Combined Effect of Cadmium and Lead on Durum Wheat


 , , , ,
, , , ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
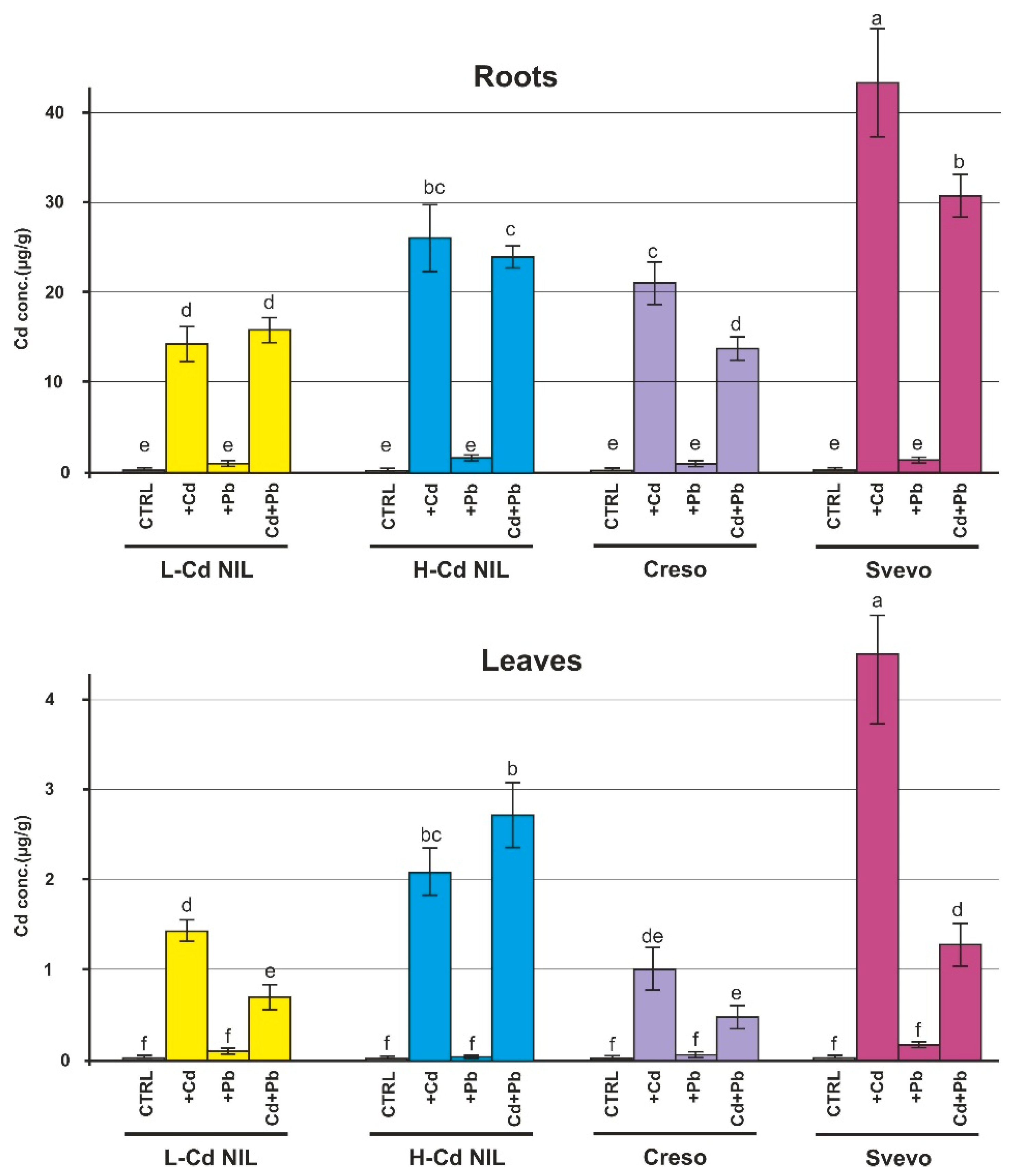
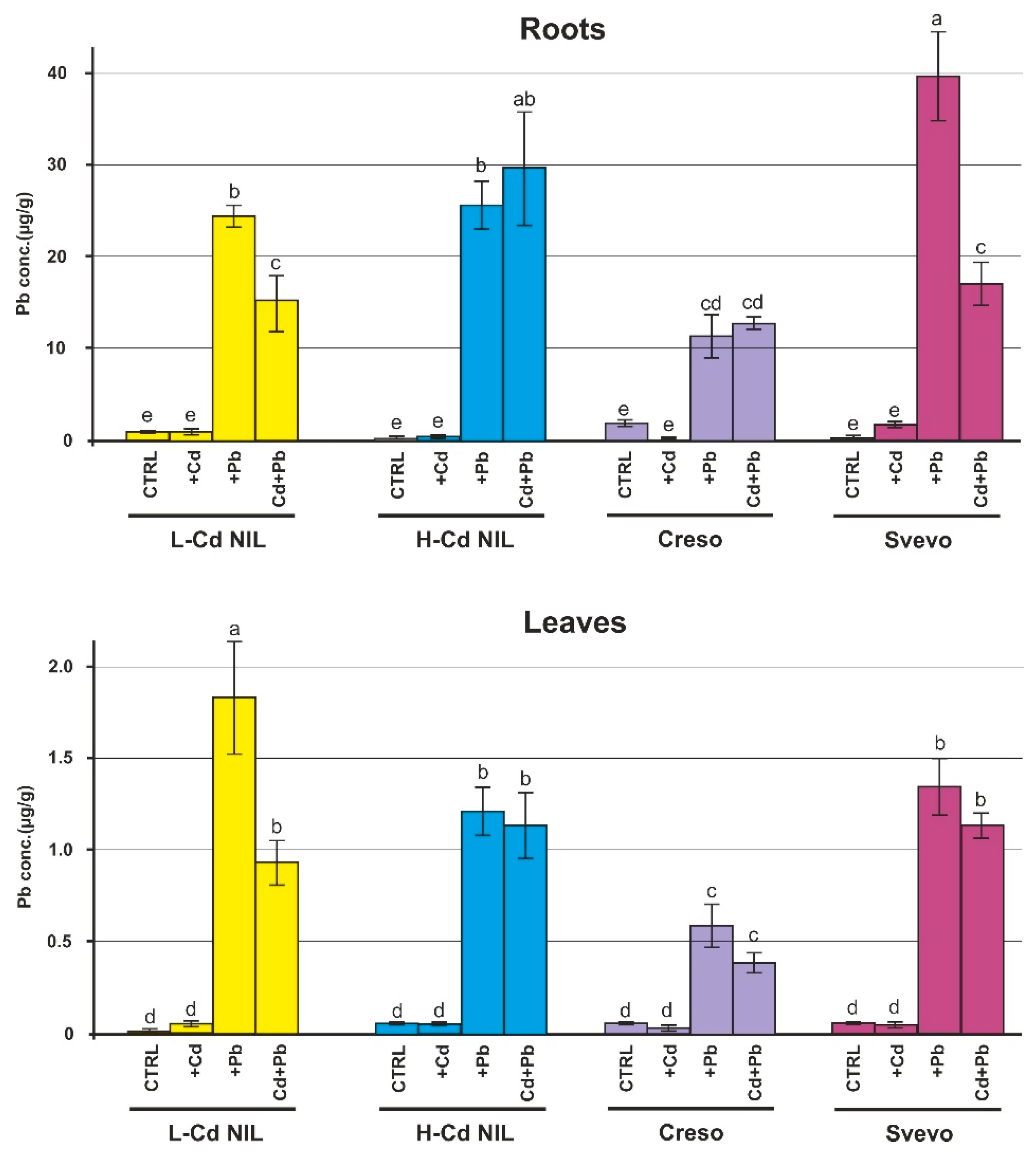
2.1. Levels of Cd and Pb in Root and Leaf of Wheat Plants
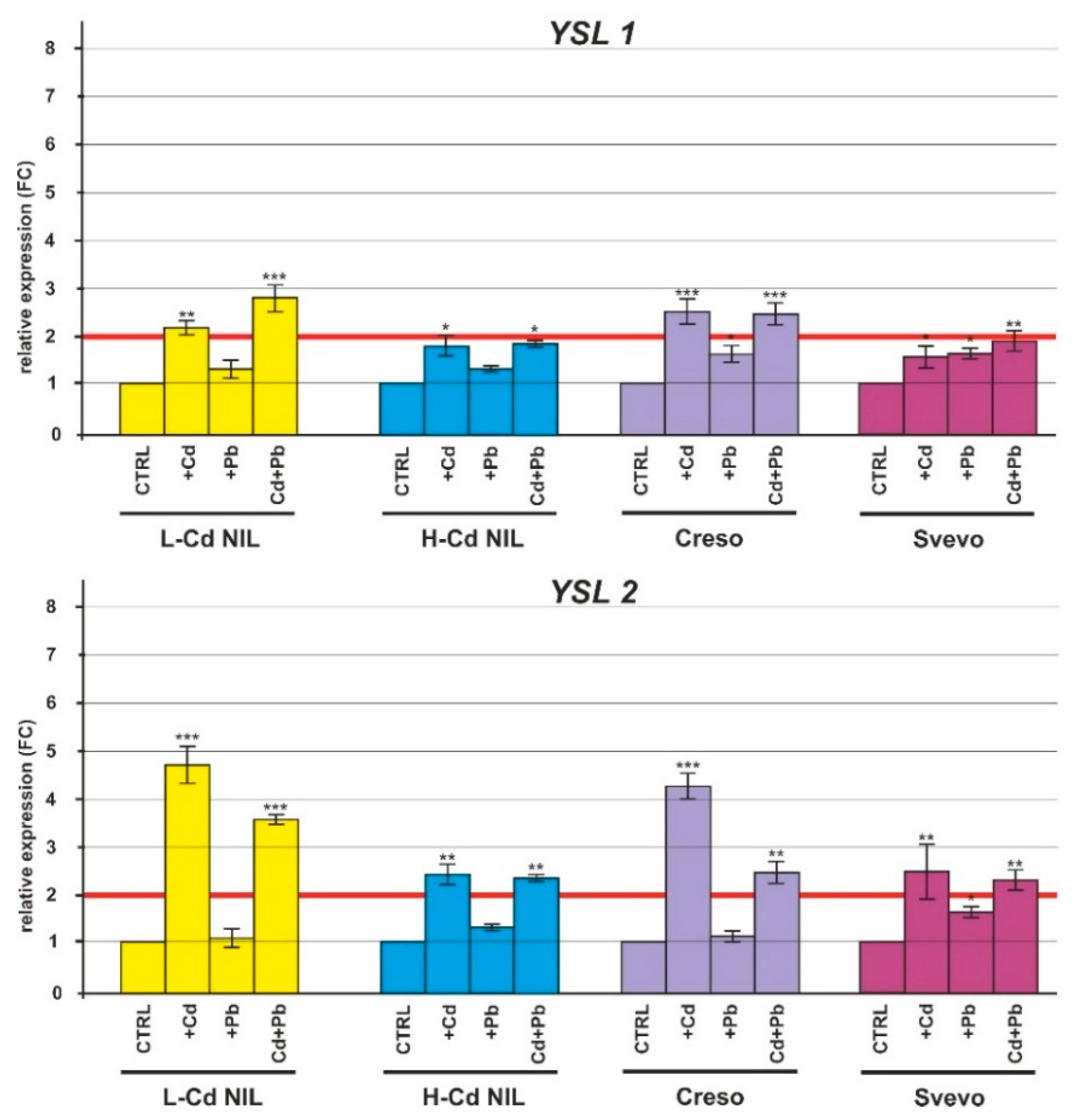
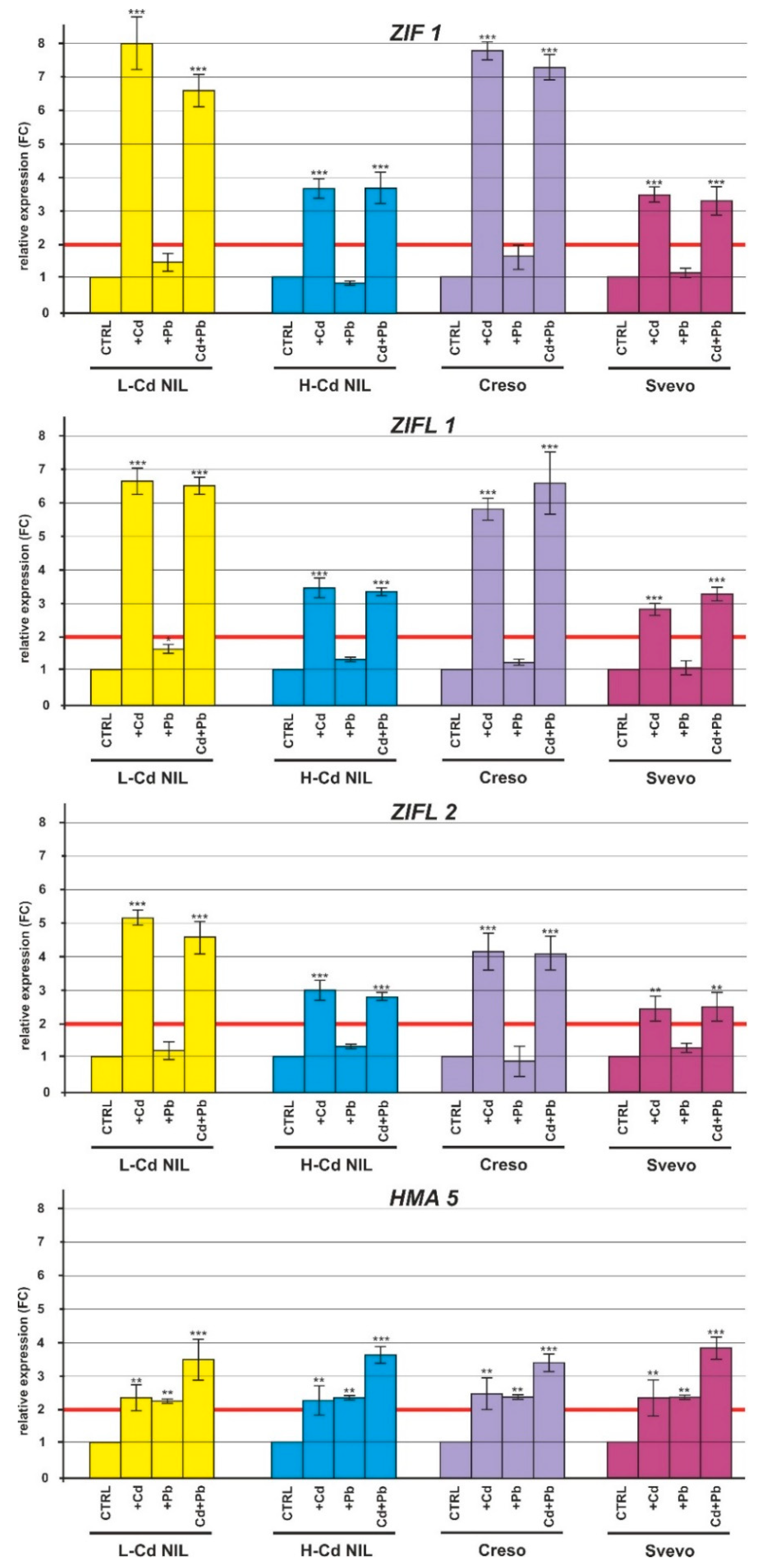
2.2. Gene Expression in Response to Cd and Pb
2.2.1. Expression of the Transcription Factors Basic Helix-Loop-Helix (bHLH) and WRKY33
2.2.2. Expression of HMs Transporters
2.2.3. Expression of the Nicotianamine Synthase Genes (NAS) and Nicotianamine Aminotransferase (NAAT)
3. Discussion
4. Materials and Methods
4.1. Genetic Materials
4.2. Experimental Design
4.3. Inductively Coupled Plasma Mass Spectrometry (ICP-MS) Analysis
4.4. Total RNA Isolation, cDNA Synthesis and qPCR Analysis of Gene Expression
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Romic, M.; Romic, D. Heavy metals distribution in agricultural topsoils in urban area. Environ. Geol. 2003, 43, 795–805. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Su, M.; Chen, Y.; Lin, F.; Luo, D.; Gao, S. Transfer characteristics of cadmium and lead from soil to the edible parts of six vegetable species in southeastern China. Environ. Pollut. 2006, 144, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, A.A.; Abduli, M.A.; Karbassi, A.R.; Nasrabadi, T.; Khajeh, M. Evaluating the Effects of Fertilizers on Bioavailable Metallic Pollution of soils, Case study of Sistan farms, Iran. Int. J. Environ. Res. 2012, 6, 565–570. [Google Scholar]
- Codex Alimentarius. General Standard for Contaminants and Toxins in Food and Feed. CXS 193-1995 (Amended 2019). Available online: http://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B193-1995%252FCXS_193e.pdf (accessed on 12 November 2019).
- Hasan, S.A.; Fariduddin, Q.; Ali, B.; Hayat, S.; Ahmad, A. Cadmium: Toxicity and tolerance in plants. J. Environ. Biol. 2009, 30, 165–174. [Google Scholar]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead Uptake, Toxicity, and Detoxification in Plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar]
- Florijn, P.J.; Van Beusichem, M.L. Uptake and distribution of cadmium in maze inbred lines. Plant Soil 1993, 150, 25–32. [Google Scholar] [CrossRef]
- Li, Y.M.; Channey, L.R.; Scheiter, A.A. Genotypic variation in kernel cadmium concentration in sunflower germplasm under varying soil conditions. Crop Sci. 1995, 35, 137–141. [Google Scholar] [CrossRef]
- DalCorso, G.; Fasani, E.; Manara, A.; Visioli, G.; Furini, A. Heavy Metal Pollutions: State of the Art and Innovation in Phytoremediation. Int. J. Mol. Sci. 2019, 20, 3412. [Google Scholar] [CrossRef]
- Backhaus, T.; Faust, M. Predictive environmental risk assessment of chemical mixtures: A conceptual framework. Environ. Sci. Technol. 2012, 4, 2564–2573. [Google Scholar] [CrossRef]
- Vergine, M.; Aprile, A.; Sabella, E.; Genga, A.; Siciliano, M.; Rampino, P.; Lenucci, M.S.; Luvisi, A.; De Bellis, L. Cadmium Concentration in Grains of Durum Wheat (Triticum turgidum L. subsp. durum). J. Agr. Food Chem. 2017, 65, 6240–6246. [Google Scholar] [CrossRef] [PubMed]
- Arduini, I.; Masoni, A.; Mariotti, M.; Pampana, S.; Ercoli, L. Cadmium uptake and translocation in durum wheat varieties differing in grain-Cd accumulation. Plant Soil Environ. 2014, 60, 43–49. [Google Scholar] [CrossRef]
- Bravin, M.L.; Le Merrer, B.; Denaix, L.; Schneider, A.; Hinsinger, P. Copper uptake kinetics in hydroponically-grown durum wheat (Triticum turgidum durum L.) as compared with soil’s ability to supply copper. Plant Soil 2010, 331, 91–104. [Google Scholar] [CrossRef]
- Shi, G.L.; Li, D.J.; Wang, Y.F.; Liu, C.H.; Hu, Z.B.; Lou, L.Q.; Rengel, Z.; Cai, Q.S. Accumulation and distribution of arsenic and cadmium in winter wheat (Triticum aestivum L.) at different developmental stages. Sci. Total Environ. 2019, 667, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Dalir, N.; Khoshgoftarmanesh, A.H. Symplastic and apoplastic uptake and root to shoot translocation of nickel in wheat as affected by exogenous amino acids. J. Plant Physiol. 2014, 171, 531–536. [Google Scholar] [CrossRef]
- Frost, H.L.; Ketchum, L.H.J. Trace metal concentration in durum wheat from application of sewage sludge and commercial fertilizer. Adv. Environ. Res. 2000, 4, 347–355. [Google Scholar] [CrossRef]
- Shafiq, S.; Zeb, Q.; Ali, A.; Sajjad, Y.; Nazir, R.; Widemann, E.; Liu, L. Lead, Cadmium and Zinc Phytotoxicity Alter DNA Methylation Levels to Confer Heavy Metal Tolerance in Wheat. Int. J. Mol. Sci. 2019, 20, 4676. [Google Scholar] [CrossRef]
- Clarke, J.M.; Leisle, D.; DePauw, R.M.; Thiessen, L.L. The registration of genetic stocks: Registration of five pairs of near-isogenic lines for cadmium concentration. Crop Sci. 1995, 37, 297. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Cattivelli, L.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef]
- Lakhdar, A.; Iannelli, M.A.; Debez, A.; Massacci, A.; Jedidi, N.; Abdelly, C. Effect of municipal solid waste compost and sewage sludge use on wheat (Triticum durum): Growth, heavy metal accumulation, and antioxidant activity. J. Sci. Food Agric. 2010, 90, 965–971. [Google Scholar] [CrossRef]
- Aprile, A.; Sabella, E.; Vergine, M.; Genga, A.; Siciliano, M.; Nutricati, E.; Rampino, P.; De Pascali, M.; Luvisi, A.; Miceli, A.; et al. Activation of a gene network in durum wheat roots exposed to cadmium. BMC Plant Biol. 2018, 18, 238. [Google Scholar] [CrossRef] [PubMed]
- Haydon, M.J.; Cobbett, C.S. Transporters of ligands for essential metal ions in plants. New Phytol. 2007, 174, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Videaa, J.R.; Gardea-Torresdeya, J.L.; Gomezc, E.; Tiemanna, K.J.; Parsonsa, J.G.; Carrillod, G. Effect of mixed cadmium, copper, nickel and zinc at different pHs upon alfalfa growth and heavy metal uptake. Environ. Pollut. 2002, 119, 291–301. [Google Scholar] [CrossRef]
- Xie, L.; Hao, P.; Cheng, Y.; Ahmed, I.M.; Cao, F. Effect of combined application of lead, cadmium, chromium and copper on grain, leaf and stem heavy metal contents at different growth stages in rice. Ecotoxicol. Environ. Saf. 2018, 162, 71–76. [Google Scholar] [CrossRef]
- Zeng, F.; Mao, Y.; Cheng, W.; Wu, F.; Zhang, G. Genotypic and environmental variation in chromium, cadmium and lead concentrations in rice. Environ. Pollut. 2008, 153, 309–314. [Google Scholar] [CrossRef]
- An, Y.J. Soil ecotoxicity assessment using cadmium sensitive plants. Environ. Pollut. 2004, 127, 21–26. [Google Scholar] [CrossRef]
- An, Y.J.; Kimb, Y.M.; Kwonb, T.I.; Jeong, S.-W. Combined effect of copper, cadmium, and lead upon Cucumis sativus growth and bioaccumulation. Sci. Total Environ. 2004, 326, 85–93. [Google Scholar] [CrossRef]
- Zaray, G.; Phuong, D.D.T.; Varga, I.; Kantor, T.; Cseh, E.; Fodor, F. Influences of lead contamination and complexing agents on the metal uptake of cucumber. Microchem. J. 1995, 51, 207–213. [Google Scholar] [CrossRef]
- Mohan, B.S.; Hosetti, B.B. Potential phytotoxicity of lead and cadmium to Lemna minor grown in sewage stabilization ponds. Environ. Pollut. 1997, 98, 233–238. [Google Scholar] [CrossRef]
- Hart, J.J.; Welch, R.M.; Norvell, W.A.; Kochian, L.V. Transport interactions between cadmium and zinc in roots of bread and durum wheat seedlings. Physiol. Plant. 2002, 116, 73–78. [Google Scholar] [CrossRef]
- Salsman, E.; Kumar, A.; AbuHammad, W.; Abbasabadi, A.O.; Dobrydina, M.; Chao, S.; Li, X.; Manthey, F.A.; Elias, E.M. Development and validation of molecular markers for grain cadmium in durum wheat. Mol. Breed. 2018, 38, 28. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculìk, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, J.; Li, B.; Ou, Y.; Jiang, W.; Liu, D.; Zou, J. Uptake and Accumulation of Cadmium and Relative Gene Expression in Roots of Cd-resistant Salix matsudana Koidz. Pol. J. Environ. Stud. 2016, 25, 2717–2723. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Fukuoka, H.; Arao, T.; Ohyama, A.; Nunome, T.; Miyatake, K.; Negoro, S. Gene expression analysis in cadmium-stressed roots of a low cadmium-accumulating solanaceous plant, Solanum torvum. J. Exp. Bot. 2010, 61, 423–437. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, Q.; Yu, J.; Yuan, M. The putative Arabidopsis zinc transporter ZTP29 is involved in the response to salt stress. Plant Mol. Biol. 2010, 73, 467–479. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, X.; Feng, Y.; Zhang, C.; Wang, C.; Zeng, J.; Huang, Z.; Kang, H.; Fan, X.; Sha, L.; et al. Physiological and transcriptome response to cadmium in cosmos (Cosmos bipinnatus Cav.) seedlings. Sci. Rep. 2017, 7, 14691. [Google Scholar] [CrossRef] [Green Version]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Jean, M.L.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.-H.; Chiecko, J.; Punshon, T.; Lanzirotti, A.; Lahner, B.; Salt, D.E.; Walker, E.L. Successful Reproduction Requires the Function of Arabidopsis YELLOW STRIPE-LIKE1 and YELLOW STRIPE-LIKE3 Metal-Nicotianamine Transporters in Both Vegetative and Reproductive Structure. Plant Physiol. 2010, 154, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Conte, S.S.; Chu, H.H.; Chan Rodriguez, D.; Punshon, T.; Vasques, K.A.; Salt, D.E.; Walker, E.L. Arabidopsis thaliana Yellow Stripe1- Like4 and Yellow Stripe1-Like6 localize to internal cellular membranes and are involved in metal ion homeostasis. Front. Plant Sci. 2013, 4, 283. [Google Scholar] [CrossRef] [Green Version]
- John, R.; Ahmad, P.; Gadgil, K.; Sharma, S. Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ. 2008, 54, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.E.; Mills, R.F. P(1B-)ATPases—An ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef]
- DalCorso, G.; Farinati, S.; Furini, A. Regulatory networks of cadmium stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Sheng, Y.; Yan, X.; Huang, Y.; Han, Y.; Zhang, C.; Ren, Y.; Fan, T.; Xiao, F.; Liu, Y.; Cao, S. The WRKY transcription factor, WRKY13, activates PDR8 expression to positively regulate cadmium tolerance in Arabidopsis. Plant Cell Environ. 2019, 42, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Long, T.A.; Tsukagoshi, H.; Busch, W.; Lahner, B.; Salt, D.E.; Benfey, P.N. The bHLH transcription factor POPEYE regulates response to Iron deficiency in Arabidopsis roots. Plant Cell 2010, 22, 2219–2236. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Rai, J.P.N. Phytochelatins: Peptides Involved in Heavy Metal Detoxification. Appl. Biochem. Biotechnol. 2010, 160, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Fernández, A.; Díaz-Benito, P.; Abadía, A.; López-Millán, A.-F.; Abadía, J. Metal species involved in long distance metal transport in plants. Front. Plant Sci. 2014, 5, 105. [Google Scholar] [CrossRef] [Green Version]
- Beasley, J.T.; Bonneau, J.P.; Johnson, A.A.T. Characterisation of the nicotianamine aminotransferase and deoxymugineic acid synthase genes essential to Strategy II iron uptake in bread wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0177061. [Google Scholar] [CrossRef]
- Harris, N.S.; Taylor, G.J. Cadmium uptake and partitioning in durum wheat during grain filling. BMC Plant Biol. 2013, 13, 103. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Wang, X.-R.; Ding, S.-M.; Yuan, X.-F. Effects of Interaction Between Cadmium and Plumbum on Phytochelatins and Glutathione Production in Wheat (Triticum aestivum L.). J. Integr. Plant Biol. 2015, 47, 435–442. [Google Scholar] [CrossRef]
- Durante, C.; Bertacchini, L.; Cocchi, M.; Manzini, D.; Marchetti, A.; Rossi, M.C.; Sighinolfi, S.; Tassi, L. Development of 87Sr/86Sr maps as targeted strategy to support wine quality. Food Chem. 2018, 255, 139–146. [Google Scholar] [CrossRef]
- Marè, C.; Aprile, A.; Roncaglia, E.; Tocci, E.; Corino, L.G.; De Bellis, L.; Cattivelli, L. Rootstock and soil induce transcriptome modulation of phenylpropanoid pathway in grape leaves. J. Plant Interact. 2014, 8, 334–349. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aprile, A.; Sabella, E.; Francia, E.; Milc, J.; Ronga, D.; Pecchioni, N.; Ferrari, E.; Luvisi, A.; Vergine, M.; De Bellis, L. Combined Effect of Cadmium and Lead on Durum Wheat. Int. J. Mol. Sci. 2019, 20, 5891. https://doi.org/10.3390/ijms20235891
Aprile A, Sabella E, Francia E, Milc J, Ronga D, Pecchioni N, Ferrari E, Luvisi A, Vergine M, De Bellis L. Combined Effect of Cadmium and Lead on Durum Wheat. International Journal of Molecular Sciences. 2019; 20(23):5891. https://doi.org/10.3390/ijms20235891
Chicago/Turabian StyleAprile, Alessio, Erika Sabella, Enrico Francia, Justyna Milc, Domenico Ronga, Nicola Pecchioni, Erika Ferrari, Andrea Luvisi, Marzia Vergine, and Luigi De Bellis. 2019. "Combined Effect of Cadmium and Lead on Durum Wheat" International Journal of Molecular Sciences 20, no. 23: 5891. https://doi.org/10.3390/ijms20235891