (MfTIL1) confers tolerance to cold and oxidative stress. Plant Mol. Biol. 2015, 87, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Su, W.; Li, H.; Guo, Z. Abscisic acid improves drought tolerance of triploid bermudagrass and involves H2O2- and NO-induced antioxidant enzyme activities. Plant Physiol. Biochem. 2009, 47, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Alberto Ferrarini, A.; De Stefano, M.; Baudouin, E.; Pucciariello, E.; Polverari, A.; Alain Puppo, A.; Delledonne, M. Expression of Medicago truncatula genes responsive to nitric oxide in pathogenic and symbiotic conditions. Mol. Plant Microbe Interact. 2008, 21, 781–790. [Google Scholar] [CrossRef]
- Huang, J.; Wei, H.; Li, L.; Yu, S. Transcriptome analysis of nitric oxideresponsive genes in upland cotton (Gossypium hirsutum). PLoS ONE 2018, 13, e0192367. [Google Scholar] [CrossRef]
- Chen, J.; Guo, Z.; Fang, J.; Lu, S. Physiological responses of a centipedegrass mutant to chilling stress. Agron. J. 2013, 105, 1814–1820. [Google Scholar] [CrossRef]
- Lu, S.; Wang, X.; Guo, Z. Differential responses to chilling in Stylosanthes guianensis (Aublet) Sw. and its mutants. Agron. J. 2013, 105, 377. [Google Scholar] [CrossRef]
- Zhou, B.; Guo, Z.; Xing, J.; Huang, B. Nitric oxide is involved in abscisic acid-induced antioxidant activities in Stylosanthes guianensis. J. Exp. Bot. 2005, 56, 3223–3228. [Google Scholar] [CrossRef]
- Zhang, A.; Jiang, M.; Zhang, J.; Ding, H.; Xu, S.; Hu, X.; Tan, M. Nitric oxide induced by hydrogen peroxide mediates abscisic acid-induced activation of the mitogen-activated protein kinase cascade involved in antioxidant defense in maize leaves. New Phytol. 2007, 175, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, J.; Guo, Z.; Lu, S.; He, S.; Shu, W.; Zhou, B. Increased abscisic acid levels in transgenic tobacco over-expressing 9 cis-epoxycarotenoid dioxygenase influence H2O2 and NO production and antioxidant defences. Plant Cell Environ. 2009, 32, 509–519. [Google Scholar] [CrossRef]
- Santisree, P.; Bhatnagar-Mathur, P.; Sharma, K.K. Molecular insights into the functional role of nitric oxide (NO) as a signal for plant responses in chickpea. Funct. Plant Biol. 2018, 45, 267–283. [Google Scholar] [CrossRef]
- Wilkins, K.; Bancroft, J.; Bosch, M.; Ings, J.; Smirnoff, N.; Franklin-Tong, V. ROS and NO mediate actin reorganization and programmed cell death in the self-incompatibility response of Papaver. Plant Physiol. 2011, 156, 404–416. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene name | Accession No. | Primer name | Sequence |
|---|---|---|---|
| MtNIA1/MfNIA1 | MTR3g073180 | ZG2619 | TGGCTCAACCTTGGATAT |
| ZG2620 | TGCTTACCGTGAACCATA | ||
| MtNIA2/MfNIA2 | MTR5g059820 | ZG2621 | GAGGATTGTTGTTACTACTG |
| ZG2622 | CACGGAGTTTATGTTCA | ||
| MtCu,Zn-SOD1/MfCu,Zn-SOD1 | MTR4g057240 | ZG6010 ZG6011 | GCTTAATGTCCTAGATGAGT AGCAGGCAAGAAGTATTG |
| MtCu,Zn-SOD2/MfCu,Zn-SOD2 | MTR6g029200 | ZG6012 ZG6013 | ACTCCAGTCATCTGTTTAG CCATTACGCATAGAACAAC |
| MtCu,Zn-SOD3/MfCu,Zn-SOD3 | MTR7g114240 | ZG6014 ZG6015 | TTCCATATCCATGCCTTG CGTGCTCCTTACCATTAG |
| MtcAPX1/MfcAPX1 | MTR3g107060 | ZG6016 | GGAGGTCCTACTATCACA |
| ZG6017 | CCAGAAGCATCAAGAGTT | ||
| MtcAPX2/MfcAPX2 | MTR4g061140 | ZG6020 | CCTGATGGAGTGTTCAAT |
| ZG6021 | ACCTTGCCTACTTCATATC | ||
| MtcAPX3/MfcAPX3 | MTR5g022510 | ZG6022 | GATAAGGCTCTAGTTGATGA |
| ZG6023 | TGAGGCAATAACCATTCC | ||
| MtcpAPX1/MfcpAPX1 | MTR3g088160 | ZG6024 | TGGATTCTGAACAGTGAAC |
| ZG6025 | CGACCTCCTCTATCTTGA | ||
| MtActin/MfActin | MTR3g095530 | ZG1613 | ATTCACGAGACCACCTAC |
| ZG1614 | GAGCCACAACCTTAATCTTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Li, S.; Zhao, P.; Guo, Z.; Lu, S. Comparative Physiological Analysis Reveals the Role of NR-Derived Nitric Oxide in the Cold Tolerance of Forage Legumes. Int. J. Mol. Sci. 2019, 20, 1368. https://doi.org/10.3390/ijms20061368
Zhang P, Li S, Zhao P, Guo Z, Lu S. Comparative Physiological Analysis Reveals the Role of NR-Derived Nitric Oxide in the Cold Tolerance of Forage Legumes. International Journal of Molecular Sciences. 2019; 20(6):1368. https://doi.org/10.3390/ijms20061368
Chicago/Turabian StyleZhang, Peipei, Shuangshuang Li, Pengcheng Zhao, Zhenfei Guo, and Shaoyun Lu. 2019. "Comparative Physiological Analysis Reveals the Role of NR-Derived Nitric Oxide in the Cold Tolerance of Forage Legumes" International Journal of Molecular Sciences 20, no. 6: 1368. https://doi.org/10.3390/ijms20061368