In Silico Molecular Docking and In Vivo Validation with Caenorhabditis elegans to Discover Molecular Initiating Events in Adverse Outcome Pathway Framework: Case Study on Endocrine-Disrupting Chemicals with Estrogen and Androgen Receptors


Abstract
:1. Introduction
2. Results and Discussion
2.1. Preparation of Ligands from Tox21 Assays
2.2. Homology Modeling of C. elegans Receptors
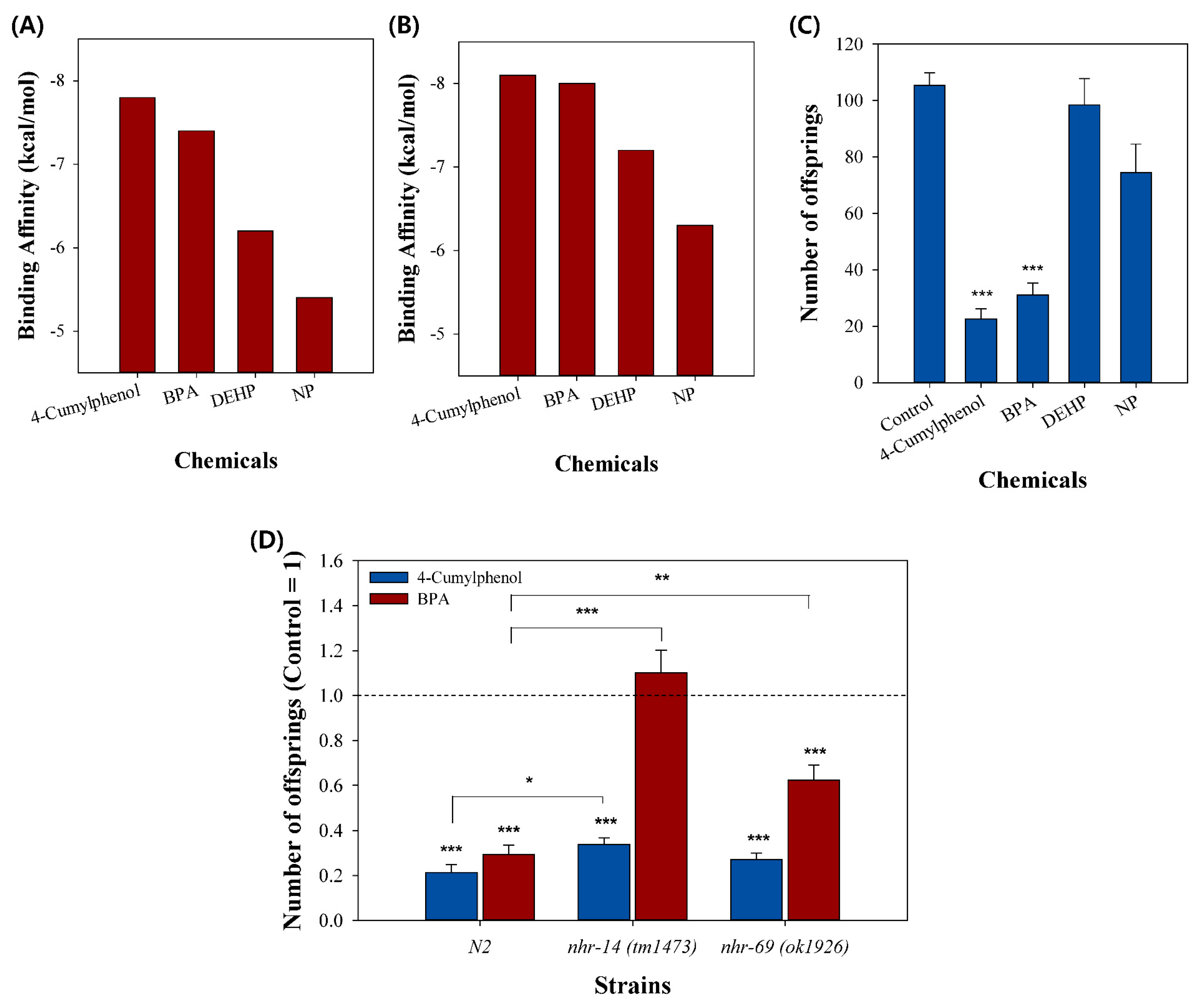
2.3. Docking Simulation with Human Receptor
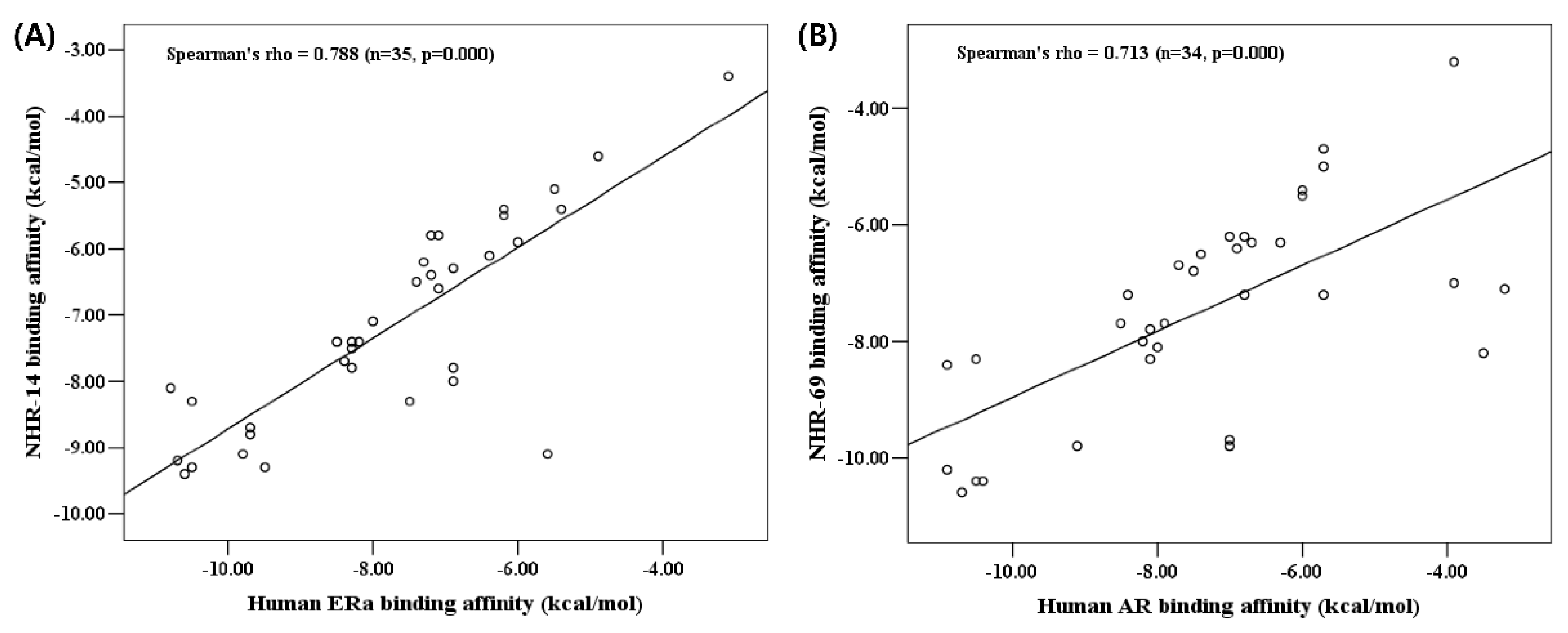
2.4. Docking Simulation with C. elegans Receptors
2.5. Experimental Validation on NHR-14 and NHR-69
3. Materials and Methods
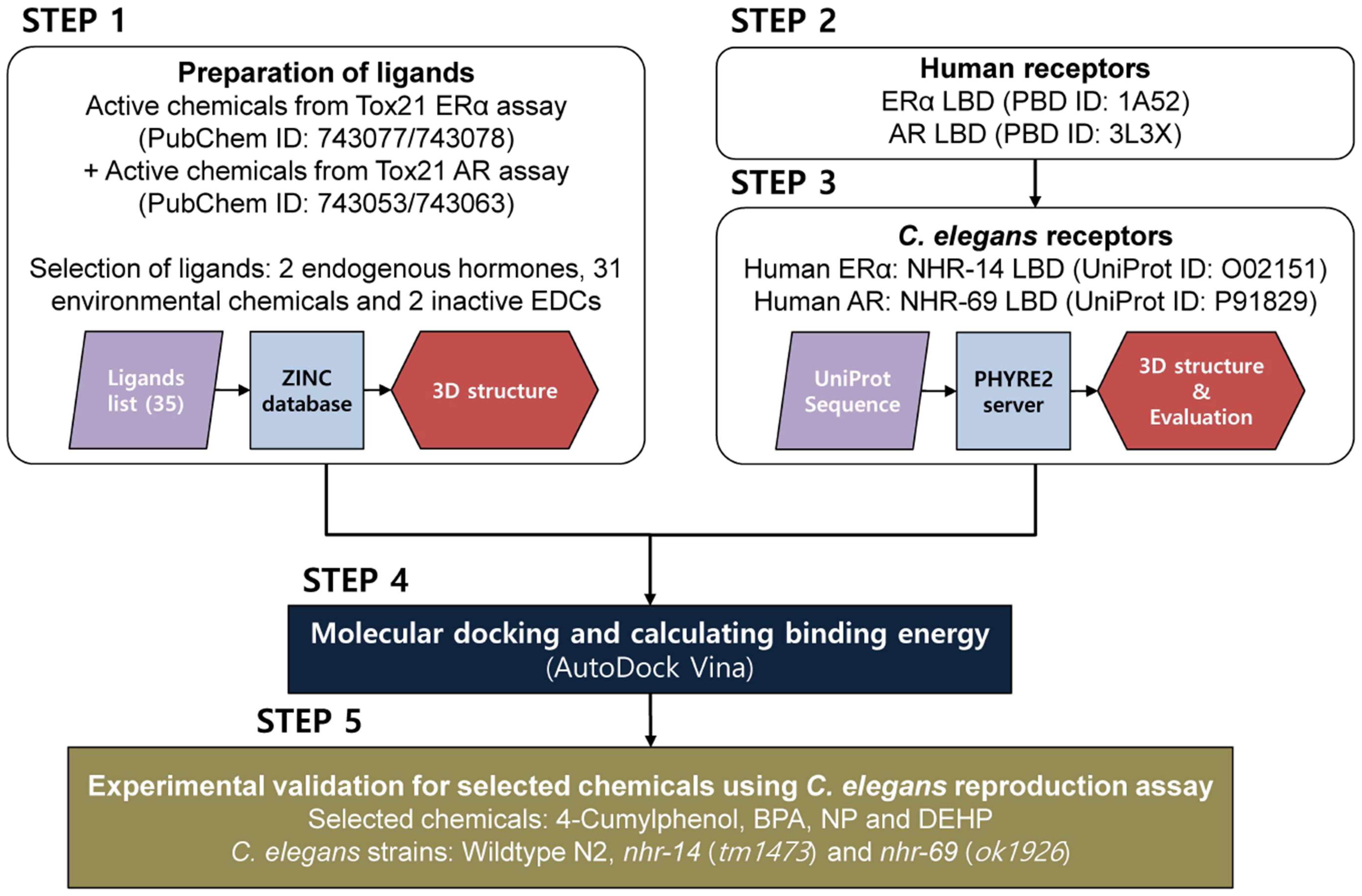
3.1. Study Design
- Step 1: Active chemicals from Tox21 ERα agonists/antagonists (PubChem ID: 743077/743078) and AR agonists/antagonists (PubChem ID: 743053/743063) assays were used as ligands for docking analysis.
- Step 2: Human ERα and AR LBDs were prepared from the Protein Data Bank (PDB).
- Step 3: The LBD sequences of C. elegans nuclear hormone receptor (NHR) known to homologous to human ERα and AR were collected from UniProt. 3D structures of each protein were built using homology modeling by PHYRE2 server, and the binding grids of each structure were defined.
- Step 4: The optimized ligands were then docked into the binding grid of each 3D human ERα and AR and C. elegans NHR structures using AutoDock Vina. The order of the calculated binding free energy was compared between human and C. elegans receptors.
- Step 5: Experimental validation was conducted using C. elegans reproduction assay on the selected chemicals.
3.2. Preparation of Ligands
3.3. Preparation of Human Receptors
3.4. Homology Modeling of C. elegans Receptors
3.5. Docking Simulations
3.6. C. elegans Reproduction Assay
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AOP | Adverse Outcome Pathway |
| AR | Androgen Receptor |
| BPA | Bisphenol A |
| DEHP | Bis(2-ethylhexyl)phthalate |
| EDCs | Endocrine-disrupting Chemicals |
| ER | Estrogen Receptor |
| LBD | Ligand Binding Domain |
| MIE | Molecular Initiating Event |
| NHR | Nuclear Hormone Receptor |
| NP | 4-Nonylphenol |
References
- Parthasarathi, R. In Silico Approaches for Predictive Toxicology. In Vitro Toxicol. 2018, 91–109. [Google Scholar]
- Raies, A.B.; Bajic, V.B. In silico toxicology: Computational methods for the prediction of chemical toxicity. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2016, 6, 147–172. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lin, Z.; Chen, R.; Wang, T.; Liu, S.; Yao, Z.; Yin, D. Using molecular docking to compare toxicity of reactive chemicals to freshwater and marine luminous bacteria. Mol. Inform. 2012, 31, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.R.; Goldsmith, M.R.; Little, S.B.; Pasquinelli, M.A. Computational Molecular Modeling for evaluating the toxicity of environmental chemicals: Prioritizing bioassay requirements. Environ. Health Perspect. 2008, 116, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Al Sharif, M.; Tsakovska, I.; Pajeva, I.; Alov, P.; Fioravanzo, E.; Bassan, A.; Kovarich, S.; Yang, C.; Mostrag-Szlichtyng, A.; Vitcheva, V.; et al. The application of molecular modelling in the safety assessment of chemicals: A case study on ligand-dependent PPARγ dysregulation. Toxicology 2017, 392, 140–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. Guidance Document on Developing and Assessing Adverse Outcome Pathways; OECD: Paris, France, 2013; Volume 184. [Google Scholar]
- Le Maire, A.; Bourguet, W.; Balaguer, P. A structural view of nuclear hormone receptor: Endocrine disruptor interactions. Cell. Mol. Life Sci. 2010, 67, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Swedenborg, E.; Rüegg, J.; Mäkelä, S.; Pongratz, I. Endocrine disruptive chemicals: Mechanisms of action and involvement in metabolic disorders. J. Mol. Endocrinol. 2009, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.E.H.; Goodman, J.M.; Gutsell, S.; Russell, P.J. A History of the Molecular Initiating Event. Chem. Res. Toxicol. 2016, 29, 2060–2070. [Google Scholar] [CrossRef] [PubMed]
- Avila, D.S.; Adams, M.R.; Chakraborty, S.; Aschner, M. Caenorhabditis elegans as a model to assess reproductive and developmental toxicity. Reprod. Dev. Toxicol. 2011, 193–205. [Google Scholar]
- Hunt, P.R. The C. elegans model in toxicity testing. J. Appl. Toxicol. 2017, 37, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Tsyusko, O.V.; Unrine, J.M.; Chatterjee, N.; Ahn, J.M.; Yang, X.; Thornton, B.L.; Ryde, I.T.; Starnes, D.; Meyer, J.N. A micro-sized model for the in vivo study of nanoparticle toxicity: What has Caenorhabditis elegans taught us? Environ. Chem. 2014, 11, 227–246. [Google Scholar] [CrossRef]
- Williams, P.L.; Dusenbery, D.B. Using the Nematode Caenorhabditis Elegans To Predict Mammalian Acute Lethality To Metallic Salts. Toxicol. Ind. Health 1988, 4, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.D.; Anderson, G.L.; Williams, P.L. The nematode Caenorhabditis elegans as a model of organophosphate-induced mammalian neurotoxicity. Toxicol. Appl. Pharmacol. 2004, 194, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Boyd, W.A.; McBride, S.J.; Rice, J.R.; Snyder, D.W.; Freedman, J.H. A high-throughput method for assessing chemical toxicity using a Caenorhabditis elegans reproduction assay. Toxicol. Appl. Pharmacol. 2010, 245, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.H.; Liu, A.L.; Chen, Y.Y.; Zhang, L.; Zhang, H.J.; Jin, B.X.; Lu, W.H.; Li, X.Y.; Lu, W.Q. DNA damage and oxidative stress in human liver cell L-02 caused by surface water extracts during drinking water treatment in a waterworks in China. Environ. Mol. Mutagen. 2014, 55, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Ferguson; Boyer, M.S.; Sprando, R.L. A method for ranking compounds based on their relative toxicity using neural networking, C. elegans, axenic liquid culture, and the COPAS parameters TOF and EXT. Open Access Bioinform. 2010, 2010, 139–144. [Google Scholar] [CrossRef]
- Hunt, P.R.; Olejnik, N.; Sprando, R.L. Toxicity ranking of heavy metals with screening method using adult Caenorhabditis elegans and propidium iodide replicates toxicity ranking in rat. Food Chem. Toxicol. 2012, 50, 3280–3290. [Google Scholar] [CrossRef] [PubMed]
- Vaux, D.L.; Weissman, I.L.; Kim, S.K. Prevention of programmed cell death in Caenorhabditis elegans by human bcl-2. Science 1992, 258, 1955–1957. [Google Scholar] [CrossRef] [PubMed]
- Ogg, S.; Paradis, S.; Gottlieb, S.; Patterson, G.I.; Lee, L.; Tissenbaum, H.A.; Ruvkun, G. The fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 1997, 389, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Mimoto, A.; Fujii, M.; Usami, M.; Shimamura, M.; Hirabayashi, N.; Kaneko, T.; Sasagawa, N.; Ishiura, S. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2007, 364, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Gámez-del-Estal, M.; Contreras, I.; Prieto-Perez, R.M.; Ruiz-Rubio, M. Epigenetic effect of testosterone in the behavior of C. elegans. A clue to explain androgen-dependent autistic traits? Front. Cell. Neurosci. 2014, 8, 69. [Google Scholar] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Wass, M.N.; Kelley, L.A.; Sternberg, M.J.E. 3DLigandSite: Predicting ligand-binding sites using similar structures. Nucleic Acids Res. 2010, 38, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hong, S.; Liu, X.; Kim, C.; Jung, D.; Yim, U.H.; Shim, W.J.; Khim, J.S.; Giesy, J.P.; Choi, K. Endocrine disrupting potential of PAHs and their alkylated analogues associated with oil spills. Environ. Sci. Process. Impacts 2017, 19, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dong, S.; Wang, H.; Tao, S.; Kiyama, R. Biological impact of environmental polycyclic aromatic hydrocarbons (ePAHs) as endocrine disruptors. Environ. Pollut. 2016, 213, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Bonefeld-Jørgensen, E.C.; Long, M.; Hofmeister, M.V.; Vinggaard, A.M. Endocrine-disrupting potential of Bisphenol A, Bisphenol A dimethacrylate, 4-n-nonylphenol, and 4-n-octylphenol in vitro: New data and a brief review. Environ. Health Perspect. 2007, 115, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Meerts, I.A.; Letcher, R.J.; Hoving, S.; Marsh, G.; Bergman, A.; Lemmen, J.G.; Van Der Burg, B.; Brouwer, A. In vitro estrogenicity of polybrominated diphenyl ethers, hydroxylated PBDEs, and polybrominated bisphenol A compounds. Environ. Health Perspect. 2001, 109, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Shanle, E.K.; Xu, W. Endocrine disrupting chemicals targeting estrogen receptor signaling: Identification and mechanisms of action. Chem. Res. Toxicol. 2011, 24, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Kolšek, K.; Mavri, J.; Sollner Dolenc, M.; Gobec, S.; Turk, S. Endocrine disruptome—An open source prediction tool for assessing endocrine disruption potential through nuclear receptor binding. J. Chem. Inf. Model. 2014, 54, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Grignard, E.; Lapenna, S.; Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicol. In Vitro 2012, 26, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Montes-Grajales, D.; Olivero-Verbel, J. Computer-aided identification of novel protein targets of bisphenol A. Toxicol. Lett. 2013, 222, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.-W.; Cho, J.; Ahnn, J.; Song, H.-O. Novel Findings of Anti-Filarial Drug Target and Structure-Based Virtual Screening for Drug Discovery. Int. J. Mol. Sci. 2018, 19, 3579. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Tong, W.; Branham, W.S.; Moland, C.L.; Dial, S.L.; Hong, H.; Xie, Q.; Perkins, R.; Owens, W.; Sheehan, D.M. Study of 202 Natural, Synthetic, and Environmental Chemicals for Binding to the Androgen Receptor. Chem. Res. Toxicol. 2003, 16, 1338–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, J.P.; Huang, W.; Chapman, L.M.; Chua, S.; Moore, D.D.; Baldwin, W.S. The environmental estrogen, nonylphenol, activates the constitutive androstane receptor. Toxicol. Sci. 2007, 98, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Nishiyama, N.; Sugiyama, S.I.; Yamauchi, K. The effect of endocrine disrupting chemicals on thyroid hormone binding to Japanese quail transthyretin and thyroid hormone receptor. Gen. Comp. Endocrinol. 2003, 134, 36–43. [Google Scholar] [CrossRef]
- Janicki, T.; Krupiński, M.; Długoński, J. Degradation and toxicity reduction of the endocrine disruptors nonylphenol, 4-tert-octylphenol and 4-cumylphenol by the non-ligninolytic fungus Umbelopsis isabellina. Bioresour. Technol. 2016, 200, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Kerdivel, G.; Habauzit, D.; Pakdel, F. Assessment and molecular actions of endocrine-disrupting chemicals that interfere with estrogen receptor pathways. Int. J. Endocrinol. 2013, 2013, 501851. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhao, L.; Wei, L.; Li, L. DEHP reduces thyroid hormones via interacting with hormone synthesis-related proteins, deiodinases, transthyretin, receptors, and hepatic enzymes in rats. Environ. Sci. Pollut. Res. 2015, 22, 12711–12719. [Google Scholar] [CrossRef] [PubMed]
- Rosenmai, A.K.; Dybdahl, M.; Pedersen, M.; van Vugt-Lussenburg, B.M.A.; Wedebye, E.B.; Taxvig, C.; Vinggaard, A.M. Are structural analogues to bisphenol a safe alternatives? Toxicol. Sci. 2014, 139, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Zhu, Z. Simultaneous identification and quantification of 4-cumylphenol, 2,4-bis-(dimethylbenzyl)phenol and bisphenol A in prawn Macrobrachium rosenbergii. Chemosphere 2014, 107, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Sterling, T.; Mysinger, M.M.; Bolstad, E.S.; Coleman, R.G. ZINC: A free tool to discover chemistry for biology. J. Chem. Inf. Model. 2012, 52, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The Protein Data Bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Nowotny, J.; Cao, R.; Cheng, J. 3Drefine: An interactive web server for efficient protein structure refinement. Nucleic Acids Res. 2016, 44, W406–W409. [Google Scholar] [CrossRef] [PubMed]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B.; de Bakker, P.I.W.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Cα geometry: ϕ,ψ and Cβ deviation. Proteins Struct. Funct. Bioinforma. 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Chemical | Estrogen Receptor Alpha | Androgen Receptor | |||
|---|---|---|---|---|---|---|
| Name | CAS No. | ERα (kcal/mol) | NHR-14 (kcal/mol) | AR (kcal/mol) | NHR-69 (kcal/mol) | |
| 1 | 17beta-Estradiol | 50-28-2 | −10.5 | −8.3 | −10.9 | −8.4 |
| 2 | Testosterone | 58-22-0 | −10.8 | −8.1 | −10.5 | −8.3 |
| 3 | 1,2-Benzenedicarboxaldehyde | 643-79-8 | −5.4 | −5.4 | −5.7 | −5.0 |
| 4 | 1,3-Diiminobenz[f]isoindoline | 65558-69-2 | −8.3 | −7.5 | −8.4 | −7.2 |
| 5 | 1,6-Hexanediol diacrylate | 13048-33-4 | −5.5 | −5.1 | −6.0 | −5.4 |
| 6 | 10-Chloro-9-anthraldehyde | 10527-16-9 | −8.0 | −7.1 | −7.9 | −7.7 |
| 7 | 2,2′-Methylenebis(4-methyl-6-tert-butylphenol) | 119-47-1 | −6.9 | −8.0 | −3.5 | −8.2 |
| 8 | 2,4-Bis(1-methyl-1-phenylethyl)phenol | 2772-45-4 | −9.7 * | −8.7 | −7.0 | −9.8 * |
| 9 | 2-Aminoanthracene | 613-13-8 | −8.5 | −7.4 | −8.5 | −7.7 |
| 10 | 4,4′-Thiobis(6-tert-butyl-m-cresol) | 96-69-5 | −6.9 | −7.8 | −3.2 | −7.1 |
| 11 | 4,6-Di-tert-butyl-m-cresol | 497-39-2 | −6.9 | −6.3 | −6.8 | −7.2 |
| 12 | 4-Cumylphenol | 599-64-4 | −8.3 | −7.8 | −8.0 | −8.1 |
| 13 | 4-Nitrosodiphenylamine | 156-10-5 | −7.1 | −6.6 | −7.5 | −6.8 |
| 14 | 4-Nonylphenol | 104-40-5 | −6.2 | −5.4 | −6.3 | −6.3 |
| 15 | 7-(Dimethylamino)-4-methylcoumarin | 87-01-4 | −7.2 | −6.4 | −7.4 | −6.5 |
| 16 | 7-Diethylamino-4-methylcoumarin | 91-44-1 | −7.4 | −6.5 | −7.7 | −6.7 |
| 17 | 7-Methylbenzo[a]pyrene | 63041-77-0 | −10.6 * | −9.4 * | −10.5 * | −10.4 * |
| 18 | 9,10-Dihydrobenzo[a]pyren-7(8H)-one | 3331-46-2 | −10.5 * | −9.3 * | −10.4 * | −10.4 * |
| 19 | 9-Bromoanthracene | 1564-64-3 | −8.2 | −7.4 | −8.1 | −7.8 |
| 20 | 9-Cyanoanthracene | 1210-12-4 | −8.4 | −7.7 | −8.1 | −8.3 |
| 21 | alpha-Terthiophene | 1081-34-1 | −6.4 | −6.1 | −6.8 | −6.2 |
| 22 | Benzo[a]pyrene | 50-32-8 | −10.7 * | −9.2 * | −10.7 * | −10.6 * |
| 23 | Benzo[b]fluoranthene | 205-99-2 | −9.5 | −9.3 * | −9.1 * | −9.8 * |
| 24 | Benzo[e]pyrene | 192-97-2 | −9.7 * | −8.8 | −7.0 | −9.7 |
| 25 | Benzo[k]fluoranthene | 207-08-9 | −9.8 * | −9.1 * | −10.9 * | −10.2 * |
| 26 | Bis(2-Ethylhexyl)phthalate (DEHP) | 117-81-7 | −7.3 | −6.2 | −5.7 | −7.2 |
| 27 | Bisphenol A | 80-05-07 | −8.3 | −7.4 | −8.2 | −8.0 |
| 28 | Chlorothalonil | 1897-45-6 | −6.0 | −5.9 | −6.7 | −6.3 |
| 29 | Crystal Violet lactone | 1552-42-7 | −5.6 | −9.1 * | NA | −7.3 |
| 30 | Dodecyl gallate | 1166-52-5 | −7.2 | −5.8 | −6.9 | −6.4 |
| 31 | Ethylene acrylate | 2274-11-5 | −4.9 | −4.6 | −5.7 | −4.7 |
| 32 | Fluazinam | 79622-59-6 | −7.5 | −8.3 | −3.9 | −7.0 |
| 33 | Octyl gallate | 1034-01-1 | −7.1 | −5.8 | −7.0 | −6.2 |
| 34 | Tribromoacetaldehyde | 115-17-3 | −3.1 | −3.4 | −3.9 | −3.2 |
| 35 | Trimethylolpropane triacrylate | 15625-89-5 | −6.2 | −5.5 | −6.0 | −5.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.; Kim, H.; Choi, J. In Silico Molecular Docking and In Vivo Validation with Caenorhabditis elegans to Discover Molecular Initiating Events in Adverse Outcome Pathway Framework: Case Study on Endocrine-Disrupting Chemicals with Estrogen and Androgen Receptors. Int. J. Mol. Sci. 2019, 20, 1209. https://doi.org/10.3390/ijms20051209
Jeong J, Kim H, Choi J. In Silico Molecular Docking and In Vivo Validation with Caenorhabditis elegans to Discover Molecular Initiating Events in Adverse Outcome Pathway Framework: Case Study on Endocrine-Disrupting Chemicals with Estrogen and Androgen Receptors. International Journal of Molecular Sciences. 2019; 20(5):1209. https://doi.org/10.3390/ijms20051209
Chicago/Turabian StyleJeong, Jaeseong, Hunbeen Kim, and Jinhee Choi. 2019. "In Silico Molecular Docking and In Vivo Validation with Caenorhabditis elegans to Discover Molecular Initiating Events in Adverse Outcome Pathway Framework: Case Study on Endocrine-Disrupting Chemicals with Estrogen and Androgen Receptors" International Journal of Molecular Sciences 20, no. 5: 1209. https://doi.org/10.3390/ijms20051209