Soyuretox, an Intrinsically Disordered Polypeptide Derived from Soybean (Glycine Max) Ubiquitous Urease with Potential Use as a Biopesticide

 , , , , and
, , , , and
Abstract
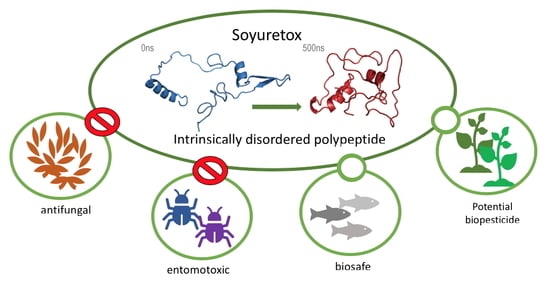
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Cloning, Expression, and Purification of Soyuretox
2.2. CD and NMR Spectroscopic Studies
2.3. Molecular Modeling and Dynamics Simulation
2.4. Antifungal Activity of Soyuretox
2.5. Entomotoxic Activity of Soyuretox
2.6. Biological Activity of Soyuretox against Zebrafish
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression, and Purification of Soyuretox
4.2. CD and NMR Spectroscopic Studies
4.3. Molecular Modeling and Dynamics Simulation
4.4. Antifungal Activity of Soyuretox
4.5. Entomotoxic Activity of Soyuretox
4.6. Biological Activity of Soyuretox against Zebrafish
4.7. Statistical Analysis of Biological Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CNTX | Canatoxin |
| JBU | Jack bean urease |
| NMR | Nuclear Magnetic Resonance |
| uSBU | Soybean ubiquitous urease |
| MD | Molecular Dynamics |
| CD | Circular Dichroism |
| SDS | Sodium Dodecyl Sulfate |
| CMC | Critical Micellar Concentration |
| ROS | Reactive Oxygen Species |
| TCEP | Tris(2-carboxyethyl)phosphine |
| dpf | Days post-fertilization |
| LB | Luria-Bertani broth |
| IPTG | Isopropyl β-D-1-thiogalactopyranoside |
| PCR | Polymerase Chain Reaction |
| CFU | Colony Forming Units |
| DMSO | Dimethyl sulfoxide |
References
- Maroney, M.J.; Ciurli, S. Nonredox nickel enzymes. Chem. Rev. 2014, 114, 4206–4228. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, L.; Musiani, F.; Ciurli, S. Urease. In The Biological Chemistry of Nickel; Zamble, D., Rowińska-Żyrek, M., Kozłowski, H., Eds.; Royal Society of Chemistry: London, UK, 2017; pp. 60–97. [Google Scholar]
- Zambelli, B.; Musiani, F.; Benini, S.; Ciurli, S. Chemistry of Ni2+ in urease: Sensing, trafficking, and catalysis. Acc. Chem. Res. 2011, 44, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, B. Ureases I. Functional, catalytic and kinetic properties: A review. J. Mol. Catal. B-Enzym. 2009, 59, 9–21. [Google Scholar] [CrossRef]
- Ligabue-Braun, R.; Andreis, F.C.; Verli, H.; Carlini, C.R. 3-to-1: Unraveling structural transitions in ureases. Naturwissenschaften 2013, 100, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Carlini, C.R.; Ligabue-Braun, R. Ureases as multifunctional toxic proteins: A review. Toxicon 2016, 110, 90–109. [Google Scholar] [CrossRef] [PubMed]
- Carlini, C.R.; Oliveira, A.E.A.; Azambuja, P.; Xavier, J.; Wells, M.A. Biological effects of canatoxin in different insect models: Evidence for a proteolytic activation of the toxin by insect cathepsinlike enzymes. J. Econ. Entomol. 1997, 90, 340–348. [Google Scholar] [CrossRef]
- Follmer, C.; Wassermann, G.E.; Carlini, C.R. Separation of jack bean (Canavalia ensiformis) urease isoforms by immobilized metal affinity chromatography and characterization of insecticidal properties unrelated to ureolytic activity. Plant Sci. 2004, 167, 241–246. [Google Scholar] [CrossRef]
- Staniscuaski, F.; Carlini, C.R. Plant ureases and related peptides: Understanding their entomotoxic properties. Toxins 2012, 4, 55–67. [Google Scholar] [CrossRef]
- Ferreira-DaSilva, C.T.; Gombarovits, M.E.C.; Masuda, H.; Oliveira, C.M.; Carlini, C.R. Proteolytic activation of canatoxin, a plant toxic protein, by insect cathepsin-like enzymes. Arch. Insect Biochem. 2000, 44, 162–171. [Google Scholar] [CrossRef]
- Piovesan, A.R.; Staniscuaski, F.; Marco-Salvadori, J.; Real-Guerra, R.; Defferrari, M.S.; Carlini, C.R. Stage-specific gut proteinases of the cotton stainer bug Dysdercus peruvianus: Role in the release of entomotoxic peptides from Canavalia ensiformis urease. Insect Biochem. Molec. 2008, 38, 1023–1032. [Google Scholar] [CrossRef]
- Defferrari, M.S.; Demartini, D.R.; Marcelino, T.B.; Pinto, P.M.; Carlini, C.R. Insecticidal effect of Canavalia ensiformis major urease on nymphs of the milkweed bug Oncopeltus fasciatus and characterization of digestive peptidases. Insect Biochem. Molec. 2011, 41, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Real-Guerra, R.; Carlini, C.R.; Staniscuaski, F. Role of lysine and acidic amino acid residues on the insecticidal activity of jackbean urease. Toxicon 2013, 71, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Mulinari, F.; Staniscuaski, F.; Bertholdo-Vargas, L.R.; Postal, M.; Oliveira-Neto, O.B.; Rigden, D.J.; Grossi-de-Sa, M.F.; Carlini, C.R. Jaburetox-2Ec: An insecticidal peptide derived from an isoform of urease from the plant Canavalia ensiformis. Peptides 2007, 28, 2042–2050. [Google Scholar] [CrossRef] [PubMed]
- Postal, M.; Martinelli, A.H.S.; Becker-Ritt, A.B.; Ligabue-Braun, R.; Demartini, D.R.; Ribeiro, S.F.F.; Pasquali, G.; Gomes, V.M.; Carlini, C.R. Antifungal properties of Canavalia ensiformis urease and derived peptides. Peptides 2012, 38, 22–32. [Google Scholar] [CrossRef]
- Stanisçuaski, F.; Ferreira-Dasilva, C.T.; Mulinari, F.; Pires-Alves, M.; Carlini, C.R. Insecticidal effects of canatoxin on the cotton stainer bug Dysdercus peruvianus (Hemiptera: Pyrrhocoridae). Toxicon 2005, 45, 753–760. [Google Scholar] [CrossRef]
- Kappaun, K.; Piovesan, A.R.; Carlini, C.R.; Ligabue-Braun, R. Ureases: Historical aspects, catalytic, and non-catalytic properties–A review. J. Adv. Res. 2018, 13, 3–17. [Google Scholar] [CrossRef]
- Stanisçuaski, F.; Te Brugge, V.; Carlini, C.R.; Orchard, I. In vitro effect of Canavalia ensiformis urease and the derived peptide Jaburetox-2Ec on Rhodnius prolixus Malpighian tubules. J. Insect Physiol. 2009, 55, 255–263. [Google Scholar] [CrossRef]
- Galvani, G.L.; Fruttero, L.L.; Coronel, M.F.; Nowicki, S.; Demartini, D.R.; Defferrari, M.S.; Postal, M.; Canavoso, L.E.; Carlini, C.R.; Settembrini, B.P. Effect of the urease-derived peptide Jaburetox on the central nervous system of Triatoma infestans (Insecta: Heteroptera). BBA-Gen. Subj. 2015, 1850, 255–262. [Google Scholar] [CrossRef]
- Fruttero, L.L.; Moyetta, N.R.; Uberti, A.F.; Grahl, M.V.C.; Lopes, F.C.; Broll, V.; Feder, D.; Carlini, C.R. Humoral and cellular immune responses induced by the urease-derived peptide Jaburetox in the model organism Rhodnius prolixus. Parasite Vector 2016, 9. [Google Scholar] [CrossRef]
- Lopes, F.C.; Dobrovolska, O.; Real-Guerra, R.; Broll, V.; Zambelli, B.; Musiani, F.; Uversky, V.N.; Carlini, C.R.; Ciurli, S. Pliable natural biocide: Jaburetox is an intrinsically disordered insecticidal and fungicidal polypeptide derived from jack bean urease. FEBS J. 2015, 282, 1043–1064. [Google Scholar] [CrossRef]
- Barros, P.R.; Stassen, H.; Freitas, M.S.; Carlini, C.R.; Nascimento, M.A.C.; Follmer, C. Membrane-disruptive properties of the bioinsecticide Jaburetox-2Ec: Implications to the mechanism of the action of insecticidal peptides derived from ureases. Bba-Proteins Proteom. 2009, 1794, 1848–1854. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, A.H.; Kappaun, K.; Ligabue-Braun, R.; Defferrari, M.S.; Piovesan, A.R.; Stanisçuaski, F.; Demartini, D.R.; Dal Belo, C.A.; Almeida, C.G.; Follmer, C.; et al. Structure-function studies on jaburetox, a recombinant insecticidal peptide derived from jack bean (Canavalia ensiformis) urease. Biochim. Biophys. Acta 2014, 1840, 935–944. [Google Scholar] [CrossRef]
- Micheletto, Y.M.S.; Moro, C.F.; Lopes, F.C.; Ligabue-Braun, R.; Martinelli, A.H.S.; Marques, C.M.; Schroder, A.P.; Carlini, C.R.; da Silveira, N.P. Interaction of jack bean (Canavalia ensiformis) urease and a derived peptide with lipid vesicles. Colloid Surf. B 2016, 145, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, A.H.S.; Lopes, F.C.; Broll, V.; Defferrari, M.S.; Ligabue-Braun, R.; Kappaun, K.; Tichota, D.M.; Fruttero, L.L.; Moyetta, N.R.; Demartini, D.R.; et al. Soybean ubiquitous urease with purification facilitator: An addition to the moonlighting studies toolbox. Process Biochem. 2017, 53, 245–258. [Google Scholar] [CrossRef]
- Broll, V.; Martinelli, A.H.S.; Lopes, F.C.; Fruttero, L.L.; Zambelli, B.; Salladini, E.; Dobrovolska, O.; Ciurli, S.; Carlini, C.R. Structural analysis of the interaction between Jaburetox, an intrinsically disordered protein, and membrane models. Colloid Surf. B 2017, 159, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, B.; Felli, I.C.; Gil-Caballero, S.; Hošek, T.; Kümmerle, R.; Piai, A.; Pierattelli, R.; Sólyom, Z. NMR methods for the study of instrinsically disordered proteins structure, dynamics, and interactions: General overview and practical guidelines. In Intrinsically Disordered Proteins Studied by NMR Spectroscopy; Felli, I.C., Pierattelli, R., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 49–122. [Google Scholar]
- Balasubramanian, A.; Ponnuraj, K. Crystal structure of the first plant urease from jack bean: 83 years of journey from its first crystal to molecular structure. J. Mol. Biol. 2010, 400, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Kriwacki, R.W.; Hengst, L.; Tennant, L.; Reed, S.I.; Wright, P.E. Structural studies of p21Waf1/Cip1/Sdi1 in the free and Cdk2-bound state: Conformational disorder mediates binding diversity. Proc. Natl. Acad. Sci. USA 1996, 93, 11504–11509. [Google Scholar] [CrossRef]
- Spolar, R.S.; Record, M.T. Coupling of local folding to site-specific binding of proteins to DNA. Science 1994, 263, 777–784. [Google Scholar] [CrossRef]
- Mello, E.O.; Ribeiro, S.F.F.; Carvalho, A.O.; Santos, I.S.; Da Cunha, M.; Santa-Catarina, C.; Gomes, V.M. Antifungal activity of PvD1 defensin involves plasma membrane permeabilization, inhibition of medium acidification, and induction of ROS in fungi cells. Curr. Microbiol. 2011, 62, 1209–1217. [Google Scholar] [CrossRef]
- Wang, K.R.; Dang, W.; Xie, J.Q.; Zhu, R.R.; Sun, M.Y.; Jia, F.J.; Zhao, Y.Y.; An, X.P.; Qiu, S.; Li, X.Y.; et al. Antimicrobial peptide protonectin disturbs the membrane integrity and induces ROS production in yeast cells. BBA Biomembr. 2015, 1848, 2365–2373. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.; Lee, D.G. The antimicrobial peptide arenicin-1 promotes generation of reactive oxygen species and induction of apoptosis. BBA-Gen. Subj. 2011, 1810, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Basso, V.; Garcia, A.; Tran, D.Q.; Schaal, J.B.; Tran, P.; Ngole, D.; Aqeel, Y.; Tongaonkar, P.; Ouellette, A.J.; Selsted, M.E. Fungicidal potency and mechanisms of θ-defensins against multidrug-resistant Candida species. Antimicrob. Agents Chemother. 2018, 62, e00111–e00118. [Google Scholar] [CrossRef] [PubMed]
- Becker-Ritt, A.B.; Carlini, C.R. Fungitoxic and insecticidal plant polypeptides. J. Pept. Sci 2012, 98, 367–384. [Google Scholar] [CrossRef]
- Salditt, T.; Li, C.H.; Spaar, A. Structure of antimicrobial peptides and lipid membranes probed by interface-sensitive X-ray scattering. BBA Biomembr. 2006, 1758, 1483–1498. [Google Scholar] [CrossRef] [PubMed]
- Follmer, C.; Real-Guerra, R.; Wasserman, G.E.; Olivera-Severo, D.; Carlini, C.R. Jackbean, soybean and Bacillus pasteurii ureases. Eur. J. Biochem. 2004, 271, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Siebel, A.M.; Piato, A.L.; Capiotti, K.M.; Seibt, K.J.; Bogo, M.R.; Bonan, C.D. PTZ-induced seizures inhibit adenosine deamination in adult zebrafish brain membranes. Brain Res. Bull. 2011, 86, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, F.F.; Gaspary, K.V.; Siebel, A.M.; Bonan, C.D. Oxytocin reversed MK-801-induced social interaction and aggression deficits in zebrafish. Behav. Brain Res. 2016, 311, 368–374. [Google Scholar] [CrossRef]
- Buske, C.; Gerlai, R. Shoaling develops with age in Zebrafish (Danio rerio). Prog. Neuro-Psychop. 2011, 35, 1409–1415. [Google Scholar] [CrossRef]
- Cognato, G.d.P.; Bortolotto, J.W.; Blazina, A.R.; Christoff, R.R.; Lara, D.R.; Vianna, M.R.; Bonan, C.D. Y-Maze memory task in zebrafish (Danio rerio): The role of glutamatergic and cholinergic systems on the acquisition and consolidation periods. Neurobiol. Learn. Mem. 2012, 98, 321–328. [Google Scholar] [CrossRef]
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueffl, A.V. Zebrafish models for translational neuroscience research: From tank to bedside. Trends Neurosci. 2014, 37, 264–278. [Google Scholar] [CrossRef]
- Pereira, V.M.; Bortolotto, J.W.; Kist, L.W.; de Azevedo, M.B.; Fritsch, R.S.; Oliveira, R.D.; Pereira, T.C.B.; Bonan, C.D.; Vianna, M.R.; Bogo, M.R. Endosulfan exposure inhibits brain AChE activity and impairs swimming performance in adult zebrafish (Danio rerio). Neurotoxicology 2012, 33, 469–475. [Google Scholar] [CrossRef]
- Senger, M.R.; Rico, E.P.; Arizi, M.D.B.; Frazzon, A.P.G.; Dias, R.D.; Bogo, M.R.; Bonan, C.D. Exposure to Hg2+ and Pb2+ changes NTPDase and ecto-5 ‘-nucleotidase activities in central nervous system of zebrafish (Danio rerio). Toxicology 2006, 226, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Nathan, F.M.; Parhar, I.S. Habenular kisspeptin modulates fear in the zebrafish. Proc. Natl. Acad. Sci. USA 2014, 111, 3841–3846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, M.O.; Annan, L.V.; Kanellopoulos, A.H.; Brock, A.J.; Combe, F.J.; Baiamonte, M.; Teh, M.T.; Brennan, C.H. The utility of zebrafish to study the mechanisms by which ethanol affects social behavior and anxiety during early brain development. Prog. Neuro-Psychop. 2014, 55, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonan, C.D.; Norton, W.H.J. The utility of zebrafish as a model for behavioural genetics. Curr. Opin. Behav. Sci. 2015, 2, 34–38. [Google Scholar] [CrossRef]
- Bowton, E.; Saunders, C.; Erreger, K.; Sakrikar, D.; Matthies, H.J.; Sen, N.; Jessen, T.; Colbran, R.J.; Caron, M.G.; Javitch, J.A. Dysregulation of dopamine transporters via dopamine D2 autoreceptors triggers anomalous dopamine efflux associated with attention-deficit hyperactivity disorder. J. Neurosci. 2010, 30, 6048–6057. [Google Scholar] [CrossRef]
- Liu, Q.S.; Hallerman, E.; Peng, Y.F.; Li, Y.H. Development of Bt rice and Bt maize in China and their efficacy in target pest control. Int. J. Mol. Sci. 2016, 17, 1561. [Google Scholar] [CrossRef]
- Sundaresha, S.; Rohini, S.; Appanna, V.K.; Arthikala, M.K.; Shanmugam, N.B.; Shashibhushan, N.B.; Kishore, C.M.H.; Pannerselvam, R.; Kirti, P.B.; Udayakumar, M. Co-overexpression of Brassica juncea NPR1 (BjNPR1) and Trigonella foenum-graecum defensin (Tfgd) in transgenic peanut provides comprehensive but varied protection against Aspergillus flavus and Cercospora arachidicola. Plant Cell Rep. 2016, 35, 1189–1203. [Google Scholar] [CrossRef]
- Aghazadeh, R.; Zamani, M.; Motallebi, M.; Moradyar, M.; Jahromi, Z.M. Co-transformation of canola by chimeric chitinase and tlp genes towards improving resistance to Sclerotinia sclerotiorum. World J. Microb. Biot. 2016, 32. [Google Scholar] [CrossRef]
- Zhang, F.L.; Ruan, X.L.; Wang, X.; Liu, Z.H.; Hu, L.Z.; Li, C.W. Overexpression of a chitinase gene from Trichoderma asperellum increases disease resistance in transgenic soybean. Appl. Biochem. Biotech. 2016, 180, 1542–1558. [Google Scholar] [CrossRef] [PubMed]
- Christova, P.K.; Christov, N.K.; Mladenov, P.V.; Imai, R. The wheat multidomain cystatin TaMDC1 displays antifungal, antibacterial, and insecticidal activities in planta. Plant Cell Rep. 2018, 37, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]
- Marti-Renom, M.A.; Stuart, A.C.; Fiser, A.; Sanchez, R.; Melo, F.; Sali, A. Comparative protein structure modeling of genes and genomes. Annu. Rev. Bioph. Biom. 2000, 29, 291–325. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallograph. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Luthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of protein models with three-dimensional profiles. Nature 1992, 356, 83–85. [Google Scholar] [CrossRef]
- Hess, B.; Kutzner, C.; van der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef]
- Oostenbrink, C.; Villa, A.; Mark, A.E.; Van Gunsteren, W.F. A biomolecular force field based on the free enthalpy of hydration and solvation: The GROMOS force-field parameter sets 53A5 and 53A6. J. Comput. Chem. 2004, 25, 1656–1676. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Grigera, J.R.; Straatsma, T.P. The missing term in effective pair potentials. J. Phys. Chem. 1987, 91, 6269–6271. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An NLog(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Vangunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.Z.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef]
- Defferrari, M.S.; da Silva, R.; Orchard, I.; Carlini, C.R. Jack bean (Canavalia ensiformis) urease induces eicosanoid-modulated hemocyte aggregation in the Chagas’ disease vector Rhodnius prolixus. Toxicon 2014, 82, 18–25. [Google Scholar] [CrossRef]
- Nery, L.R.; Eltz, N.S.; Hackman, C.; Fonseca, R.; Altenhofen, S.; Guerra, H.N.; Freitas, V.M.; Bonan, C.D.; Vianna, M.R.M.R. Brain intraventricular injection of Amyloid-beta in zebrafish embryo impairs cognition and increases Tau phosphorylation, effects reversed by lithium. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kappaun, K.; Martinelli, A.H.S.; Broll, V.; Zambelli, B.; Lopes, F.C.; Ligabue-Braun, R.; Fruttero, L.L.; Moyetta, N.R.; Bonan, C.D.; Carlini, C.R.; et al. Soyuretox, an Intrinsically Disordered Polypeptide Derived from Soybean (Glycine Max) Ubiquitous Urease with Potential Use as a Biopesticide. Int. J. Mol. Sci. 2019, 20, 5401. https://doi.org/10.3390/ijms20215401
Kappaun K, Martinelli AHS, Broll V, Zambelli B, Lopes FC, Ligabue-Braun R, Fruttero LL, Moyetta NR, Bonan CD, Carlini CR, et al. Soyuretox, an Intrinsically Disordered Polypeptide Derived from Soybean (Glycine Max) Ubiquitous Urease with Potential Use as a Biopesticide. International Journal of Molecular Sciences. 2019; 20(21):5401. https://doi.org/10.3390/ijms20215401
Chicago/Turabian StyleKappaun, Karine, Anne H. S. Martinelli, Valquiria Broll, Barbara Zambelli, Fernanda C. Lopes, Rodrigo Ligabue-Braun, Leonardo L. Fruttero, Natalia R. Moyetta, Carla D. Bonan, Celia R. Carlini, and et al. 2019. "Soyuretox, an Intrinsically Disordered Polypeptide Derived from Soybean (Glycine Max) Ubiquitous Urease with Potential Use as a Biopesticide" International Journal of Molecular Sciences 20, no. 21: 5401. https://doi.org/10.3390/ijms20215401