Salacia chinensis L. Stem Extract Exerts Antifibrotic Effects on Human Hepatic Stellate Cells through the Inhibition of the TGF-β1-Induced SMAD2/3 Signaling Pathway

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Results
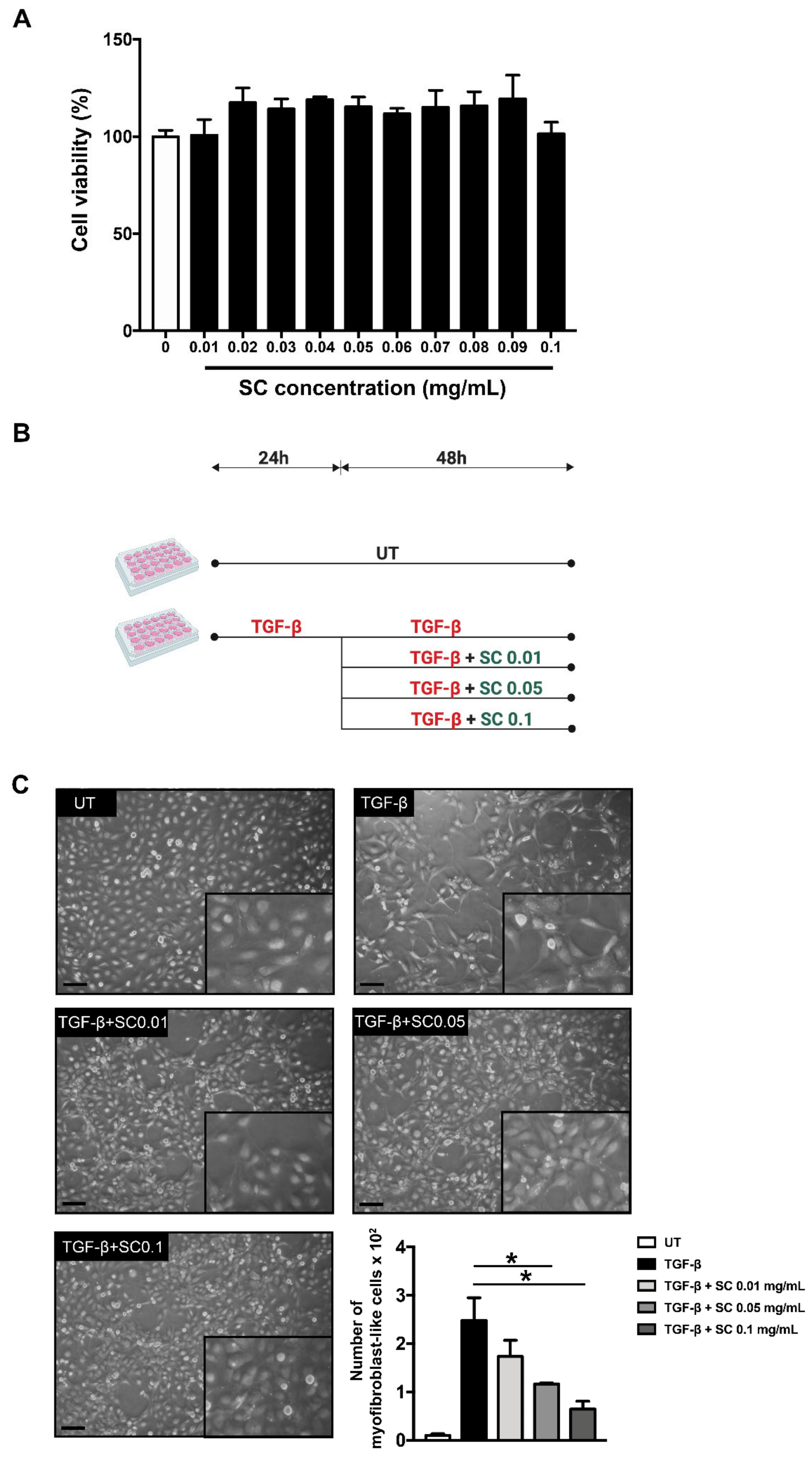
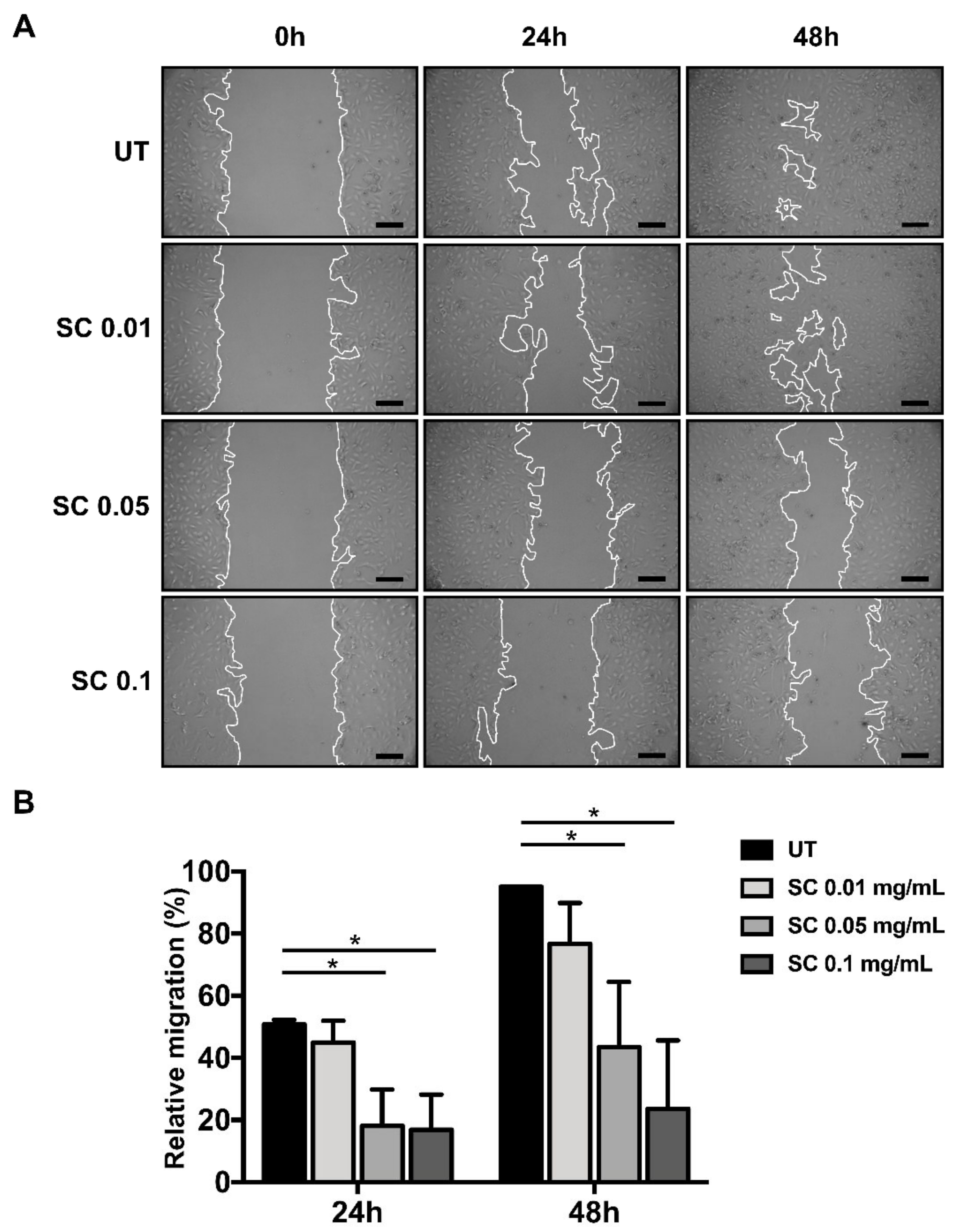
2.1. Salacia chinensis L. (SC) Stem Extract Reverses Morphology of HSCs Activation and Suppresses Its Migration via TGF-β1
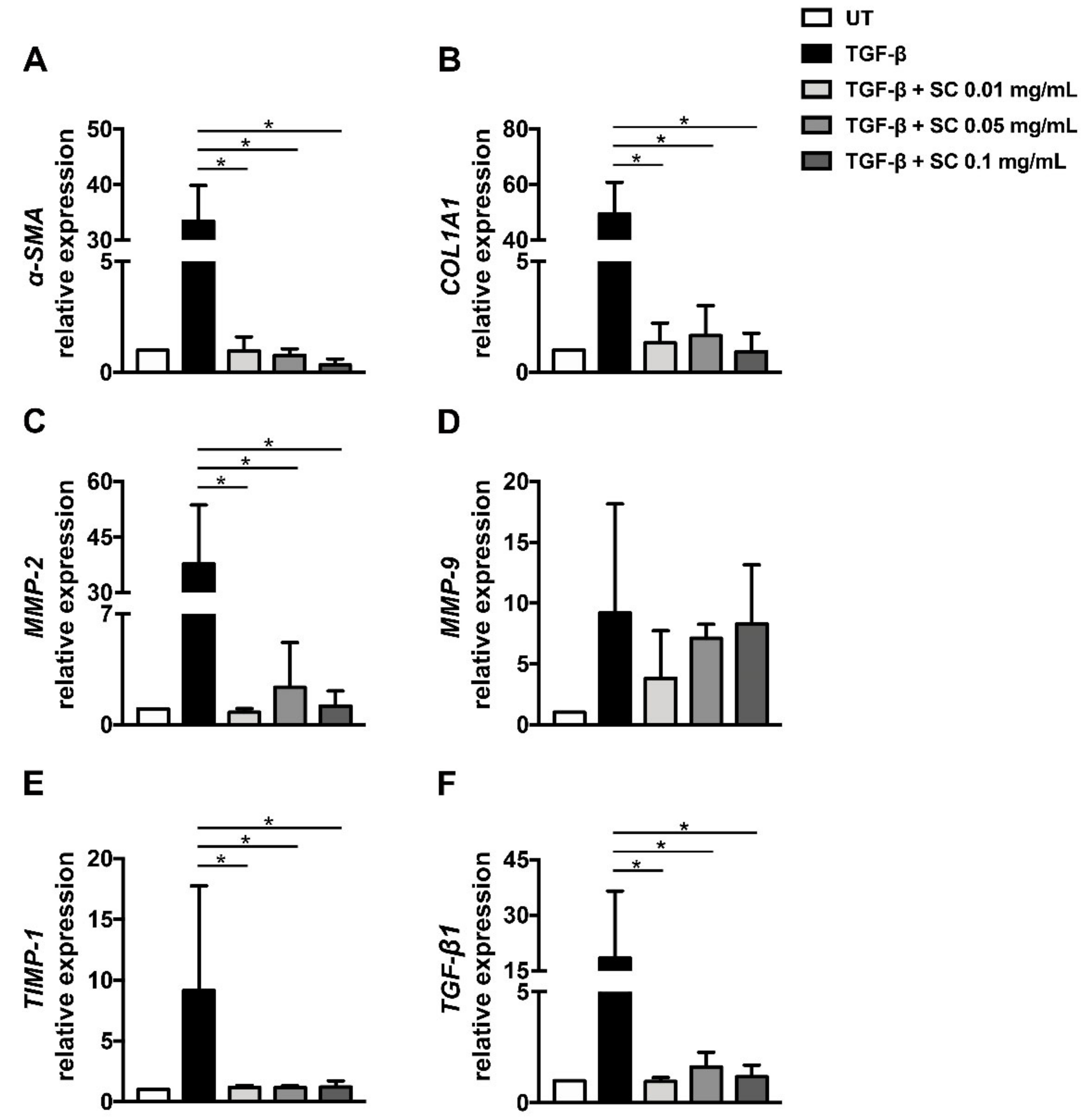
2.2. Salacia chinensis L. (SC) Stem Extract Suppresses Fibrotic Markers at the Gene Expression Level
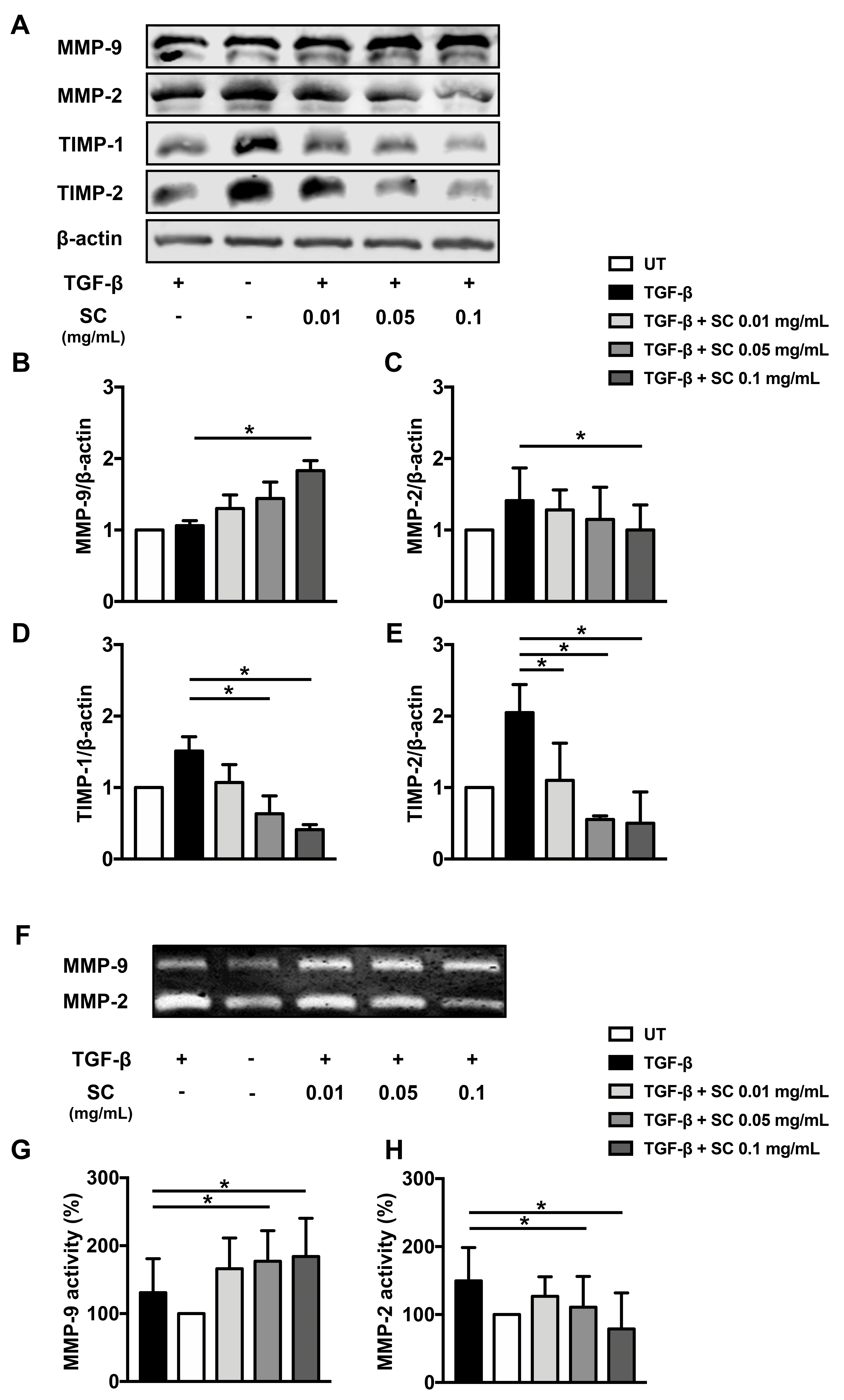
2.3. Translational Levels of Fibrotic Markers Are Suppressed by Salacia chinensis L. (SC) Stem Extract
2.4. Salacia Chinensis L. (SC) Stem Extract Reduced ECM Components
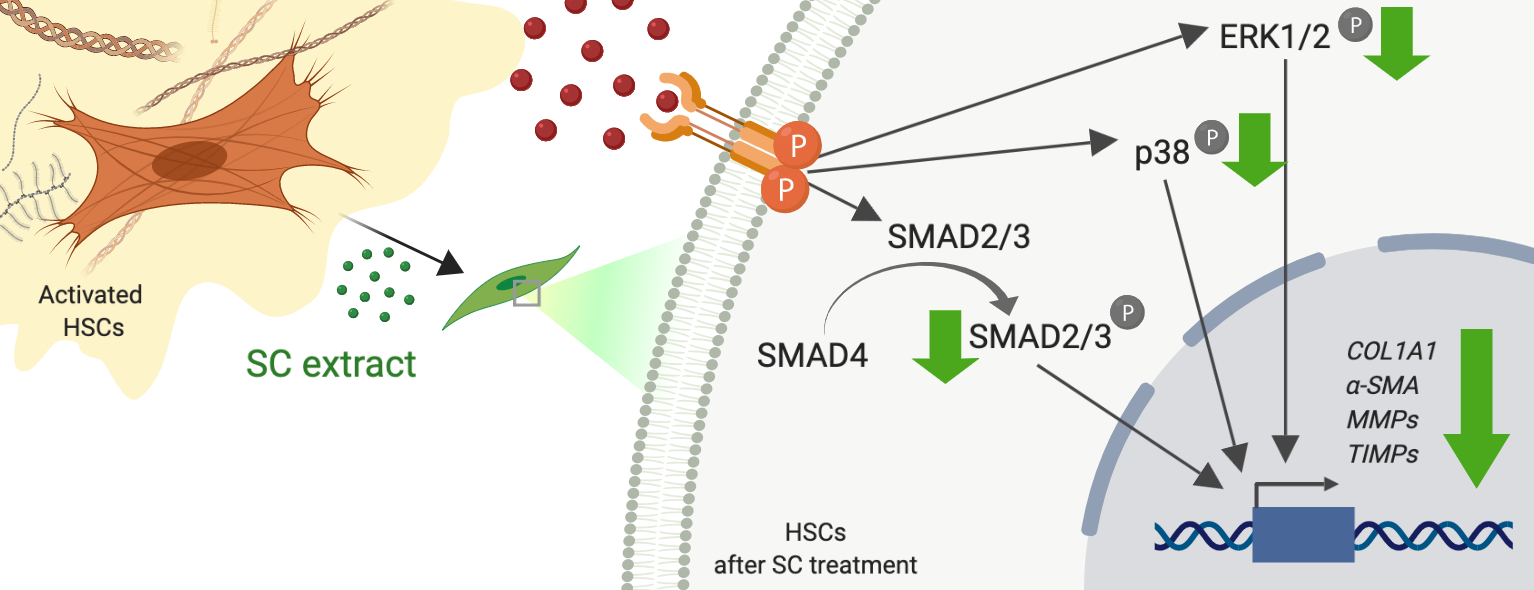
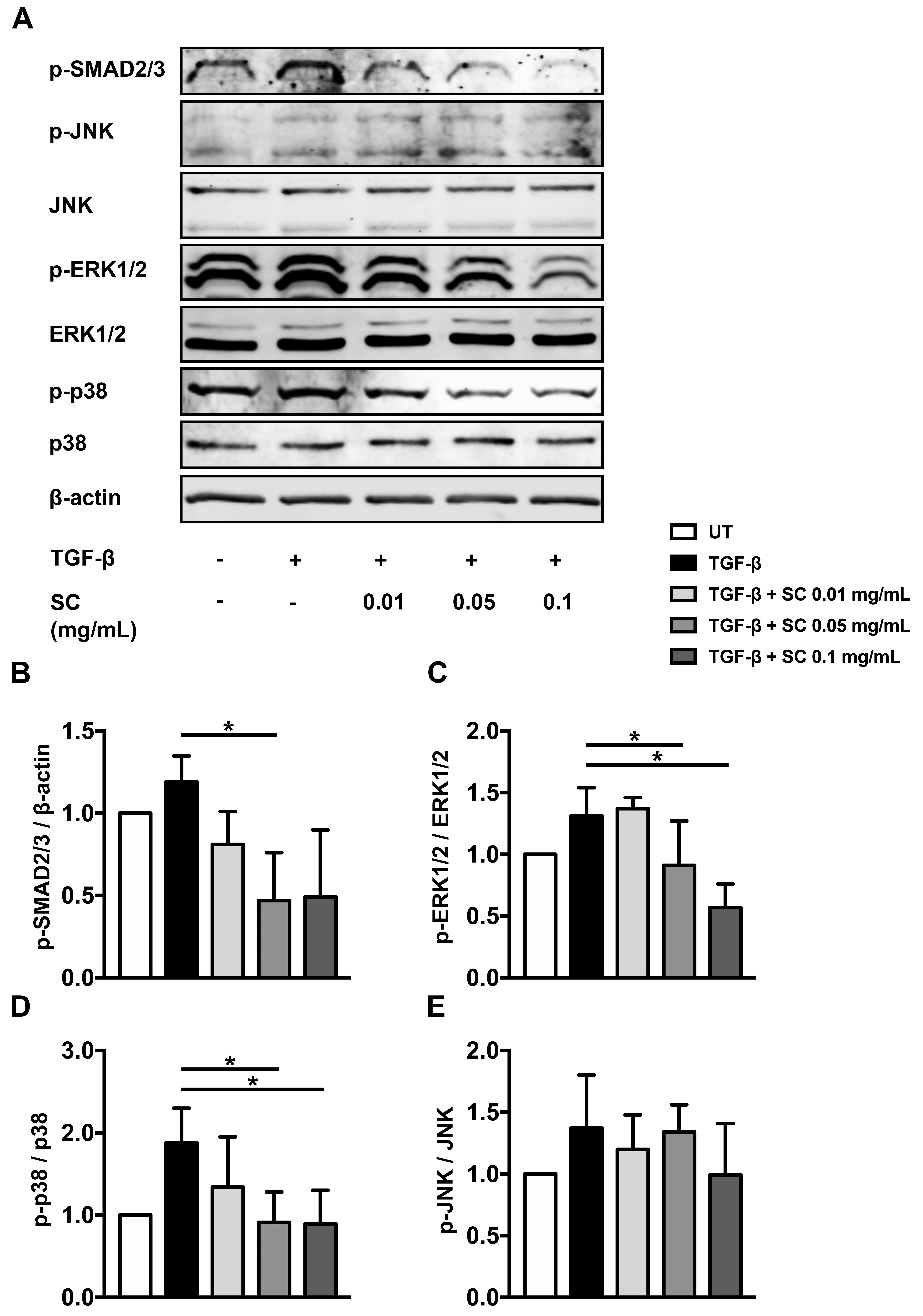
2.5. Salacia chinensis L. (SC) Stem Extract Ameliorates the Phosphorylations of SMAD2/3, ERK1/2, and p38 Induced by TGF-β1
3. Discussion
4. Materials and Methods
4.1. Preparation and Characterization of SC Stem Extract
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell Migration Assay
4.5. Immunofluorescence Study
4.6. Western Blot Analysis
4.7. Gelatin Zymography Assay
4.8. Quantitative Real-Time PCR
4.9. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase |
| ALT | valanine aminotransferase |
| AST | aspartate aminotransferase |
| COL1A1 | collagen type I alpha 1 chain |
| ECM | extracellular matrix |
| ERK | extracellular signal- regulated kinase |
| HSCs | hepatic stellate cells |
| JNK | c-Jun N-terminal kinase/stress-activated protein kinase |
| LX-2 | human hepatic stellate cell line |
| MAPK | mitogen activated protein kinases |
| MMPs | matrix metalloproteases |
| SC | Salacia chinensis L. |
| α-SMA | alpha-smooth muscle actin |
| SMAD | homology to Caenorhabditis elegans sma and drosophila gene mothers against decapentaplegic (MAD) proteins |
| TGF-β1 | transforming growth factors-beta1 |
| TIMPs | tissue inhibitors of the MMPs |
References
- Hernandez-gea, V.; Friedman, S.L. Pathogenesis of Liver Fibrosis. Ann. Rev. Pathol. Mech. Dis. 2011, 6, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, X.; Koyama, Y.; Wang, P.; Lan, T.; Kim, I.G.; Kim, I.H.; Ma, H.Y.; Kisseleva, T. The types of hepatic myofibroblasts contributing to liver fibrosis of different etiologies. Front. Pharmacol. 2014, 5, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Zhang, Z.; Wang, F.S. Liver fibrosis: Mechanisms of immune-mediated liver injury. Cell. Mol. Immunol. 2012, 9, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Galli, A.; Bochaton-Piallat, M.L.; Gabbiani, G. The myofibroblast: One function, multiple origins. Am. J. Pathol. 2007, 170, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Mormone, E.; George, J.; Nieto, N. Molecular pathogenesis of hepatic fibrosis and current therapeutic approaches. Chem. Biol. Interact. 2011, 193, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautwein, C.; Friedman, S.L.; Schuppan, D.; Pinzani, M. Hepatic fibrosis: Concept to treatment. J. Hepatol. 2015, 62, S15–S24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elpek, G.O. Cellular and molecular mechanisms in the pathogenesis of liver fibrosis: An update. World J. Gastroenterol. 2014, 20, 7260–7276. [Google Scholar] [CrossRef]
- Giannandrea, M.; Parks, W.C. Diverse functions of matrix metalloproteinases during fibrosis. Dis. Model. Mech. 2014, 7, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Baber, J.; Fujii, T.; Coito, A.J. Matrix metalloproteinases in liver injury, repair and fibrosis. Matrix Biol. 2015, 44, 147–156. [Google Scholar] [CrossRef]
- Arpino, V.; Brock, M.; Gill, S.E. The role of TIMPs in regulation of extracellular matrix proteolysis. Matrix Biol. 2015, 44, 247–254. [Google Scholar] [CrossRef]
- Park, S.A.; Kim, M.J.; Park, S.Y.; Kim, J.S.; Lim, W.; Nam, J.S.; Yhong Sheen, Y. TIMP-1 mediates TGF-β-dependent crosstalk between hepatic stellate and cancer cells via FAK signaling. Sci. Rep. 2015, 5, 16492. [Google Scholar] [CrossRef] [PubMed]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-β signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Z.; Kwong, S.Q.; Lui, E.L.H.; Friedman, S.L.; Li, F.R.; Lam, R.W.C.; Zhang, G.C.; Zhang, H.; Ye, T. Inhibition of PDGF, TGF-β, and Abl signaling and reduction of liver fibrosis by the small molecule Bcr-Abl tyrosine kinase antagonist Nilotinib. J. Hepatol. 2011, 55, 612–625. [Google Scholar] [CrossRef]
- Zhang, Y.E. Non-Smad pathways in TGF-β signaling. Cell Res. 2009, 19, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Bataller, R.; Brenner, D. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Schuppan, D.; Ashfaq-Khan, M.; Yang, A.T.; Kim, Y.O. Liver fibrosis: Direct antifibrotic agents and targeted therapies. Matrix Biol. 2018, 68, 435–451. [Google Scholar] [CrossRef]
- Koyama, Y.; Xu, J.; Liu, X.; Brenner, D.A. New Developments on the Treatment of Liver Fibrosis. Dig. Dis. 2016, 34, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Chavan, J.J.; Jagtap, U.B.; Gaikwad, N.B.; Dixit, G.B.; Bapat, V.A. Total phenolics, flavonoids and antioxidant activity of Saptarangi (Salacia chinensis L.) fruit pulp. J. Plant Biochem. Biotechnol. 2013, 22, 409–413. [Google Scholar] [CrossRef]
- Chavan, J.J.; Ghadage, D.M.; Bhoite, A.S.; Umdale, S.D. Micropropagation, molecular profiling and RP-HPLC determination of mangiferin across various regeneration stages of Saptarangi (Salacia chinensis L.). Ind. Crops Prod. 2015, 76, 1123–1132. [Google Scholar] [CrossRef]
- Ghadage, D.M.; Kshirsagar, P.R.; Pai, S.R.; Chavan, J.J. Extraction efficiency, phytochemical profiles and antioxidative properties of different parts of Saptarangi (Salacia chinensis L.)–An important underutilized plant. Biochem. Biophys. Rep. 2017, 12, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; Nguyen, T.H.A.; Vu, D.T.; Tran, V.S. Study on chemical constituents of Salacia chinensis L., collected in Thua Thien Hue. Tap Chi Hoa Hoc 2008, 46, 47–51. [Google Scholar]
- Morikawa, T.; Akaki, J.; Ninomiya, K.; Kinouchi, E.; Tanabe, G.; Pongpiriyadacha, Y.; Yoshikawa, M.; Muraoka, O. Salacinol and related analogs: New leads for type 2 diabetes therapeutic candidates from the Thai traditional natural medicine Salacia chinensis. Nutrients 2015, 7, 1480–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, M.; Shimoda, H.; Nishida, N.; Takada, M.; Matsuda, H. Salacia reticulata and Its Polyphenolic Constituents with Lipase Inhibitory and Lipolytic Activities Have Mild Antiobesity Effects in Rats. J. Nutr. 2002, 132, 1819–1824. [Google Scholar] [CrossRef] [Green Version]
- Friedman, L.G.; Qureshi, Y.H.; Yu, W.H. Promoting Autophagic Clearance: Viable Therapeutic Targets in Alzheimer’s Disease. Neurotherapeutics 2015, 12, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Hamada, T.; Kuriyama, N.; Busuttil, R.W.; Coito, A.J. TIMP-1 deficiency leads to lethal partial hepatic ischemia and reperfusion injury. Hepatology 2012, 56, 1074–1085. [Google Scholar] [CrossRef] [Green Version]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Wang, F.S.; Fan, J.G.; Zhang, Z.; Gao, B.; Wang, H.Y. The global burden of liver disease: The major impact of China. Hepatology 2014, 60, 2099–2108. [Google Scholar] [CrossRef]
- Nakamura, S.; Zhang, Y.; Matsuda, H.; Ninomiya, K.; Muraoka, O.; Yoshikawa, M. Chemical structures and hepatoprotective effects of constituents from the leaves of Salacia chinensis. Chem. Pharm. Bull. 2011, 59, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- El-Lakkany, N.M.; El-Maadawy, W.H.; Seif el-Din, S.H.; Saleh, S.; Safar, M.M.; Ezzat, S.M.; Mohamed, S.H.; Botros, S.S.; Demerdash, Z.; Hammam, O.A. Antifibrotic effects of gallic acid on hepatic stellate cells: In vitro and in vivo mechanistic study. J. Tradit. Complement. Med. 2019, 9, 45–53. [Google Scholar] [CrossRef]
- Bi, W.-R.; Yang, C.-Q.; Shi, Q. Transforming growth factor-β1 induced epithelial-mesenchymal transition in hepatic fibrosis. Hepatogastroenterology 2012, 59, 1960–1963. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.-H.; Derynck, R. Specificity and Versatility in Tgf-Β Signaling Through Smads. Annu. Rev. Cell Dev. Biol. 2005, 21, 659–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrecchia, F.; Mauviel, A. Transforming growth factor-beta and fibrosis. World J. Gastroenterol. 2007, 13, 3056–3062. [Google Scholar] [CrossRef] [PubMed]
- Pera, E.M.; Ikeda, A.; Eivers, E.; De Robertis, E.M. Integration of IGF, FGF, and anti-BMP signals via Smad1 phosphorylation in neural induction. Genes Dev. 2003, 17, 3023–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretzschmar, M.; Doody, J.; Massagué, J. Opposing BMP and EGF signalling pathways converge on the TGF-β family mediator Smad1. Nature 1997, 389, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.E.; McDonnell, M.A.; Law, B.K.; Moses, H.L. Interdependent SMAD and JNK signaling in transforming growth factor-beta-mediated transcription. J. Biol. Chem. 1999, 274, 37413–37420. [Google Scholar] [CrossRef] [Green Version]
- De Caestecker, M.P.; Parks, W.T.; Frank, C.J.; Castagnino, P.; Bottaro, D.P.; Roberts, A.B.; Lechleider, R.J. Smad2 transduces common signals from receptor serine−threonine and tyrosine kinases Smad2 transduces common signals from receptor serine–threonine and tyrosine kinases. Genes Dev. 1998, 12, 1587–1592. [Google Scholar] [CrossRef] [Green Version]
- Préaux, A.M.; Mallat, A.; Nhieu, J.T.; D’Ortho, M.P.; Hembry, R.M.; Mavier, P. Matrix metalloproteinase-2 activation in human hepatic fibrosis regulation by cell-matrix interactions. Hepatology 1999, 30, 944–950. [Google Scholar] [CrossRef]
- Henderson, N.C.; Iredale, J.P. Liver fibrosis: Cellular mechanisms of progression and resolution. Clin. Sci. 2007, 112, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Lee, U.E.; Friedman, S.L. Mechanisms of hepatic fibrogenesis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 195–206. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2000, 1477, 267–283. [Google Scholar] [CrossRef]
- Thiele, N.D.; Wirth, J.W.; Steins, D.; Koop, A.C.; Ittrich, H.; Lohse, A.W.; Kluwe, J. TIMP-1 is upregulated, but not essential in hepatic fibrogenesis and carcinogenesis in mice. Sci. Rep. 2017, 7, 714. [Google Scholar] [CrossRef] [PubMed]
- Yoshiji, H.; Kuriyama, S.; Miyamoto, Y.; Thorgeirsson, U.; Gomez, D.; Kawata, M.; Yoshii, J.; Ikenaka, Y.; Noguchi, R.; Tsujinoue, H.; et al. Tissue inhibitor of metalloproteinases-1 promotes liver fibrosis development in a transgenic mouse model. Hepatology 2000, 32, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Arthur, M.J.P.; Iredale, J.P.; Mann, D.A. Tissue Inhibitors of Metalloproteinases: Role in Liver Fibrosis and Alcoholic Liver Disease. Alcohol. Clin. Exp. Res. 1999, 23, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Arthur, M. Fibrogenesis II. Metalloproteases and their Inhibitors in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G245–G249. [Google Scholar] [CrossRef]
- Murawaki, Y.; Yamada, S.; Ikuta, Y.; Kawasaki, H. Clinical usefulness of serum matrix metalloproteinase-2 concentration in patients with chronic viral liver disease. J. Hepatol. 1999, 30, 1090–1098. [Google Scholar] [CrossRef]
- Prystupa, A.; Boguszewska-Czubara, A.; Bojarska-Junak, A.; Toruń-Jurkowska, A.; Roliński, J.; Załuska, W. Activity of MMP-2, MMP-8 and MMP-9 in serum as a marker of progression of alcoholic liver disease in people from Lublin region, eastern Poland. Ann. Agric. Environ. Med. 2015, 22, 325–328. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Fold Change of Protein Production | Ratio of Production | ||||||
|---|---|---|---|---|---|---|---|---|
| MMP-1 | MMP-2 | MMP-9 | TIMP-1 | TIMP-2 | ||||
| UT | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 ± 0.00 | 1 | 1 | 1 |
| TGF-β | 1.20 ± 0.08 | 1.41 ± 0.46 | 0.96 ± 0.07 | 1.5 ± 0.20 | 2.05 ± 0.39 | 0.80 | 0.69 | 0.64 |
| TGF-β + SC0.01 | 1.20 ± 0.01 | 1.28 ± 0.28 | 1.30 ± 0.19 | 1.07 ± 0.25 | 1.10 ± 0.52 | 0.63 | 0.62 | 1.27 |
| TGF-β + SC0.05 | 1.17 ± 0.13 | 1.15 ± 0.45 | 1.44 ± 0.23 | 0.63 ± 0.25 | 0.55 ± 0.05 | 1.85 * | 2.08 | 2.29 |
| TGF-β + SC0.1 | 0.77 ± 0.07 | 1.00 ± 0.35 | 1.83 ± 0.14 | 0.41 ± 0.07 | 0.50 ± 0.44 | 1.87 * | 2.00 | 4.46 * |
| Genes | Forward Primer (F) | Reverse Primer (R) |
|---|---|---|
| α-SMA | 5′-CGCATCCTCATCCTCCCT-3′ | 5′- GGCCGTGATCTCCTTCTG-3′ |
| COL1A1 | 5′-GTCGAGGGCCAAGACGAAG-3′ | 5′-CAGATCACGTCATCGCACAAC-3′ |
| MMP-2 | 5′-ACATCAAGGGCATTCAGGAG-3′ | 5′-GCCTCCGTATACCGCATCAAT-3′ |
| MMP-9 | 5′-CCCGGAGTGAGTTGAACCA-3′ | 5′-GGATTTACATGGCACTGCCA-3′ |
| TIMP-1 | 5′-CTTCTGCAATTCCGACCTCGT-3′ | 5′-CCCTAAGGCTTGGAACCCTTT-3′ |
| TGF-β1 | 5′-GGCCAGATCCTGTCCAAGC-3′ | 5′-GTGGGTTTCCACCATTAGCAC-3′ |
| GAPDH | 5′-ATGGGGAAGGTGAAGGTCG-3′ | 5′-GGGGTCATTGATGGCAACAATA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phaosri, M.; Jantrapirom, S.; Na Takuathung, M.; Soonthornchareonnon, N.; Sireeratawong, S.; Buacheen, P.; Pitchakarn, P.; Nimlamool, W.; Potikanond, S. Salacia chinensis L. Stem Extract Exerts Antifibrotic Effects on Human Hepatic Stellate Cells through the Inhibition of the TGF-β1-Induced SMAD2/3 Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 6314. https://doi.org/10.3390/ijms20246314
Phaosri M, Jantrapirom S, Na Takuathung M, Soonthornchareonnon N, Sireeratawong S, Buacheen P, Pitchakarn P, Nimlamool W, Potikanond S. Salacia chinensis L. Stem Extract Exerts Antifibrotic Effects on Human Hepatic Stellate Cells through the Inhibition of the TGF-β1-Induced SMAD2/3 Signaling Pathway. International Journal of Molecular Sciences. 2019; 20(24):6314. https://doi.org/10.3390/ijms20246314
Chicago/Turabian StylePhaosri, Mattareeyapar, Salinee Jantrapirom, Mingkwan Na Takuathung, Noppamas Soonthornchareonnon, Seewaboon Sireeratawong, Pensiri Buacheen, Pornsiri Pitchakarn, Wutigri Nimlamool, and Saranyapin Potikanond. 2019. "Salacia chinensis L. Stem Extract Exerts Antifibrotic Effects on Human Hepatic Stellate Cells through the Inhibition of the TGF-β1-Induced SMAD2/3 Signaling Pathway" International Journal of Molecular Sciences 20, no. 24: 6314. https://doi.org/10.3390/ijms20246314