Combined Treatment with Low-Dose Ionizing Radiation and Ketamine Induces Adverse Changes in CA1 Neuronal Structure in Male Murine Hippocampi


 , , ,
, , , 
Abstract
:1. Introduction
2. Results
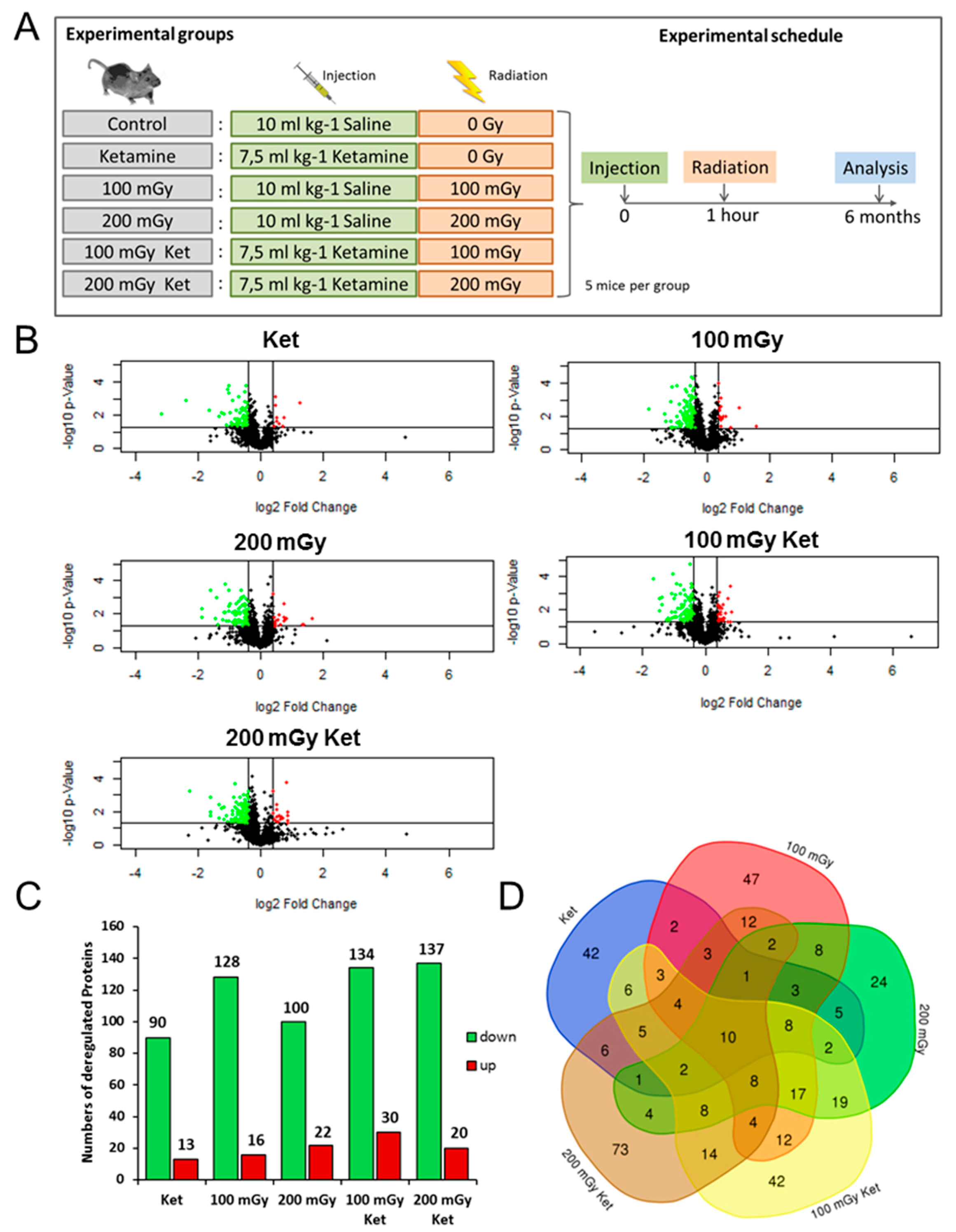
2.1. Analysis of the Hippocampal Proteome after Single or Combined Treatment
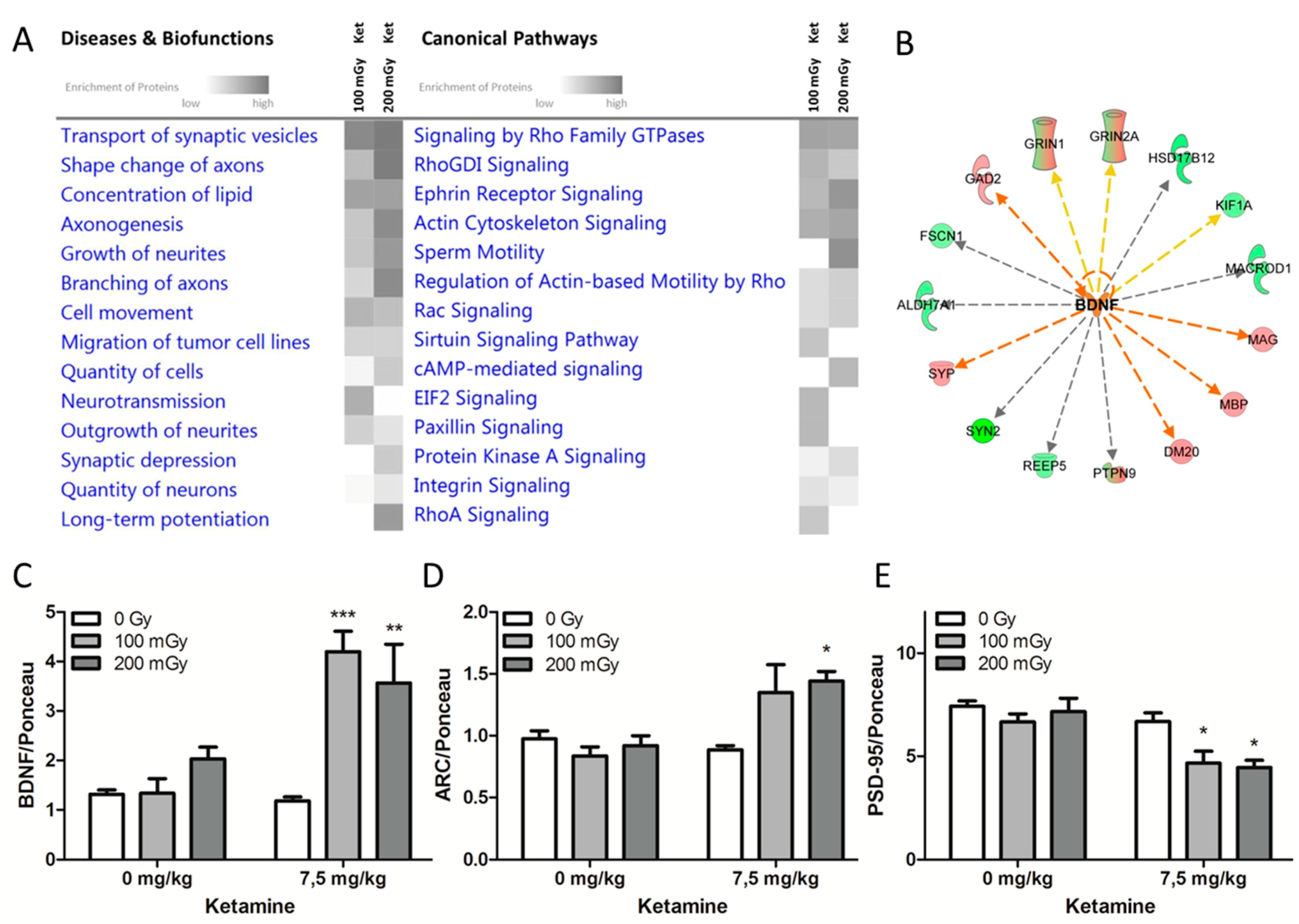
2.2. Effects on Neuronal Cytoskeleton and Synaptic Plasticity Following Combined Exposure to Ketamine and Irradiation
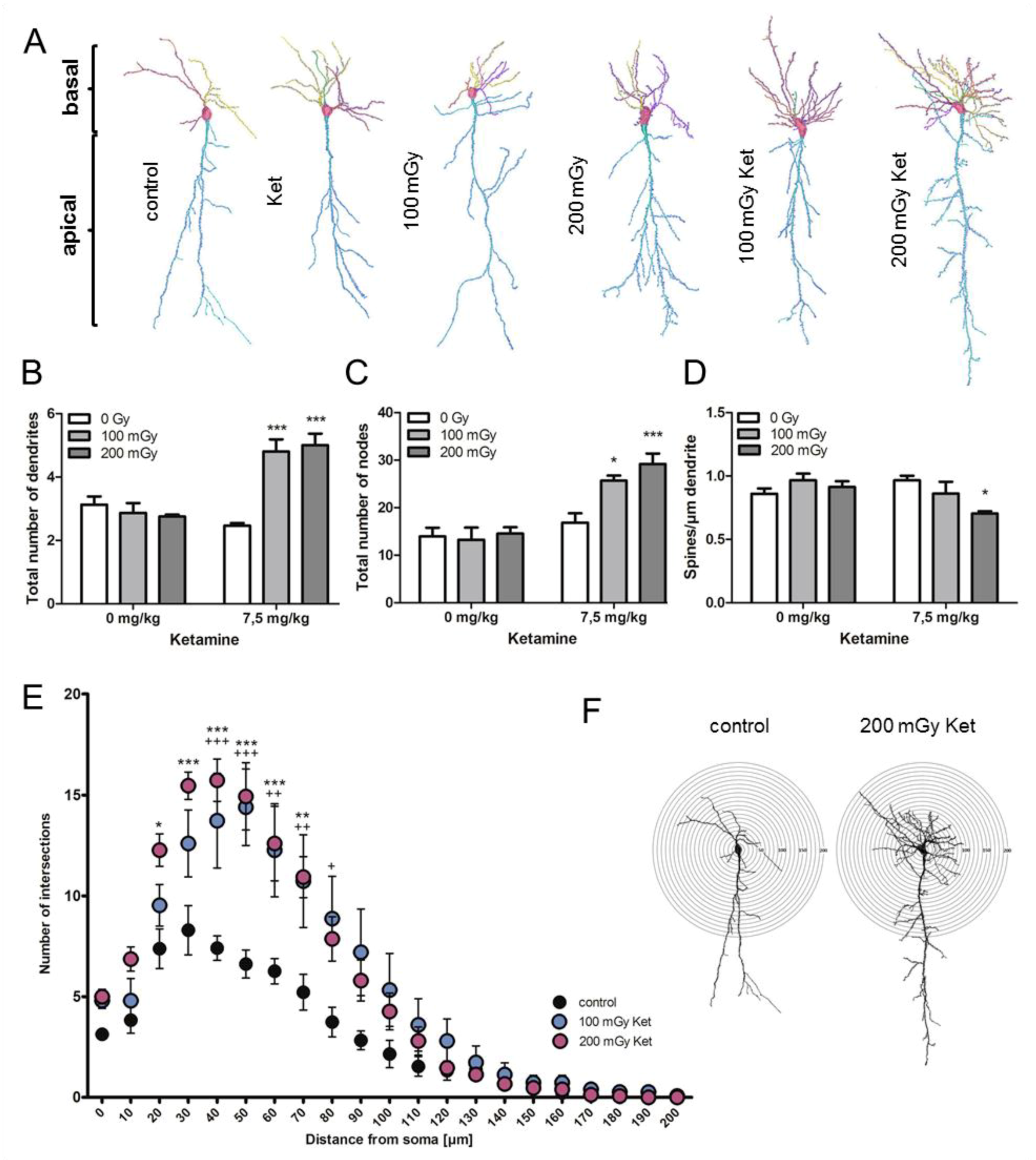
2.3. Morphological Abnormalities of Hippocampal CA1 Neurons only after Combined Treatment with Ketamine and Irradiation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Exposure
4.3. Tissue Collection
4.4. Protein Lysis and Determination of Protein Concentration
4.5. Mass Spectrometry (MS)
4.6. Protein Identification and Quantification
4.7. Pathway Analysis
4.8. Western Blot Analysis
4.9. Golgi Staining
4.10. Imaging and Analysis of Dendrites and Spines
4.11. Statistical Analysis
4.12. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miglioretti, D.L.; Johnson, E.; Williams, A.; Greenlee, R.T.; Weinmann, S.; Solberg, L.I.; Feigelson, H.S.; Roblin, D.; Flynn, M.J.; Vanneman, N.; et al. The use of computed tomography in pediatrics and the associated radiation exposure and estimated cancer risk. JAMA Pediatr. 2013, 167, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.S.; Salotti, J.A.; Little, M.P.; McHugh, K.; Lee, C.; Kim, K.P.; Howe, N.L.; Ronckers, C.M.; Rajaraman, P.; Sir Craft, A.W.; et al. Radiation exposure from CT scans in childhood and subsequent risk of leukaemia and brain tumours: A retrospective cohort study. Lancet 2012, 380, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.X.; Kachniarz, B.; Gilani, S.; Shin, J.J. Risk of malignancy associated with head and neck CT in children: A systematic review. Otolaryngol. Head Neck Surg. 2014, 151, 554–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMullen, K.P.; Hanson, T.; Bratton, J.; Johnstone, P.A. Parameters of anesthesia/sedation in children receiving radiotherapy. Radiat. Oncol. 2015, 10, 65. [Google Scholar] [CrossRef] [Green Version]
- Green, S.M.; Roback, M.G.; Kennedy, R.M.; Krauss, B. Clinical practice guideline for emergency department ketamine dissociative sedation: 2011 update. Ann. Emerg. Med. 2011, 57, 449–461. [Google Scholar] [CrossRef]
- Li, F.; Tsien, J.Z. Memory and the NMDA receptors. N. Engl. J. Med. 2009, 361, 302–303. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, H.; Singh, S.K.; Mancusso, R.; Gouaux, E. Subunit arrangement and function in NMDA receptors. Nature 2005, 438, 185–192. [Google Scholar] [CrossRef]
- Duchen, M.R.; Burton, N.R.; Biscoe, T.J. An intracellular study of the interactions of N-methyl-DL-aspartate with ketamine in the mouse hippocampal slice. Brain Res. 1985, 342, 149–153. [Google Scholar] [CrossRef]
- Fredriksson, A.; Archer, T.; Alm, H.; Gordh, T.; Eriksson, P. Neurofunctional deficits and potentiated apoptosis by neonatal NMDA antagonist administration. Behav. Brain Res. 2004, 153, 367–376. [Google Scholar] [CrossRef]
- Viberg, H.; Ponten, E.; Eriksson, P.; Gordh, T.; Fredriksson, A. Neonatal ketamine exposure results in changes in biochemical substrates of neuronal growth and synaptogenesis, and alters adult behavior irreversibly. Toxicology 2008, 249, 153–159. [Google Scholar] [CrossRef]
- Fredriksson, A.; Archer, T. Hyperactivity following postnatal NMDA antagonist treatment: Reversal by D-amphetamine. Neurotox. Res. 2003, 5, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Buratovic, S.; Stenerlow, B.; Sundell-Bergman, S.; Fredriksson, A.; Viberg, H.; Gordh, T.; Eriksson, P. Effects on adult cognitive function after neonatal exposure to clinically relevant doses of ionising radiation and ketamine in mice. Br. J. Anaesth. 2018, 120, 546–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acheson, A.; Conover, J.C.; Fandl, J.P.; DeChiara, T.M.; Russell, M.; Thadani, A.; Squinto, S.P.; Yancopoulos, G.D.; Lindsay, R.M. A BDNF autocrine loop in adult sensory neurons prevents cell death. Nature 1995, 374, 450–453. [Google Scholar] [CrossRef]
- Charych, E.I.; Akum, B.F.; Goldberg, J.S.; Jornsten, R.J.; Rongo, C.; Zheng, J.Q.; Firestein, B.L. Activity-independent regulation of dendrite patterning by postsynaptic density protein PSD-95. J. Neurosci. 2006, 26, 10164–10176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, C.; Dash, H.H. Procedural sedation and analgesia in pediatric patients. J. Pediatr. Neurosci. 2014, 9, 1–6. [Google Scholar]
- Penzes, P.; Cahill, M.E.; Jones, K.A.; VanLeeuwen, J.E.; Woolfrey, K.M. Dendritic spine pathology in neuropsychiatric disorders. Nat. Neurosci. 2011, 14, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.Y.; Zou, D.J.; Rajan, I.; Cline, H. Dendritic dynamics in vivo change during neuronal maturation. J. Neurosci. 1999, 19, 4472–4483. [Google Scholar] [CrossRef] [Green Version]
- McAllister, A.K. Cellular and molecular mechanisms of dendrite growth. Cereb. Cortex 2000, 10, 963–973. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, W.E.; Moser, H.W. Dendritic anomalies in disorders associated with mental retardation. Cereb. Cortex 2000, 10, 981–991. [Google Scholar] [CrossRef]
- Purpura, D.P. Dendritic differentiation in human cerebral cortex: Normal and aberrant developmental patterns. Adv. Neurol. 1975, 12, 91–134. [Google Scholar]
- Metz, A.E.; Yau, H.J.; Centeno, M.V.; Apkarian, A.V.; Martina, M. Morphological and functional reorganization of rat medial prefrontal cortex in neuropathic pain. Proc. Natl. Acad. Sci. USA 2009, 106, 2423–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Soltanzadeh Zarandi, S.; Sood, A.; Paddy, M.R.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef] [PubMed]
- Lepack, A.E.; Fuchikami, M.; Dwyer, J.M.; Banasr, M.; Duman, R.S. BDNF release is required for the behavioral actions of ketamine. Int. J. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, C.G.; Adams, T.G.; Kelmendi, B.; Esterlis, I.; Sanacora, G.; Krystal, J.H. Ketamine’s Mechanism of Action: A Path to Rapid-Acting Antidepressants. Depress. Anxiety 2016, 33, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Parihar, V.K.; Limoli, C.L. Cranial irradiation compromises neuronal architecture in the hippocampus. Proc. Natl. Acad. Sci. USA 2013, 110, 12822–12827. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Firestein, B.L. RhoA regulates dendrite branching in hippocampal neurons by decreasing cypin protein levels. J. Neurosci. 2007, 27, 8378–8386. [Google Scholar] [CrossRef] [Green Version]
- Arikkath, J. Molecular mechanisms of dendrite morphogenesis. Front. Cell. Neurosci. 2012, 6, 61. [Google Scholar] [CrossRef] [Green Version]
- Leemhuis, J.; Boutillier, S.; Barth, H.; Feuerstein, T.J.; Brock, C.; Nurnberg, B.; Aktories, K.; Meyer, D.K. Rho GTPases and phosphoinositide 3-kinase organize formation of branched dendrites. J. Biol. Chem. 2004, 279, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Luo, L. Actin cytoskeleton regulation in neuronal morphogenesis and structural plasticity. Annu. Rev. Cell Dev. Biol. 2002, 18, 601–635. [Google Scholar] [CrossRef] [Green Version]
- McAllister, A.K.; Lo, D.C.; Katz, L.C. Neurotrophins regulate dendritic growth in developing visual cortex. Neuron 1995, 15, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Bramham, C.R.; Panja, D. BDNF regulation of synaptic structure, function, and plasticity. Neuropharmacology 2014, 76(Pt. C), 601–602. [Google Scholar] [CrossRef]
- Tongiorgi, E.; Baj, G. Functions and mechanisms of BDNF mRNA trafficking. Novartis Found. Symp. 2008, 289, 136–147, discussion 147–151, 193–135. [Google Scholar]
- Wang, M.; Li, D.; Yun, D.; Zhuang, Y.; Repunte-Canonigo, V.; Sanna, P.P.; Behnisch, T. Translation of BDNF-gene transcripts with short 3’ UTR in hippocampal CA1 neurons improves memory formation and enhances synaptic plasticity-relevant signaling pathways. Neurobiol. Learn. Mem. 2017, 138, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Ash, R.T.; Baker, S.A.; Suter, B.; Ferguson, A.; Park, J.; Rudy, J.; Torsky, S.P.; Chao, H.T.; Zoghbi, H.Y.; et al. Dendritic arborization and spine dynamics are abnormal in the mouse model of MECP2 duplication syndrome. J. Neurosci. 2013, 33, 19518–19533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horch, H.W.; Katz, L.C. BDNF release from single cells elicits local dendritic growth in nearby neurons. Nat. Neurosci. 2002, 5, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hu, Y.M.; Zhou, Z.Q.; Zhang, G.F.; Yang, J.J. Acute administration of ketamine in rats increases hippocampal BDNF and mTOR levels during forced swimming test. Upsala J. Med. Sci. 2013, 118, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, Y.; Yang, M.; Kang, S.; Lee, S.; Kim, J.; Park, S.; Kim, J.S.; Jo, S.K.; Jung, U.; Shin, T.; et al. Cranial irradiation regulates CREB-BDNF signaling and variant BDNF transcript levels in the mouse hippocampus. Neurobiol. Learn. Mem. 2015, 121, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, M.V.; Melo, C.V.; Pereira, D.B.; Carvalho, R.F.; Carvalho, A.L.; Duarte, C.B. BDNF regulates the expression and traffic of NMDA receptors in cultured hippocampal neurons. Mol. Cell. Neurosci. 2007, 35, 208–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Levy, J.M.; Hou, A.; Winters, C.; Azzam, R.; Sousa, A.A.; Leapman, R.D.; Nicoll, R.A.; Reese, T.S. PSD-95 family MAGUKs are essential for anchoring AMPA and NMDA receptor complexes at the postsynaptic density. Proc. Natl. Acad. Sci. USA 2015, 112, E6983–E6992. [Google Scholar] [CrossRef] [Green Version]
- Kempf, S.J.; Casciati, A.; Buratovic, S.; Janik, D.; von Toerne, C.; Ueffing, M.; Neff, F.; Moertl, S.; Stenerlow, B.; Saran, A.; et al. The cognitive defects of neonatally irradiated mice are accompanied by changed synaptic plasticity, adult neurogenesis and neuroinflammation. Mol. Neurodegener. 2014, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Kempf, S.J.; Sepe, S.; von Toerne, C.; Janik, D.; Neff, F.; Hauck, S.M.; Atkinson, M.J.; Mastroberardino, P.G.; Tapio, S. Neonatal Irradiation Leads to Persistent Proteome Alterations Involved in Synaptic Plasticity in the Mouse Hippocampus and Cortex. J. Proteome Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lisek, M.; Ferenc, B.; Studzian, M.; Pulaski, L.; Guo, F.; Zylinska, L.; Boczek, T. Glutamate Deregulation in Ketamine-Induced Psychosis-A Potential Role of PSD95, NMDA Receptor and PMCA Interaction. Front. Cell. Neurosci. 2017, 11, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beurel, E.; Grieco, S.F.; Amadei, C.; Downey, K.; Jope, R.S. Ketamine-induced inhibition of glycogen synthase kinase-3 contributes to the augmentation of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptor signaling. Bipolar Disord. 2016, 18, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Buratovic, S.; Stenerlow, B.; Fredriksson, A.; Sundell-Bergman, S.; Viberg, H.; Eriksson, P. Neonatal exposure to a moderate dose of ionizing radiation causes behavioural defects and altered levels of tau protein in mice. Neurotoxicology 2014, 45, 48–55. [Google Scholar] [CrossRef]
- Eriksson, P.; Buratovic, S.; Fredriksson, A.; Stenerlow, B.; Sundell-Bergman, S. Neonatal exposure to whole body ionizing radiation induces adult neurobehavioural defects: Critical period, dose--response effects and strain and sex comparison. Behav. Brain Res. 2016, 304, 11–19. [Google Scholar] [CrossRef]
- Kempf, S.J.; Buratovic, S.; von Toerne, C.; Moertl, S.; Stenerlow, B.; Hauck, S.M.; Atkinson, M.J.; Eriksson, P.; Tapio, S. Ionising radiation immediately impairs synaptic plasticity-associated cytoskeletal signalling pathways in HT22 cells and in mouse brain: An in vitro/in vivo comparison study. PLoS ONE 2014, 9, e110464. [Google Scholar] [CrossRef] [Green Version]
- Schmal, Z.; Isermann, A.; Hladik, D.; von Toerne, C.; Tapio, S.; Rube, C.E. DNA damage accumulation during fractionated low-dose radiation compromises hippocampal neurogenesis. Radiother. Oncol. 2019, 137, 45–54. [Google Scholar] [CrossRef]
- Grosche, A.; Hauser, A.; Lepper, M.F.; Mayo, R.; von Toerne, C.; Merl-Pham, J.; Hauck, S.M. The Proteome of Native Adult Muller Glial Cells from Murine Retina. Mol. Cell. Proteom. 2016, 15, 462–480. [Google Scholar] [CrossRef] [Green Version]
- Dial, S.; Silver, P.; Bock, K.; Sagy, M. Pediatric sedation for procedures titrated to a desired degree of immobility results in unpredictable depth of sedation. Pediatr. Emerg. Care 2001, 17, 414–420. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Symbol | Entrez Gene Name | Fold-Change | Biological Function | GO Number | ||
|---|---|---|---|---|---|---|
| 100 mGy Ket | 200 mGy Ket | |||||
| 1 | ABHD10 | abhydrolase domain containing 10 | −2.014 | −1.602 | glucuronoside catabolic process | GO:0019391 |
| 2 | ACAN | aggrecan | −2.742 | −3.063 | negative regulation of cell migration | GO:0030336 |
| 3 | ADAM11 | ADAM metallopeptidase domain 11 | −1.344 | −1.622 | proteolysis | GO:0006508 |
| 4 | ADAM23 | ADAM metallopeptidase domain 23 | −1.325 | −1.409 | proteolysis | GO:0006508 |
| 5 | ARF6 | ADP ribosylation factor 6 | −1.432 | −1.365 | regulation of dendritic spine development | GO:0060998 |
| 6 | ARMC1 | armadillo repeat containing 1 | −2.059 | −1.752 | metal ion transport | GO:0030001 |
| 7 | ARPC1A | actin related protein 2/3 complex subunit 1A | 1.358 | 1.515 | regulation of actin filament polymerization | GO:0030833 |
| 8 | ASPA | aspartoacylase | 1.406 | 1.575 | positive regulation of oligodendrocyte differentiation | GO:0048714 |
| 9 | BRSK2 | BR serine/threonine kinase 2 | −1.680 | −1.829 | neuron differentiation | GO:0030182 |
| 10 | CBR3 | carbonyl reductase 3 | −1.478 | −1.577 | cognition | GO:0050890 |
| 11 | CDC42 | cell division cycle 42 | −1.588 | −1.474 | modification of synaptic structure | GO:0099563 |
| 12 | CRK | CRK proto-oncogene. adaptor protein | −1.948 | −2.240 | dendrite development | GO:0016358 |
| 13 | DNAJC6 | DnaJ heat shock protein family (Hsp40) member C6 | −1.381 | −1.484 | synaptic vesicle uncoating | GO:0016191 |
| 14 | DYNLL2 | dynein light chain LC8-type 2 | −1.316 | −1.329 | microtubule-based process | GO:0007017 |
| 15 | ELMO2 | engulfment and cell motility 2 | −2.612 | −2.402 | cytoskeleton organization | GO:0007010 |
| 16 | FBXO2 | F-box protein 2 | −1.503 | −1.307 | regulation of protein ubiquitination | GO:0031396 |
| 17 | GDPD1 | glycerophosphodiester phosphodiesterase domain containing 1 | −1.286 | −1.293 | N-acylethanolamine metabolic process | GO:0070291 |
| 18 | GGT7 | gamma-glutamyltransferase 7 | −2.601 | −4.805 | regulation of response to oxidative stress | GO:1902883 |
| 19 | GUK1 | guanylate kinase 1 | −1.496 | −1.412 | ATP metabolic process | GO:0046034 |
| 20 | HIST1H2BD | histone cluster 1 H2B family member d | 1.562 | 1.515 | protein ubiquitination | GO:0016567 |
| 21 | HNRNPUL1 | heterogeneous nuclear ribonucleoprotein U like 1 | −3.209 | −1.488 | RNA processing | GO:0006396 |
| 22 | HTT | huntingtin | −1.862 | −1.686 | learning or memory | GO:0007611 |
| 23 | IPO5 | importin 5 | −1.568 | −1.478 | protein import into nucleus | GO:0006606 |
| 24 | MICU3 | mitochondrial calcium uptake family member 3 | −1.285 | −1.484 | mitochondrial calcium ion transmembrane transport | GO:0006851 |
| 25 | NACA | nascent polypeptide associated complex subunit alpha | −1.297 | −1.290 | positive regulation of nucleic acid-templated transcription | GO:1903508 |
| 26 | NDRG2 | NDRG family member 2 | −1.295 | −1.326 | nervous system development | GO:0001818 |
| 27 | NIF3L1 | NGG1 interacting factor 3 like 1 | −1.649 | −1.671 | neuron differentiation | GO:0030182 |
| 28 | NRP1 | neuropilin 1 | −1.935 | −2.487 | axon guidance | GO:0007411 |
| 29 | OCIAD1 | OCIA domain containing 1 | 1.529 | 1.818 | regulation of stem cell differentiation | GO:2000736 |
| 30 | PAK3 | p21 (RAC1) activated kinase 3 | −2.309 | −2.063 | dendritic spine development | GO:0060996 |
| 31 | PCDH1 | protocadherin 1 | −1.874 | −3.052 | cell adhesion | GO:0007155 |
| 32 | PFDN6 | prefoldin subunit 6 | −2.242 | −1.669 | protein folding | GO:0006457 |
| 33 | PIP5K1C | phosphatidylinositol-4-phosphate 5-kinase type 1 gamma | −1.286 | −1.306 | axonogenesis | GO:0007409 |
| 34 | PRKAR2A | protein kinase cAMP-dependent type II regulatory subunit alpha | −1.292 | −1.351 | modulation of chemical synaptic transmission | GO:0050804 |
| 35 | PTGES3 | prostaglandin E synthase 3 | −1.543 | −1.780 | prostaglandin biosynthetic process | GO:0001516 |
| 36 | PTPRS | protein tyrosine phosphatase. receptor type S | −1.545 | −1.301 | hippocampus development | GO:0021766 |
| 37 | RAB1A | RAB1A. member RAS oncogene family | −1.302 | −1.374 | intracellular protein transport | GO:0006886 |
| 38 | RAB5C | RAB5C. member RAS oncogene family | −1.323 | −1.423 | intracellular protein transport | GO:0006886 |
| 39 | RABL6 | RAB. member RAS oncogene family like 6 | 1.737 | 1.548 | intracellular protein transport | GO:0006886 |
| 40 | RIMBP2 | RIMS binding protein 2 | −1.385 | −1.856 | neuromuscular synaptic transmission | GO:0007274 |
| 41 | RPLP2 | ribosomal protein lateral stalk subunit P2 | −1.543 | −1.508 | translational elongation | GO:0006414 |
| 42 | SEC24C | SEC24 homolog C. COPII coat complex component | −1.342 | −1.390 | vesicle-mediated transport | GO:0016192 |
| 43 | SLC1A4 | solute carrier family 1 member 4 | −1.414 | −1.344 | cognition | GO:0050890 |
| 44 | SNCA | synuclein alpha | −1.432 | −1.421 | synaptic transmission | GO:0001963 |
| 45 | STX7 | syntaxin 7 | −1.324 | −1.298 | vesicle-mediated transport | GO:0016192 |
| 46 | SUCLG1 | succinate-CoA ligase alpha subunit | 1.342 | 1.356 | succinyl-CoA metabolic process | GO:0006104 |
| 47 | TIMM13 | translocase of inner mitochondrial membrane 13 | −1.845 | −1.487 | protein insertion into mitochondrial inner membrane | GO:0045039 |
| 48 | TPD52 | tumor protein D52 | −1.468 | −1.384 | positive regulation of cell population proliferation | GO:0008284 |
| 49 | TRAPPC10 | trafficking protein particle complex 10 | −1.392 | −1.664 | vesicle-mediated transport | GO:0016192 |
| 50 | TRIO | trio Rho guanine nucleotide exchange factor | −1.562 | −1.369 | G-protein-coupled receptor signaling pathway | GO:0007186 |
| 51 | TUBA8 | tubulin alpha 8 | −1.472 | −1.343 | microtubule cytoskeleton organization | GO:0000226 |
| 52 | UBXN6 | UBX domain protein 6 | −1.360 | −1.419 | macroautophagy | GO:0016236 |
| 53 | UCHL3 | ubiquitin C-terminal hydrolase L3 | −1.413 | −1.427 | adult walking behavior | GO:0007628 |
| 54 | VBP1 | VHL binding protein 1 | −1.747 | −1.809 | protein folding | GO:0006457 |
| 55 | WASF3 | WAS protein family member 3 | −1.904 | −1.429 | actin cytoskeleton organization | GO:0030036 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hladik, D.; Buratovic, S.; Von Toerne, C.; Azimzadeh, O.; Subedi, P.; Philipp, J.; Winkler, S.; Feuchtinger, A.; Samson, E.; Hauck, S.M.; et al. Combined Treatment with Low-Dose Ionizing Radiation and Ketamine Induces Adverse Changes in CA1 Neuronal Structure in Male Murine Hippocampi. Int. J. Mol. Sci. 2019, 20, 6103. https://doi.org/10.3390/ijms20236103
Hladik D, Buratovic S, Von Toerne C, Azimzadeh O, Subedi P, Philipp J, Winkler S, Feuchtinger A, Samson E, Hauck SM, et al. Combined Treatment with Low-Dose Ionizing Radiation and Ketamine Induces Adverse Changes in CA1 Neuronal Structure in Male Murine Hippocampi. International Journal of Molecular Sciences. 2019; 20(23):6103. https://doi.org/10.3390/ijms20236103
Chicago/Turabian StyleHladik, Daniela, Sonja Buratovic, Christine Von Toerne, Omid Azimzadeh, Prabal Subedi, Jos Philipp, Stefanie Winkler, Annette Feuchtinger, Elenore Samson, Stefanie M. Hauck, and et al. 2019. "Combined Treatment with Low-Dose Ionizing Radiation and Ketamine Induces Adverse Changes in CA1 Neuronal Structure in Male Murine Hippocampi" International Journal of Molecular Sciences 20, no. 23: 6103. https://doi.org/10.3390/ijms20236103