HDAC Inhibitors Induce BDNF Expression and Promote Neurite Outgrowth in Human Neural Progenitor Cells-Derived Neurons


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Primary Screenings for Enhancers of BDNF mRNA Expression
2.2. HNPCs and Fibroblasts Shared Common Hits
2.3. HDACis Increased BDNF in Secondary Assays
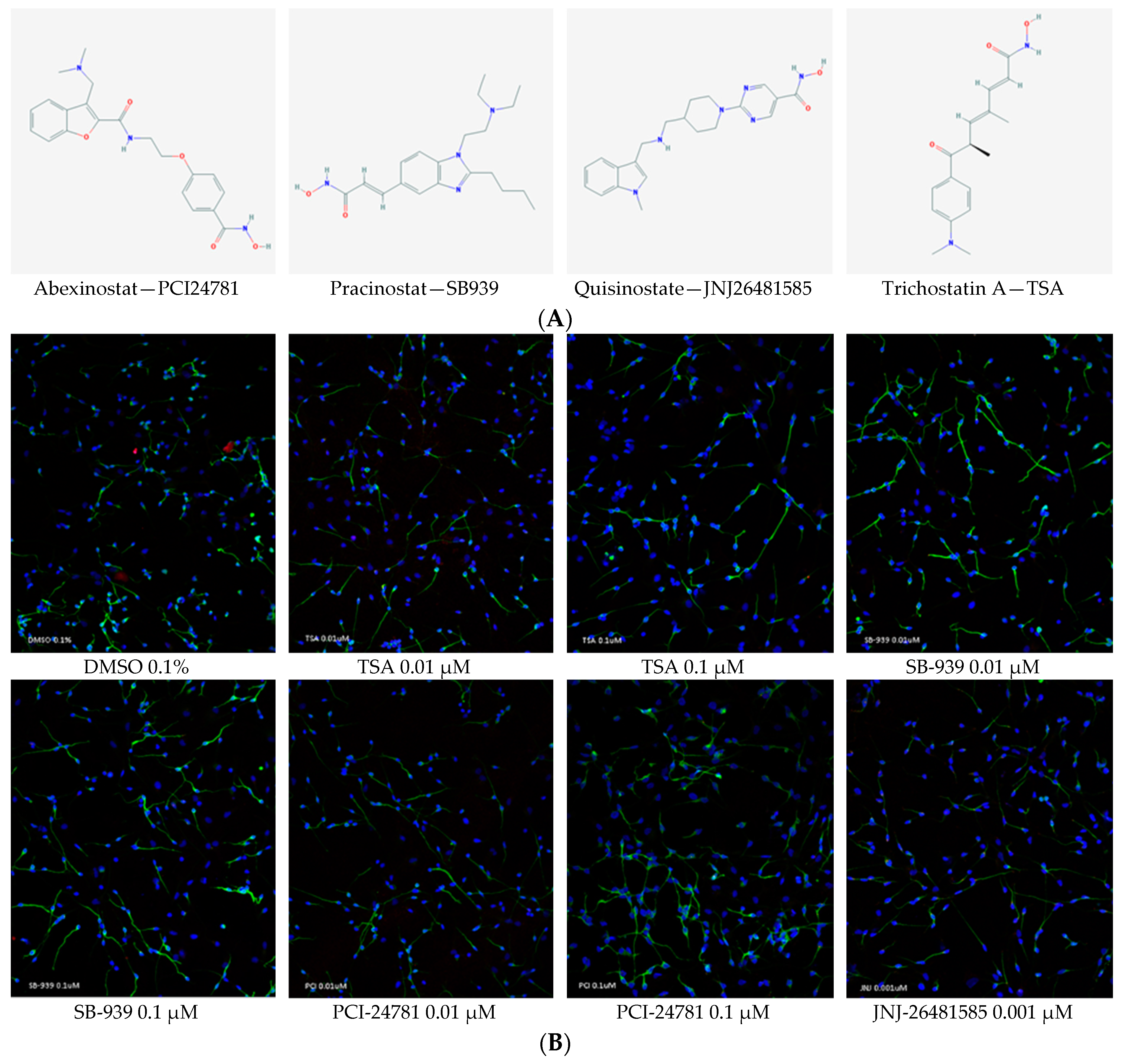
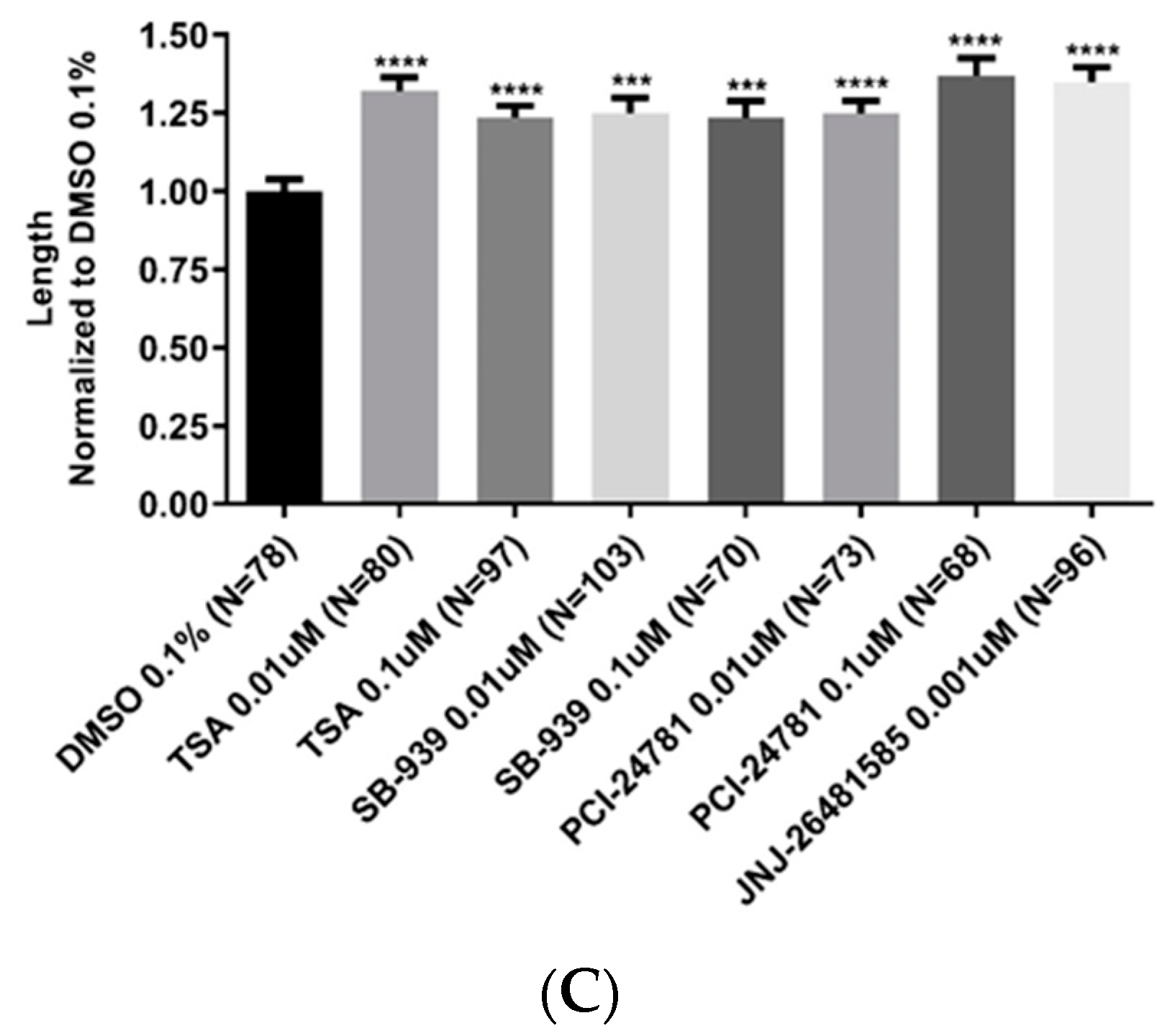
2.4. Neurite Length Was Significantly Enhanced upon Treatment with Hydroxamate-Based HDACis
2.5. The Hydroxamate-Based HDACis Showed No Toxicity at the Concentrations Used
3. Discussion
4. Materials and Methods
4.1. High-Throughput Screening Assay Using an In-House Epigenetics Compound Library
4.2. Neurite Outgrowth Experiment
4.2.1. Isolation and Culture of Human Neural Progenitor Cells (HNPCs)
4.2.2. Differentiation and Treatment of HNPCs
4.2.3. Immunocytochemistry (ICC)
4.2.4. Image Acquisition and Neurite Outgrowth Quantification
4.2.5. RNA Extraction, cDNA Synthesis, and Real-Time PCR
4.2.6. Cell Culture Drug Treatment
4.2.7. Cell Viability Assay
4.2.8. ELISA Experiments
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BDNF | Brain-derived Neurotrophic Factor |
| HNPCs | Human Neural Progenitor Cells |
| AD | Alzheimer’s Disease |
| PD | Parkinson’s Disease |
| ALS | Amyotrophic Lateral Sclerosis |
| GDNF | Glial Cell-Derived Neurotrophic Factor |
| HDACi | Histone Deacetylase inhibitor |
| KDM | Lysine Demethylase |
| HAT | Histone Acetyltransferase |
| DNMT | DNA Methyltransferase |
| cDNA | Complementary DNA |
| HTS | High-Throughput Screening |
| RGC | Radial Glial Cells |
| ICC | Immunocytochemistry |
References
- Checkoway, H.; Lundin, J.I.; Kelada, S.N. Neurodegenerative Diseases; IARC Scientific Publications: Lyon, France, 2010; pp. 407–419. [Google Scholar]
- Qiu, X.; Xiao, X.; Li, N.; Li, Y. Histone deacetylases inhibitors (HDACis) as novel therapeutic application in various clinical diseases. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 72, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Glaser, K.B. HDAC inhibitors: Clinical update and mechanism-based potential. Biochem. Pharmacol. 2007, 74, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, A.G.; Thompson, L.M. Therapeutic application of histone deacetylase inhibitors for central nervous system disorders. Nat. Rev. Drug Discov. 2008, 7, 854. [Google Scholar] [CrossRef] [PubMed]
- Khan, O.; La Thangue, N.B. HDAC inhibitors in cancer biology: Emerging mechanisms and clinical applications. Immunol. Cell Biol. 2012, 90, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Casaccia-Bonnefil, P.; Carter, B.D.; Dobrowsky, R.T.; Chao, M.V. Death of oligodendrocytes mediated by the interaction of nerve growth factor with its receptor p75. Nature 1996, 383, 716. [Google Scholar] [CrossRef] [PubMed]
- DeFreitas, M.F.; McQuillen, P.S.; Shatz, C.J. A novel p75NTR signaling pathway promotes survival, not death, of immunopurified neocortical subplate neurons. J. Neurosci. 2001, 21, 5121–5129. [Google Scholar] [CrossRef] [PubMed]
- Ernfors, P.; Kucera, J.; Lee, K.F.; Loring, J.; Jaenisch, R. Studies on the physiological role of brain-derived neurotrophic factor and neurotrophin-3 in knockout mice. Int. J. Dev. Biol. 2003, 39, 799–807. [Google Scholar]
- Mao, L.-M.; Fibuch, E.E.; Wang, J.Q. Decoding BDNF-LTP coupling in cocaine addiction. Neuron 2010, 67, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Bekinschtein, P.; Cammarota, M.; Katche, C.; Slipczuk, L.; Rossato, J.I.; Goldin, A.; Izquierdo, I.; Medina, J.H. BDNF is essential to promote persistence of long-term memory storage. Proc. Natl. Acad. Sci. USA 2008, 105, 2711–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pencea, V.; Bingaman, K.D.; Wiegand, S.J.; Luskin, M.B. Infusion of brain-derived neurotrophic factor into the lateral ventricle of the adult rat leads to new neurons in the parenchyma of the striatum, septum, thalamus, and hypothalamus. J. Neurosci. 2001, 21, 6706–6717. [Google Scholar] [CrossRef] [PubMed]
- Pezet, S.; Malcangio, M. Brain-derived neurotrophic factor as a drug target for CNS disorders. Expert Opin. Ther. Targets 2004, 8, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Boulle, F.; Van Den Hove, D.L.A.; Jakob, S.B.; Rutten, B.P.; Hamon, M.; Van Os, J.; Lesch, K.P.; Lanfumey, L.; Steinbusch, H.W.; Kenis, G. Epigenetic regulation of the BDNF gene: Implications for psychiatric disorders. Mol. Psychiatry 2012, 17, 584. [Google Scholar] [CrossRef] [PubMed]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Miller, D.L.; Roecklein, K.A. The aging hippocampus: Interactions between exercise, depression, and BDNF. Neuroscientist 2012, 18, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Frielingsdorf, H.; Bath, K.G.; Soliman, F.; DiFede, J.; Casey, B.J.; Lee, F.S. Variant brain-derived neurotrophic factor Val66Met endophenotypes: Implications for posttraumatic stress disorder. Ann. N. Y. Acad. Sci. 2010, 1208, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Mahan, A.L.; Ressler, K.J. Fear conditioning, synaptic plasticity and the amygdala: Implications for posttraumatic stress disorder. Trends Neurosci. 2012, 35, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Sweatt, J.D. Epigenetic marking of the BDNF gene by early-life adverse experiences. Horm. Behav. 2011, 59, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nature 2004, 429, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, J.N.; Harris, K.M. Nanoscale analysis of structural synaptic plasticity. Curr. Opin. Neurobiol. 2012, 22, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Yu, X.; Perlik, A.J.; Tobin, W.F.; Zweig, J.A.; Tennant, K.; Jones, T.; Zuo, Y. Rapid formation and selective stabilization of synapses for enduring motor memories. Nature 2009, 462, 915–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger-Sweeney, J.; Hohmann, C.F. Behavioral consequences of abnormal cortical development: Insights into developmental disabilities. Behav. Brain Res. 1997, 86, 121–142. [Google Scholar] [CrossRef]
- Ramakers, G.J. Rho proteins, mental retardation and the cellular basis of cognition. Trends Neurosci. 2002, 25, 191–199. [Google Scholar] [CrossRef]
- Webb, S.J.; Monk, C.S.; Nelson, C.A. Mechanisms of postnatal neurobiological development: Implications for human development. Dev. Neuropsychol. 2001, 19, 147–171. [Google Scholar] [CrossRef] [PubMed]
- Radio, N.M.; Breier, J.M.; Shafer, T.J.; Mundy, W.R. Assessment of chemical effects on neurite outgrowth in PC12 cells using high content screening. Toxicol. Sci. 2008, 105, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, P.S.; Dallas, S.; Wilson, B.; Block, M.L.; Wang, C.-C.; Kinyamu, H.; Lu, N.; Gao, X.; Leng, Y. Histone deacetylase inhibitors up-regulate astrocyte GDNF and BDNF gene transcription and protect dopaminergic neurons. Int. J. Neuropsychopharmacol. 2008, 11, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Marini, A.M.; Lipsky, R.H. Effects of histone deacetylase inhibitor Trichostatin A on epigenetic changes and transcriptional activation of Bdnf promoter 1 by rat hippocampal neurons. Ann. N. Y. Acad. Sci. 2010, 1199, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.R.; Kim, J.-H.; Kim, Y.J.; Kwon, K.J.; Shin, C.Y.; Kim, H.Y.; Han, S.-H.; Choi, D.-H.; Lee, J. Effect of HDAC inhibitors on neuroprotection and neurite outgrowth in primary rat cortical neurons following ischemic insult. Neurochem. Res. 2013, 38, 1921–1934. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.; Frumkin, M.; Kassner, P.D. Cell-based screening: Extracting meaning from complex data. Neuron 2015, 86, 160–174. [Google Scholar] [CrossRef] [PubMed]
- An, W.F.; Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 2010, 45, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Astashkina, A.; Mann, B.; Grainger, D.W. A critical evaluation of in vitro cell culture models for high-throughput drug screening and toxicity. Pharmacol. Ther. 2012, 134, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Finan, G.M.; Realubit, R.; Chung, S.; Lütjohann, D.; Wang, N.; Cirrito, J.R.; Karan, C.; Kim, T.-W. Bioactive Compound Screen for Pharmacological Enhancers of Apolipoprotein E in Primary Human Astrocytes. Cell Chem. Biol. 2016, 23, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Magistri, M.; Khoury, N.; Mazza, E.M.C.; Velmeshev, D.; Lee, J.K.; Bicciato, S.; Tsoulfas, P.; Faghihi, M.A. A comparative transcriptomic analysis of astrocytes differentiation from human neural progenitor cells. Eur. J. Neurosci. 2016, 44, 2858–2870. [Google Scholar] [CrossRef] [PubMed]
- Bez, A.; Corsini, E.; Curti, D.; Biggiogera, M.; Colombo, A.; Nicosia, R.F.; Pagano, S.F.; Parati, E.A. Neurosphere and neurosphere-forming cells: Morphological and ultrastructural characterization. Brain Res. 2003, 993, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Buc-Caron, M.-H. Neuroepithelial progenitor cells explanted from human fetal brain proliferate and differentiatein vitro. Neurobiol. Dis. 1995, 2, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Dong, E.; Ruzicka, W.B.; Grayson, D.R.; Guidotti, A. DNA-methyltransferase1 (DNMT1) binding to CpG rich GABAergic and BDNF promoters is increased in the brain of schizophrenia and bipolar disorder patients. Schizophr. Res. 2015, 167, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Karatas, H.; Townsend, E.C.; Cao, F.; Chen, Y.; Bernard, D.; Liu, L.; Lei, M.; Dou, Y.; Wang, S. High-affinity, small-molecule peptidomimetic inhibitors of MLL1/WDR5 protein–protein interaction. J. Am. Chem. Soc. 2012, 135, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Madinier, A.; Bertrand, N.; Mossiat, C.; Prigent-Tessier, A.; Beley, A.; Marie, C.; Garnier, P. Microglial involvement in neuroplastic changes following focal brain ischemia in rats. PLoS ONE 2009, 4, e8101. [Google Scholar] [CrossRef] [PubMed]
- Besson, V.C.; Croci, N.; Boulu, R.G.; Plotkine, M.; Marchand-Verrecchia, C. Deleterious poly (ADP-ribose) polymerase-1 pathway activation in traumatic brain injury in rat. Brain Res. 2003, 989, 58–66. [Google Scholar] [CrossRef]
- Koh, S.-H.; Chang, D.-I.; Kim, H.-T.; Kim, J.; Kim, M.-H.; Kim, K.S.; Bae, I.; Kim, H.; Kim, D.W.; Kim, S.H. Effect of 3-aminobenzamide, PARP inhibitor, on matrix metalloproteinase-9 level in plasma and brain of ischemic stroke model. Toxicology 2005, 214, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Czapski, G.A.; Cakala, M.; Gajkowska, B.; Strosznajder, J.B. Poly (ADP-ribose) polymerase-1 inhibition protects the brain against systemic inflammation. Neurochem. Int. 2006, 49, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, F.; Bizzarri, B.; Maccioni, E.; Secci, D.; Bolasco, A.; Chimenti, P.; Fioravanti, R.; Granese, A.; Carradori, S.; Tosi, F. A novel histone acetyltransferase inhibitor modulating Gcn5 network: Cyclopentylidene-[4-(4′-chlorophenyl) thiazol-2-yl) hydrazone. J. Med. Chem. 2008, 52, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Leurs, U.; Clausen, R.P.; Kristensen, J.L.; Lohse, B. Inhibitor scaffold for the histone lysine demethylase KDM4C (JMJD2C). Bioorg. Med. Chem. Lett. 2012, 22, 5811–5813. [Google Scholar] [CrossRef] [PubMed]
- King, O.N.; Li, X.S.; Sakurai, M.; Kawamura, A.; Rose, N.R.; Ng, S.S.; Quinn, A.M.; Rai, G.; Mott, B.T.; Beswick, P. Quantitative high-throughput screening identifies 8-hydroxyquinolines as cell-active histone demethylase inhibitors. PLoS ONE 2010, 5, e15535. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.H.; Nielsen, A.L.; Helgstrand, C.; Lees, M.; Cloos, P.; Kastrup, J.S.; Helin, K.; Olsen, L.; Gajhede, M. Studies of H3K4me3 demethylation by KDM5B/Jarid1B/PLU1 reveals strong substrate recognition in vitro and identifies 2, 4-pyridine-dicarboxylic acid as an in vitro and in cell inhibitor. FEBS J. 2012, 279, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Markus, M.A.; Morris, B.J. Resveratrol in prevention and treatment of common clinical conditions of aging. Clin. Interv. Aging 2008, 3, 331. [Google Scholar] [PubMed]
- Saiko, P.; Szakmary, A.; Jaeger, W.; Szekeres, T. Resveratrol and its analogs: Defense against cancer, coronary disease and neurodegenerative maladies or just a fad? Mutat. Res./Rev. Mutat. Res. 2008, 658, 68–94. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Malinowski, B.; Węclewicz, M.M.; Grześk, E.; Grześk, G. Resveratrol increases serum BDNF concentrations and reduces vascular smooth muscle cells contractility via a NOS-3-independent mechanism. BioMed Res. Int. 2017, 2017, 9202954. [Google Scholar] [CrossRef] [PubMed]
- Rahvar, M.; Nikseresht, M.; Shafiee, S.M.; Naghibalhossaini, F.; Rasti, M.; Panjehshahin, M.R.; Owji, A.A. Effect of oral resveratrol on the BDNF gene expression in the hippocampus of the rat brain. Neurochem. Res. 2011, 36, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Shojaei, S.; Panjehshahin, M.R.; Shafiee, S.M.; Khoshdel, Z.; Borji, M.; Ghasempour, G.; Owji, A.A. Differential effects of resveratrol on the expression of brain-derived neurotrophic factor transcripts and protein in the hippocampus of rat brain. Iran. J. Med. Sci. 2017, 42, 32. [Google Scholar] [PubMed]
- Wang, X.; Xie, Y.; Zhang, T.; Bo, S.; Bai, X.; Liu, H.; Li, T.; Liu, S.; Zhou, Y.; Cong, X. Resveratrol reverses chronic restraint stress-induced depression-like behaviour: Involvement of BDNF level, ERK phosphorylation and expression of Bcl-2 and Bax in rats. Brain Res. Bull. 2016, 125, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and rat BDNF gene structure and expression revisited. J. Neurosci. Res. 2007, 85, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Pruunsild, P.; Kazantseva, A.; Aid, T.; Palm, K.; Timmusk, T. Dissecting the human BDNF locus: Bidirectional transcription, complex splicing, and multiple promoters. Genomics 2007, 90, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-R.; Lu, L.; Zhu, X.-G.; Gong, J.-P.; Shaham, Y.; Uhl, G.R. Rodent BDNF genes, novel promoters, novel splice variants, and regulation by cocaine. Brain Res. 2006, 1067, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-R.; Walther, D.; Drgon, T.; Polesskaya, O.; Lesnick, T.G.; Strain, K.J.; De Andrade, M.; Bower, J.H.; Maraganore, D.M.; Uhl, G.R. Human brain derived neurotrophic factor (BDNF) genes, splicing patterns, and assessments of associations with substance abuse and Parkinson’s Disease. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2005, 134, 93–103. [Google Scholar] [CrossRef] [PubMed]
- An, J.J.; Gharami, K.; Liao, G.-Y.; Woo, N.H.; Lau, A.G.; Vanevski, F.; Torre, E.R.; Jones, K.R.; Feng, Y.; Lu, B. Distinct role of long 3′ UTR BDNF mRNA in spine morphology and synaptic plasticity in hippocampal neurons. Cell 2008, 134, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Metsis, M. Regulation of BDNF promoters in the rat hippocampus. Neurochem. Int. 1994, 25, 11–15. [Google Scholar]
- Lau, A.G.; Irier, H.A.; Gu, J.; Tian, D.; Ku, L.; Liu, G.; Xia, M.; Fritsch, B.; Zheng, J.Q.; Dingledine, R. Distinct 3′ UTRs differentially regulate activity-dependent translation of brain-derived neurotrophic factor (BDNF). Proc. Natl. Acad. Sci. USA 2010, 107, 15945–15950. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Kanetaka, H.; Mochizuki, K.; Tominami, K.; Nunome, S.; Abe, G.; Kosukegawa, H.; Abe, T.; Mori, H.; Mori, K. Induction of neurite outgrowth in PC12 cells treated with temperature-controlled repeated thermal stimulation. PLoS ONE 2015, 10, e0124024. [Google Scholar] [CrossRef] [PubMed]
- Higgins, S.; Lee, J.S.; Ha, L.; Lim, J.Y. Inducing neurite outgrowth by mechanical cell stretch. BioResearch Open Access 2013, 2, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, R.; Ueno, M.; Yamashita, T. Intrinsic regenerative mechanisms of central nervous system neurons. Biosci. Trends 2009, 3, 179–183. [Google Scholar] [PubMed]
- Read, D.E.; Herbert, K.R.; Gorman, A.M. Heat shock enhances NGF-induced neurite elongation which is not mediated by Hsp25 in PC12 cells. Brain Res. 2008, 1221, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Magistri, M.; Velmeshev, D.; Makhmutova, M.; Faghihi, M.A. Transcriptomics profiling of Alzheimer’s Disease reveal neurovascular defects, altered amyloid-β homeostasis, and deregulated expression of long noncoding RNAs. J. Alzheimer’s Dis. 2015, 48, 647–665. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Category | |
|---|---|---|
| 1 | CAY10603 | HDAC Inhibitor |
| 2 | Givinostat (ITF2357) | |
| 3 | JNJ-26481585 (Quisinostat) | |
| 4 | PXD101 (Belinostat) | |
| 5 | M-344 | |
| 6 | PCI-24781 (Abexinostat) | |
| 7 | LMK 235 | |
| 8 | SB-939 (Pracinostat) | |
| 9 | Tubastatin A | |
| 10 | (S)-HDAC-42 (AR42) | |
| 11 | CAY10398 | |
| 12 | Phenylbutyrate·Na | |
| 13 | SAHA | |
| 14 | BML-210 | |
| 15 | Valproic acid | |
| 16 | Apicidin | |
| 17 | CUDC-907 | |
| 18 | Oxamflatin | |
| 19 | Trichostatin A | |
| 20 | MM-102 | Methyltransferase Inhibitor |
| 21 | 5-Aza-2′-deoxycytidine (Decitabine) | |
| 22 | Nicotinamide (Niacinamide) | PARP-1 Inhibitor |
| 23 | BYK 204165 | |
| 24 | CPTH2 | HAT Inhibitor |
| 25 | 2,4-Pyridinedicarboxylic Acid (2,4-PDCA) | Histone Demethylase Inhibitor |
| 26 | Resveratrol | Natural Compound |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagheri, A.; Habibzadeh, P.; Razavipour, S.F.; Volmar, C.-H.; Chee, N.T.; Brothers, S.P.; Wahlestedt, C.; Mowla, S.J.; Faghihi, M.A. HDAC Inhibitors Induce BDNF Expression and Promote Neurite Outgrowth in Human Neural Progenitor Cells-Derived Neurons. Int. J. Mol. Sci. 2019, 20, 1109. https://doi.org/10.3390/ijms20051109
Bagheri A, Habibzadeh P, Razavipour SF, Volmar C-H, Chee NT, Brothers SP, Wahlestedt C, Mowla SJ, Faghihi MA. HDAC Inhibitors Induce BDNF Expression and Promote Neurite Outgrowth in Human Neural Progenitor Cells-Derived Neurons. International Journal of Molecular Sciences. 2019; 20(5):1109. https://doi.org/10.3390/ijms20051109
Chicago/Turabian StyleBagheri, Amir, Parham Habibzadeh, Seyedeh Fatemeh Razavipour, Claude-Henry Volmar, Nancy T. Chee, Shaun P. Brothers, Claes Wahlestedt, Seyed Javad Mowla, and Mohammad Ali Faghihi. 2019. "HDAC Inhibitors Induce BDNF Expression and Promote Neurite Outgrowth in Human Neural Progenitor Cells-Derived Neurons" International Journal of Molecular Sciences 20, no. 5: 1109. https://doi.org/10.3390/ijms20051109