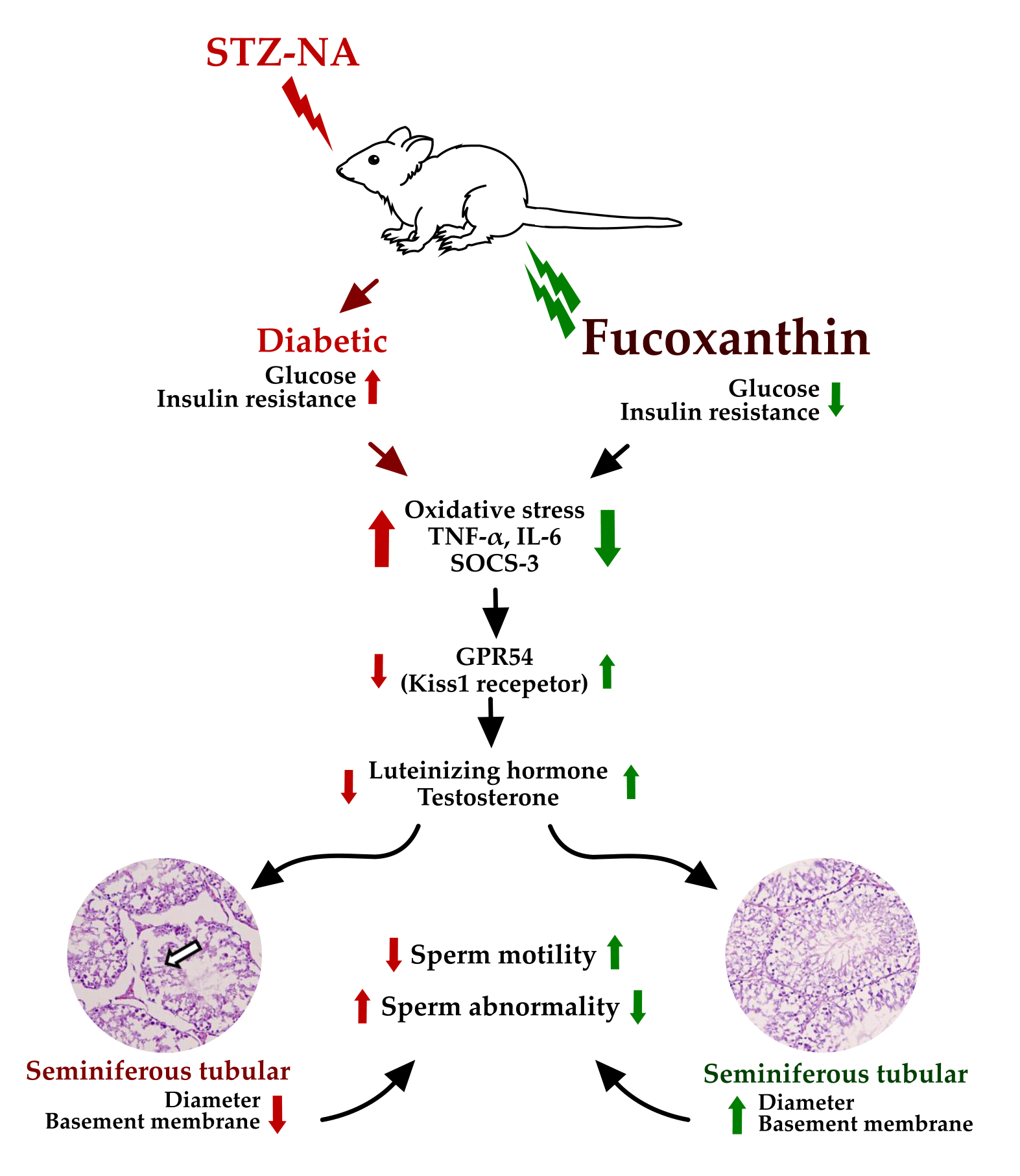
1. Introduction
Diabetes mellitus (DM) has been recognized as a metabolic disorder which occurs due to failure of insulin secretion, insulin action, or both. Type-1 diabetes is characterized by autoimmune-mediated pancreatic β-cell and results in the deficiency of insulin, whereas type-2 diabetes is peripheral insulin resistance [
1]. These conditions were characterized by an increase in blood glucose or hyperglycemia [
2]. Hyperglycemia increases the level of proinflammatory cytokines, such as interleukin (IL)-1β, IL-2, and IL-6 in the serum of DM patients and has been reported by several studies [
3,
4]. Changes in antioxidant capacity and oxidative stress also have a positive correlation with cell dysfunction and contribute to DM disease [
5]. DM disease impairs some organs, such as pancreas, liver, kidney, and testis [
6,
7]. In human and experimental models, DM also shows the adverse effects on reproductive function including essential hormone impairment of reproduction (i.e., testosterone, luteinizing hormone, follicle-stimulating hormone) or hypogonadism, sperm motility, and sperm deformity in spermatogenesis [
8,
9].
Modulation of oxidative stress represents an important strategy for the treatment of DM disease [
6]. Insulin secretagogues, alpha-glucosidase inhibitors, and thiazolidinedione (i.e., rosiglitazone) have been used for DM treatment. However, some antidiabetic agents possess adverse effects, such as headache, increased prevalence of gastrointestinal, and cardiovascular diseases [
10,
11]. Therefore, the investigation of novel antidiabetic agents with less adverse effects is a major challenge in the future research through natural products or functional foods.
Fucoxanthin is a natural product that can be extracted from brown seaweeds (i.e.,
Undaria pinnatifida, Saccharina japonica, Sargassum fulvellum) and is rich in xanthophyll content (one form of carotenoids) [
12,
13]. The ability of fucoxanthin to ameliorate several diseases, such as hypertension, obesity, anti-inflammatory effects, and anti-carcinogenic activity has been reported [
14]. Previous studies reported that fucoxanthin attenuated oxidative stress by scavenging reactive oxygen species and increasing superoxide dismutase activity [
15,
16]. Fucoxanthin-rich extracts from
Sargassum muticum reduced inflammation and attenuated colitis disease in mice models [
17]. A study reported that fucoxanthin extract shows anti-obesity and anti-diabetic effects by improving insulin resistance and reducing adipokine in white adipose tissue, as well as upregulating glucose transporter 4 skeletal muscle [
13]. According to these conditions, we hypothesized that fucoxanthin extract from brown algae (
Laminaria japonica) possesses anti-diabetes effects and also can improve reproductive function in an experimental diabetic model. Therefore, this study aimed to investigate the ameliorative effects of fucoxanthin-rich extract from
Laminaria japonica on male reproductive function in a diabetic rat model.
3. Discussion
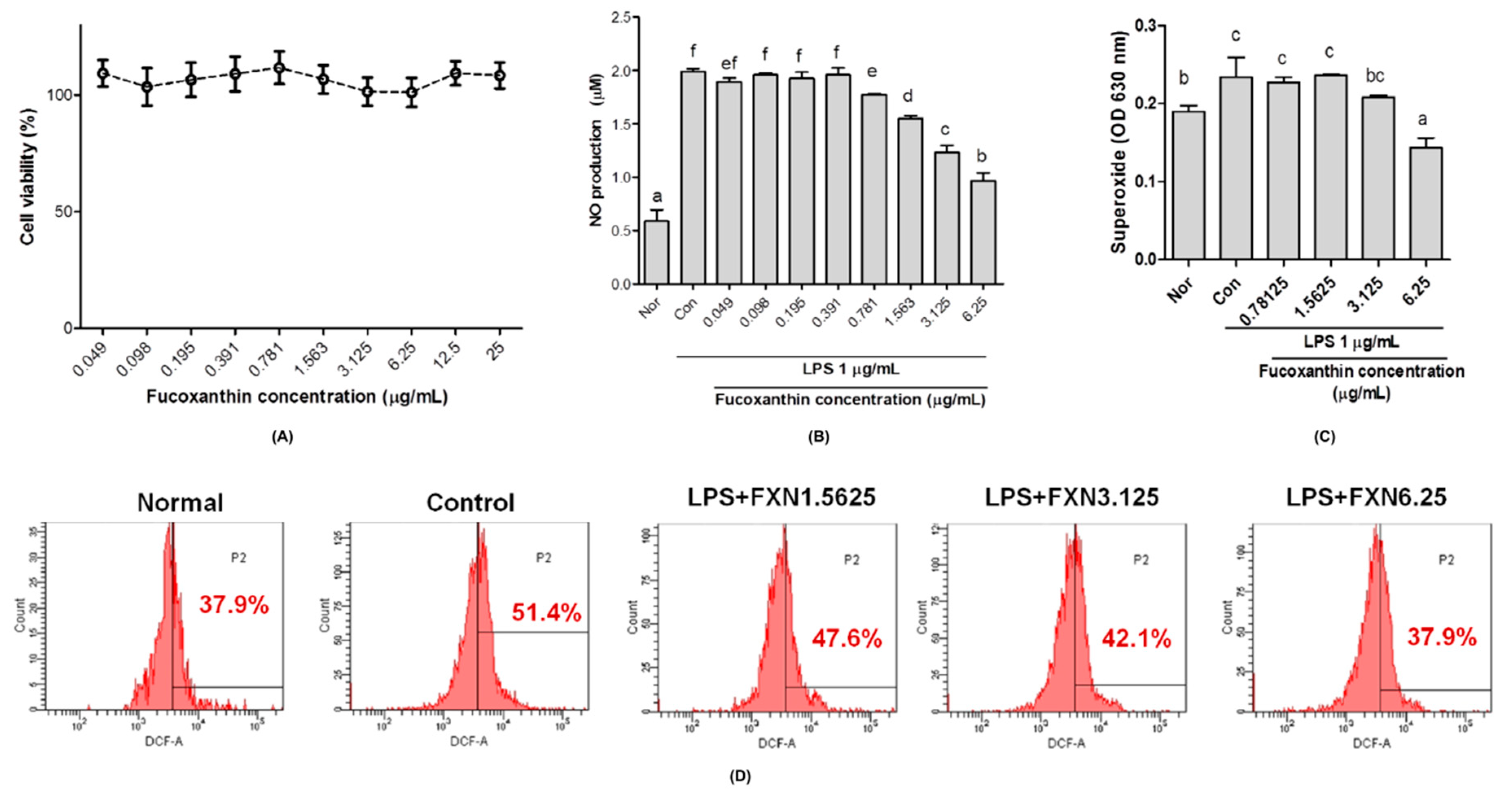
In vitro studies showed that the fucoxanthin extract from
Laminaria japonica has no effect on the proliferation of RAW 264.7 macrophages cells, which was indicated from high cell viability although in the high concentration of fucoxanthin (
Figure 1A). Fucoxanthin extract attenuated oxidative stress by reducing nitric oxide (NO), superoxide anion (O
2−), and hydrogen peroxide (H
2O
2) production in RAW 264.7 cells stimulated by lipopolysaccharides (
Figure 1C,D, respectively). NO can induce oxidative stress and apoptosis in cell studies as it is recognized as a proinflammatory mediator and marker for oxidative stress status [
18,
19,
20]. A previous study reported that fucoxanthin possesses a protective effect on H
2O
2-induced oxidative damage in human hepatic cells by inhibiting translocation of factor-erythroid 2-related factor 2 [
21]. Additionally, fucoxanthin also showed an anti-apoptotic effect on carbon tetrachloride-induced hepatotoxicity by enhancing heme oxygenase-1 expression [
22].
In vivo studies of fucoxanthin against reproductive functions was evaluated by using diabetes male rats. The diabetes condition was successfully induced by streptozotocin-nicotinamide (STZ-NA). A previous study reported that one of the non-insulin-dependent diabetes mellitus (NIDDM) experimental models without obesity was partially pancreatectomized rats and rats subjected to neonatal administration of STZ. This condition was characterized by β cell mass reduction that was considered also occurs in the NIDDM [
23]. Whereas, type 2 diabetes, formerly known as NIDDM was a metabolic disorder, which occurs due to the failure of insulin action [
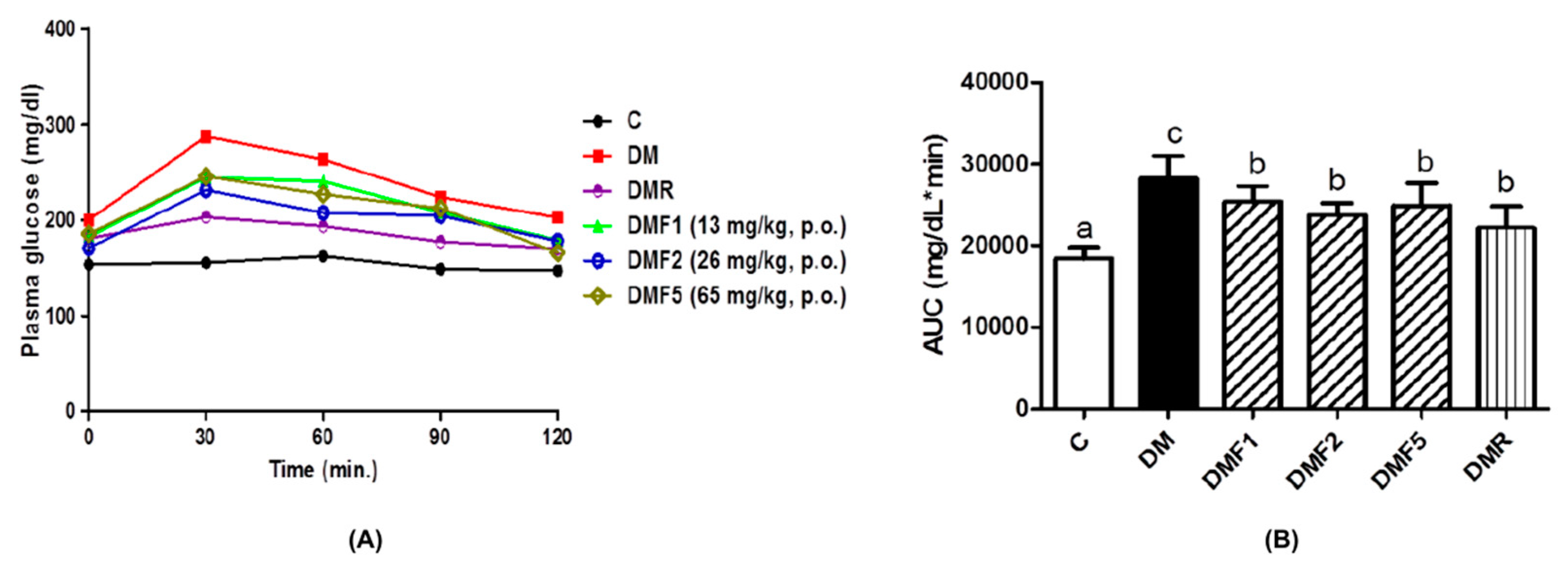
24]. The generation of diabetic conditions was confirmed by increasing plasma glucose concentration and high homeostasis model assessment as an index of insulin resistance (HOMA-IR) (
Figure 2 and
Table 2). These results were supported by a previous study in which STZ-NA increases plasma glucose concentration in rats after four weeks of administration. STZ is well recognized at inducing the damage of pancreatic B-cells, whereas NA partially protected the cells against STZ [
23]. The measurement of plasma glucose levels is one biomarker for diabetes diagnosis and HOMA-IR is widely used for insulin resistance (IR) estimation [
25]. In this study, the plasma glucose concentration and IR index were successfully reduced after four weeks of treatment with fucoxanthin extract.
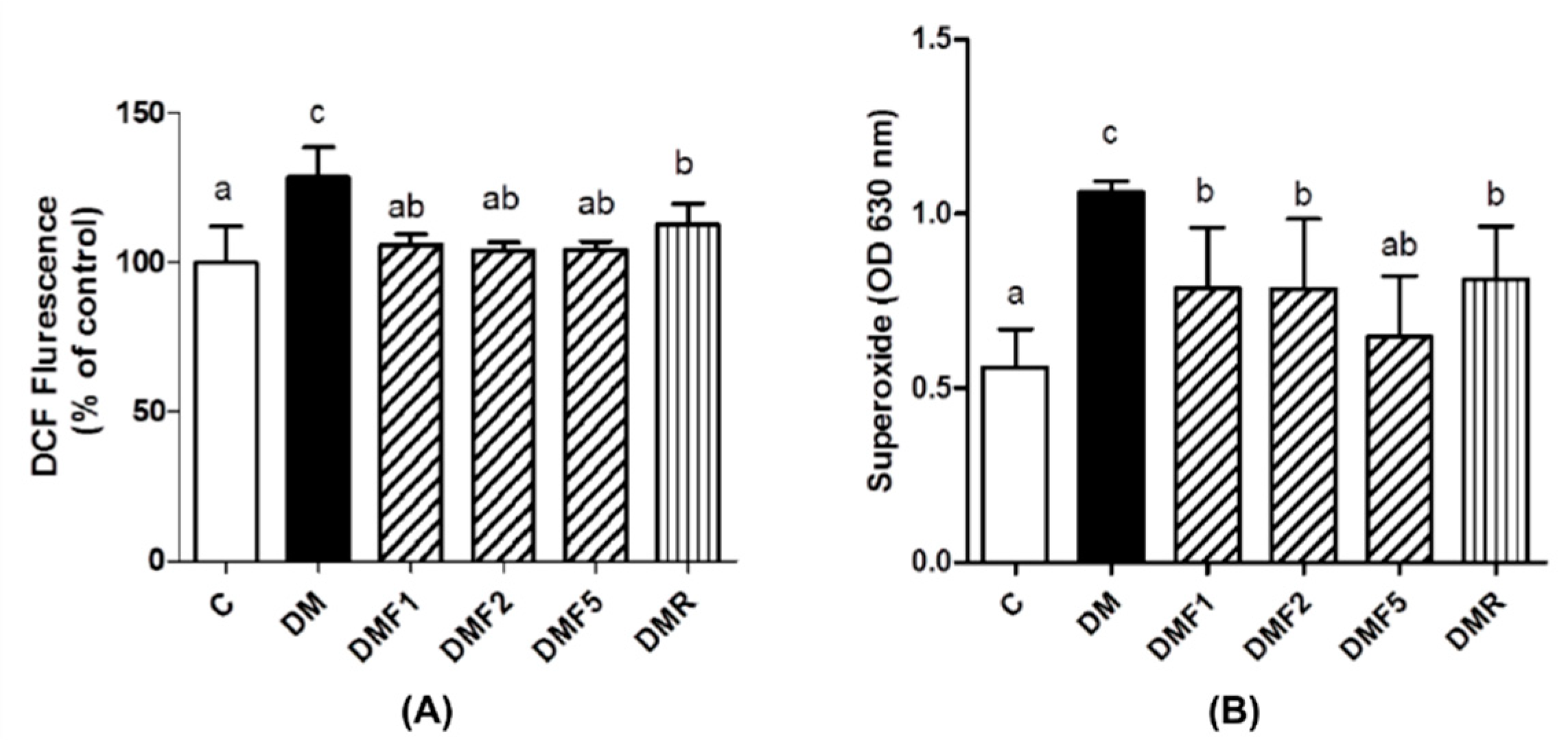
Low levels of enzymatic antioxidant (
Table 2 and
Table 3) and high levels of oxidative stress condition were observed in the untreated diabetes (DM) group (
Figure 3). However, fucoxanthin extracts successfully enhanced enzymatic antioxidant levels9 (catalase; superoxide dismutase, SOD; glutathione peroxidase, GPx) and also ameliorated oxidative stress by reducing reactive oxygen species (ROS) and superoxide anion (O
2−) productions in rat sperm (
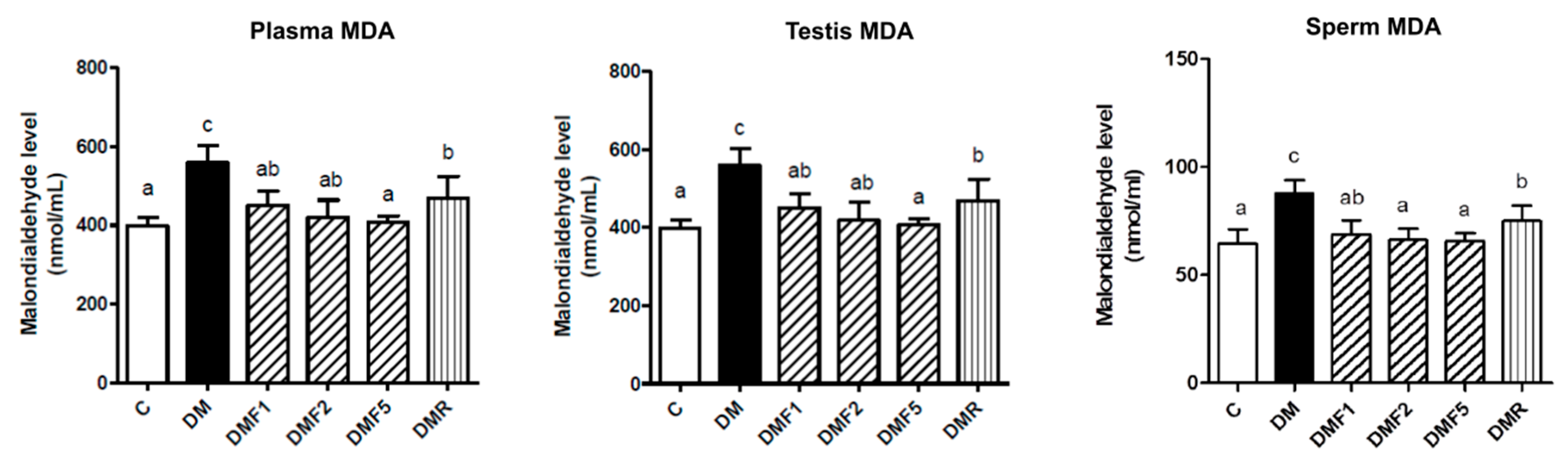
Figure 3). Additionally, fucoxanthin also reduces the malondialdehyde (MDA) level in plasma, testis, and sperm of rats, as shown in
Figure 4. MDA has been used for a biomarker of lipid peroxidation product [
26]. SOD antioxidant prevents the tissue from producing highly reactive of O
2− content by converting them to hydrogen peroxide (H
2O
2), whereas catalase acts as catalytic decomposition of harmful H
2O
2 to oxygen (O
2) and water (H
2O). In the absence of catalase or GPx, H
2O
2 leads to the production of reactive hydroxyl radicals and makes free radicals to easier for attack the tissues [
27]. Based on these results, fucoxanthin has the potential to ameliorate ROS production and acts as a scavenger for free radicals. This condition can also occur due to the inhibition of hyperglycemia by fucoxanthin. Additionally, previous studies also reported that fucoxanthin reduced ROS production by inhibiting nicotinamide adenine dinucleotide phosphate (NADPH) oxidase-4 (NOX-4) activity [
28]. Whereas, NOX-4 are known as ROS-generating enzymes which produce ROS [
29]. Hyperglycemia has been known to induce oxidative stress and be involved in the pathogenesis of diabetes [
3]. Oxidative stress has been positively correlated with male infertility due to sperm dysfunction [
30]. Additionally, a high level of ROS also contributed to abnormal spermatozoa formation [
31].
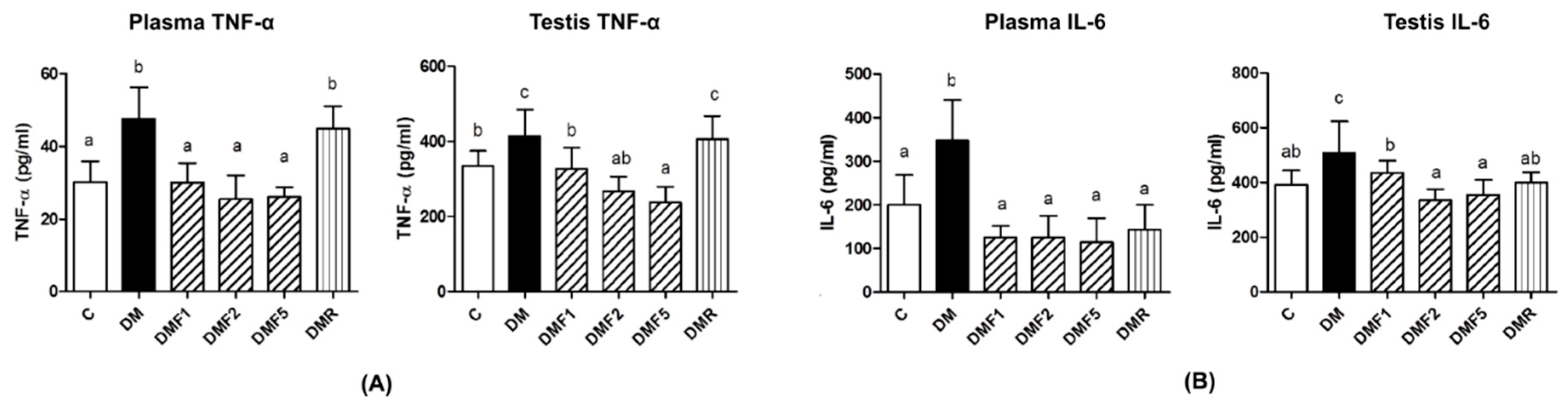
Besides increasing the oxidative stress level, hyperglycemia also increases proinflammatory cytokine levels, such as interleukin-6 and tumor necrosis factor (TNF)-α [
4,
32]. These proinflammatory cytokines also play a role in systemic inflammation and they are related to insulin resistance in diabetes condition [
33]. These proinflammatory cytokines have also caused reproductive dysfunction by triggering testicular apoptosis and atrophy [
34]. In
Figure 5, untreated-diabetes (DM) groups showed higher levels of TNF-α and IL-6 in plasma and testis than control and treated groups. After four weeks of treatment by fucoxanthin, it reduced proinflammatory cytokine levels.
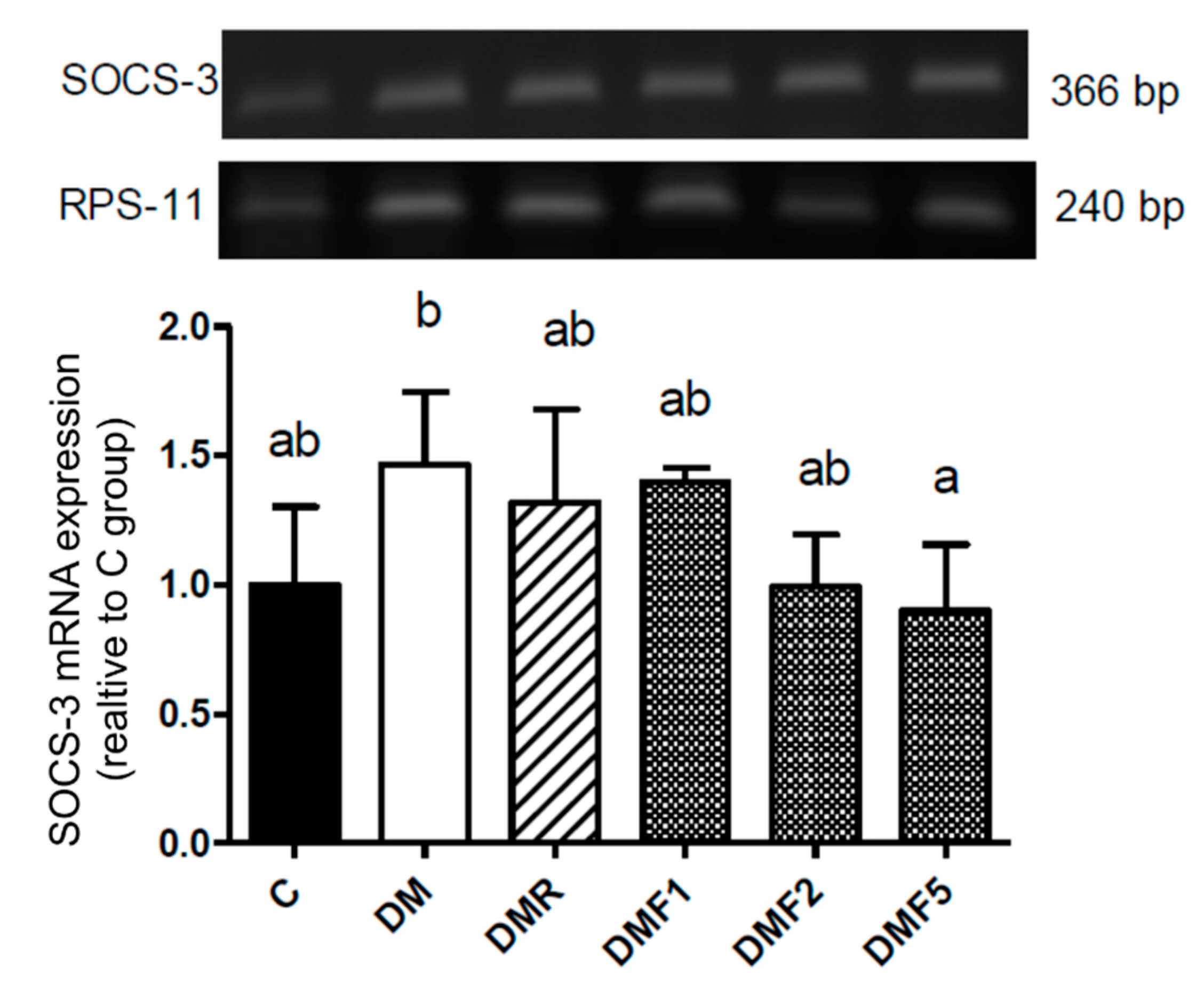
Additionally, the diabetic condition (DM group) showed a high expression of suppressor of cytokine signaling-3 (SOCS-3) (
Figure 6). SOCS family has been reported that inhibit transduction of insulin receptor signaling and resulting in insulin resistance. High level of IL-6 and blood glucose also may induce expression of SOCS-3 in the diabetic patient [
35]. We hypothesized that the high expression of SOCS-3 also contributes to severe reproductive dysfunction. Treatment with fucoxanthin successfully inhibited its expression.
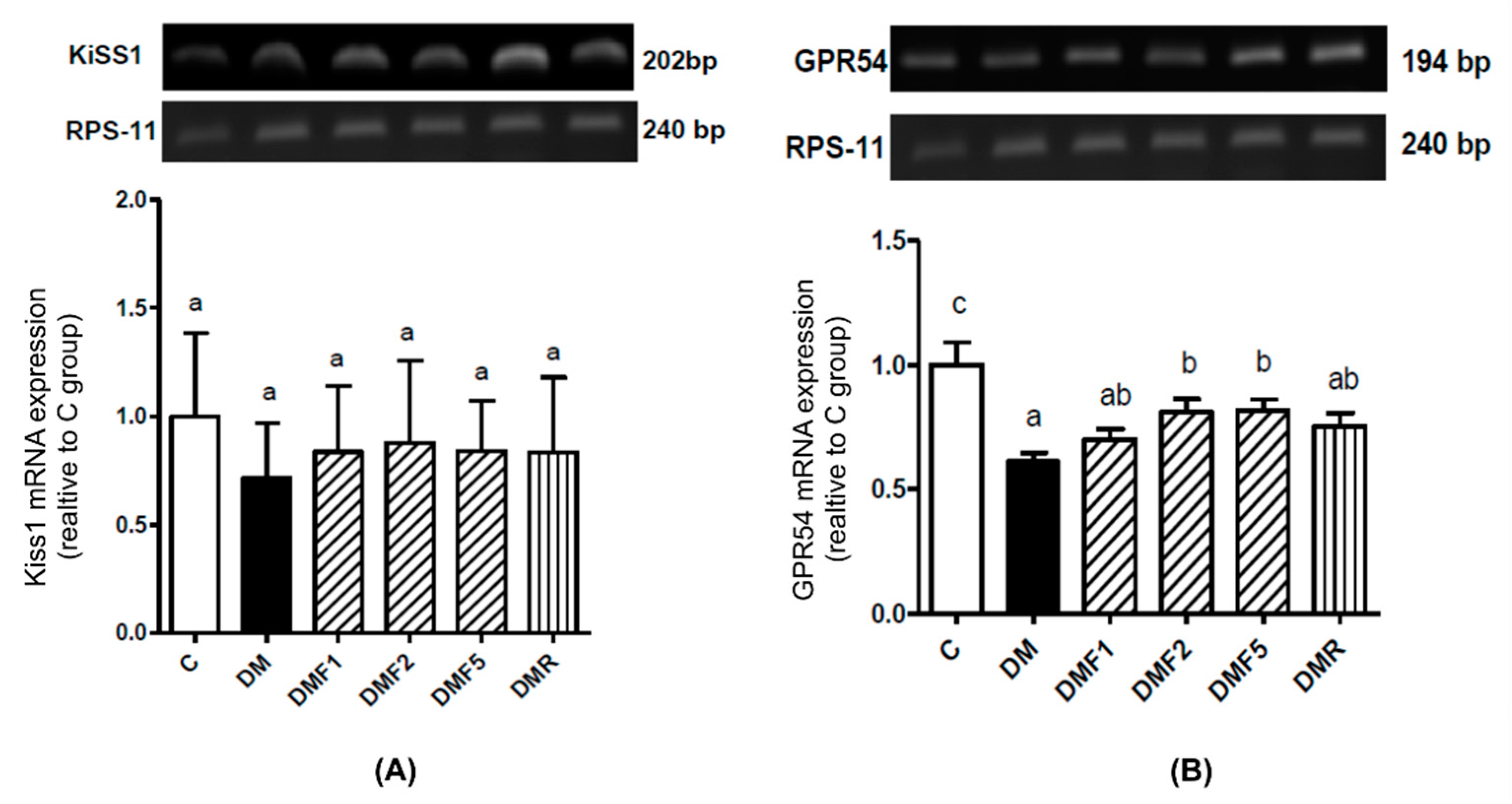
In this study, low expression of G-protein coupled receptor 54 (GPR54, Kiss1 receptor) was shown in the diabetic (DM) group. Fucoxanthin treatment significantly increases the GPR54 expression in rat hypothalamic, as shown in
Figure 7. Complex Kiss1/Kiss1 receptors are used in reproductive function regulation by controlling gonadotropin secretion and are involved in testicular function [
36]. Low levels of Kiss1 or Kiss1 receptors might be caused by high levels of proinflammatory cytokines. A previous study reported that TNF-α reduced Kiss1 receptor expression [
37]. Additionally, insulin expression also impaired reproductive function by modulating kisspeptin signaling [
38].
Hypogonadism associated with diabetes is characterized by a low level of luteinizing hormone (LH) and testosterone observed in the diabetic condition (
Table 4). Diabetes has been reported to disrupt LH from circulating, resulting in increased testis resistance against this hormone, leading to a low level of testosterone [
39]. Treatment of diabetic rats with fucoxanthin has an ameliorated effect on the level of LH and testosterone. Based on this condition, we hypothesized that fucoxanthin administration successfully protects the gonadotropin hormone LH against oxidative stress and improve the action of Kiss1/Kiss1 receptor in the hypothalamic-pituitary-gonadal (HPG) axis. Kiss1 has been reported as a gonadotropin hormone stimulator by stimulating gonadotropin-releasing hormone (GnRH) in the hypothalamus [
40].
Overall, based on the biochemical observation, fucoxanthin administration successfully ameliorated diabetic condition as well as improved the gonadotropin hormone and might improve male reproductive dysfunction. To confirm the improvement effects of fucoxanthin, we have observed the sperm count, motility and morphology. As shown in
Table 5, the diabetic condition leads to reducing sperm motility and increasing abnormal morphology. Diabetes has been known to give rise to an abnormality of sperm morphology, whereas, structural and functional changes throughout sperm tail morphogenesis led to a decrease in sperm motility [
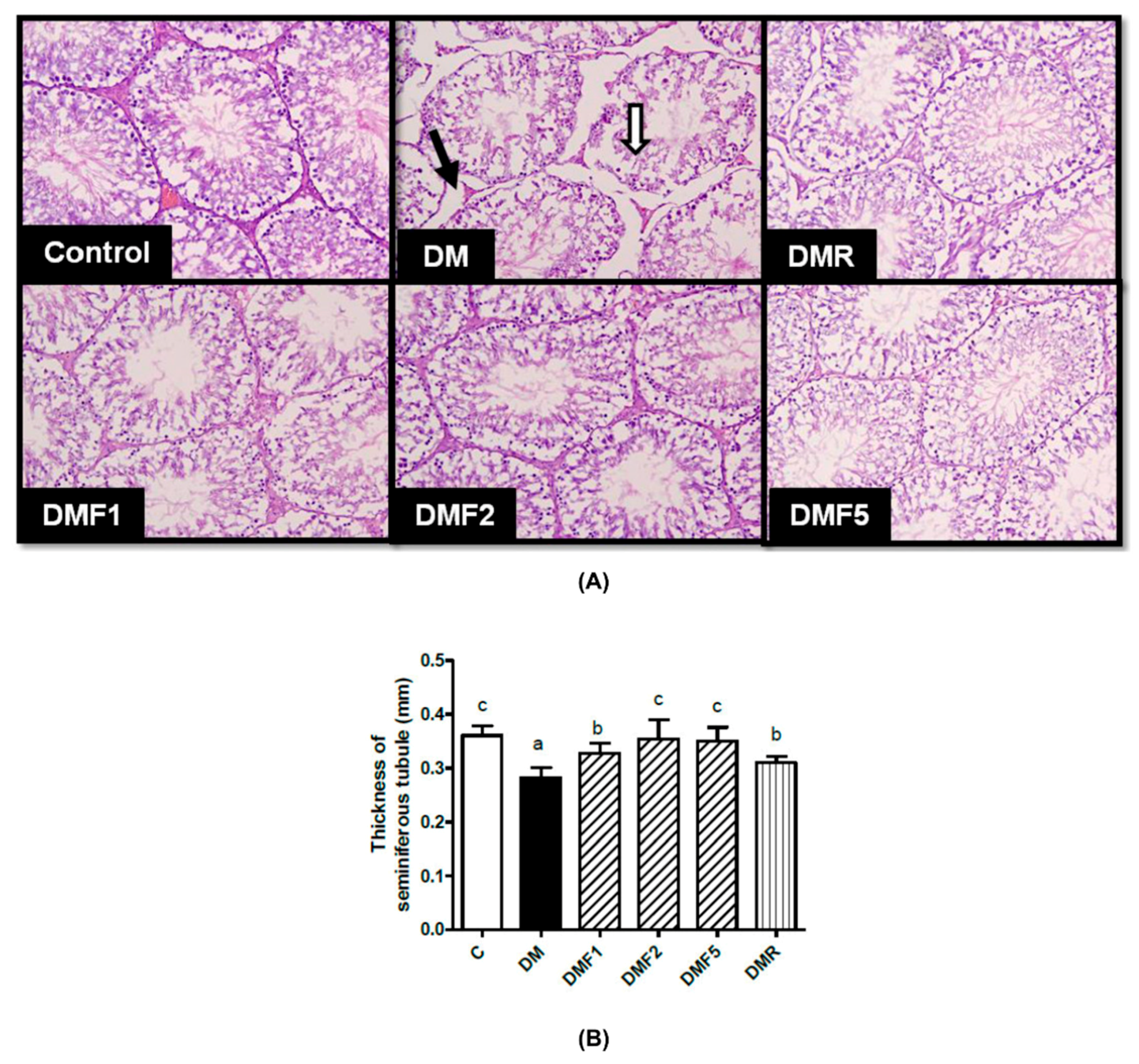
41]. Treatment with fucoxanthin increases sperm motility. In the case of the seminiferous tubular structure,
Figure 8 showed seminiferous tubular degeneration, shrunken, and separation from each other as well as high thickness basement membrane of seminiferous tubulars in the diabetic group. After treatment for four weeks, fucoxanthin successfully ameliorated seminiferous tubular morphology. This condition was supported by a previous study that the basement membrane of the seminiferous tubular and its diameter decreased in diabetic rats [
42]. Additionally, continual renewal of spermatogonia and their differentiation into spermatogenic cells are the regulators for male fertility [
43]. A previous study reported that in type-1 diabetes patients, they had lower progressive motility of spermatozoa compared to the normal condition [
44].
Used in this study as a positive control; rosiglitazone has been demonstrated to enhance insulin sensitivity [
45]. Additionally, rosiglitazone also ameliorates oxidative stress and inflammation by reducing MDA level, enhancing SOD activity and total antioxidant capacity, as well as decreasing TNF-α and IL-6 level in the diabetic patient [
46]. On the other hand, rosiglitazone has been reported to associate with cardiovascular diseases [
47,
48]. Although rosiglitazone was reported as having a low risk of gastrointestinal side effects [
49].
4. Materials and Methods
4.1. Materials
The fucoxanthin was extracted from brown algae Laminaria japonica. Fucoxanthin extract was provided by Oryza Oil & Fat Chemical Co., Ltd. (Tokyo, Japan). The RAW 264.7 macrophage cells were supplied from American Type Culture Collection (ATCC). Dulbecco’s Modified Eagle Medium (DMEM) was purchased from GIBCO (Carlsbad, CA, USA). The fetal calf serum (FCS) was purchased from HyClone (Carlsbad, CA, USA). The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) powder, nitro blue tetrazolium (NBT) powder, dimethyl sulfoxide (DMSO), 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) buffer, dichloro-dihydro-fluorescein diacetate (DCFH-DA), and Trypan Blue were purchased from Sigma Aldrich (St. Louis, MO, USA). The glucose enzymatic kit was purchased from Kyokuto Pharmaceutical Industrial Co., Ltd. (Tokyo, Japan) and insulin ELISA kits was purchased from Mercodia AB Inc. (Uppsala, Sweden). Luteinizing hormone (LH) RIA kits were provided by Dr. A. F. Parlow, National Institute of Diabetes and Digestive and Kidney Diseases, National Hormone and Peptide Program (Torrance, CA, USA) and testosterone EIA as well as enzymatic antioxidant (catalase, glutathione peroxidase, superoxide dismutase) commercial kits were purchased from Eugene Chen Co., Ltd. (Taipei, Taiwan).
4.2. RAW 264.7 Macrophage Cells Study
4.2.1. Cell Culture
The RAW 264.7 macrophages cells (1 × 106 cells/disk) were cultured in 10 mL of complete culture medium (Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 100 U/mL of Penicillin-Streptomycin and 10% fetal calf serum (FCS)) in 75 cm2 disk culture. The cells were incubated at 37 °C in an atmosphere of 5% CO2. The cells were sub-cultured every 2 days until the cell count was approximately sufficient to be used. The sub-culture was harvested and prepared by scraping methods by using a cell scraper.
4.2.2. Cells Viability and Nitric Oxide Assay
Cell viability was estimated by the 3-[4,5-dimethylthiazol-2yl]-2,5-diphenyl-tetrazolium bromide (MTT) assay by following the previous method [
50]. The cells (2 × 10
5 cells/well) were plated into 96-well plates in complete medium, pre-incubated for 2 h with fucoxanthin, and added with LPS (1 µg/mL) then incubated at 37 °C for 24 h. After the incubation time, the culture supernatant was taken out and put into a new 96-well plate for the nitric oxide (NO) assay and then each well was washed by a phosphate buffer saline (PBS) and added to 100 µL of MTT (5 mg/mL) solution for 4 h in a CO
2 incubator at 37 °C. Then, the cells were dissolved with 100 μL of dimethyl sulfoxide (DMSO) and shaken in the dark at room temperature for 15 min. Cell viability was estimated by the colorimetric assay of formazan intensity in a plate reader at 570 nm. The NO assay was determined using the Griess reagent as described by previous methods [
51]. Briefly, 50 µL of cell culture supernatant was mixed with 50 µL of Griess reagent (Sulfanilamide and N-(1-Naphthyl)-ethylenediamine), the mixture was incubated at room temperature for 10 min and light was avoided, and the absorbance at 540 nm was measured in a microplate reader.
4.2.3. Reactive Oxygen Species (ROS) Assay In Vitro
The reactive oxygen species (ROS) level was evaluated by using dichloro-dihydro-fluorescein diacetate (DCFH-DA) staining and nitro blue tetrazolium (NBT) reduction assay. The hydrogen peroxide (H
2O
2) level was measured by using the probe 2′,7′-dichlorofluorescein-diacetate (DCFH-DA) staining method as described by previous methods [
52]. Briefly, RAW 264.7 cells (2 × 10
5 cells/well) were cultured in 12 wells with LPS (1 µg/mL) either untreated or treated with fucoxanthin and incubated at 37 °C for 24 h. Then, 1 mL of cells culture was loaded with the stain DCFH-DA and incubated for 30 min at 37 °C and then centrifuged at 400×
g for 4 min in the dark. After washing the cells twice with PBS, the conversion of DCFH to dichlorofluorescein (DCF) was observed, which had a green fluorescent (DCF-DA) color that was detected and evaluated between 500 and 530 nm by flow cytometry. Whereas, superoxide anion (O
2−) was determined by using NBT assay according to previous methods [
53]. Briefly, RAW264.7 cells (10
6 cells/well) were cultured with LPS (1 µg/mL) either untreated or treated with fucoxanthin and incubation at 37 °C for 18 h. The supernatant was removed and added with NBT solution (0.1 mg/mL NBT, 5% FCS, 3% DMSO in 10 mL of DMEM) and incubation for 1 h, then centrifuged to remove the supernatant and dissolved in DMSO. The absorbance was measured at 620 nm by using the ELISA reader.
4.3. Animal Experiment
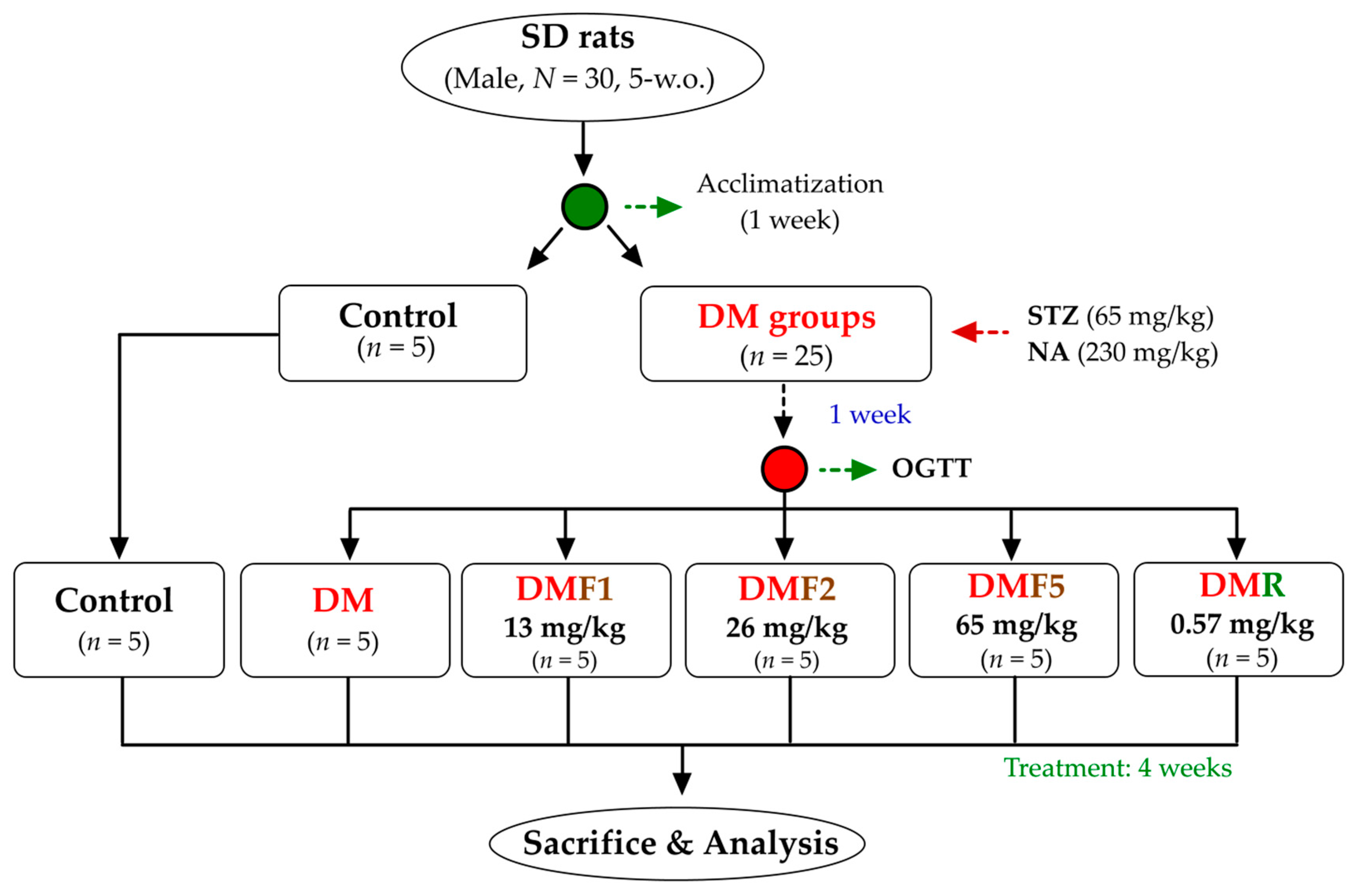
Five-weeks-old of male Sprague-Dawley rats (200 ± 10 g) were maintained under standard laboratory conditions at temperature 22–25 °C under 12 h light/dark cycle. The Institutional Animal Care and Use Committee (IACUC Approval No. 101038; Date: 30 May 2012) of the National Taiwan Ocean University reviewed and approved all protocols. Briefly, thirty SD rats were housed individually and provided standard rodent diet (LabDiet 5001) and water ad libitum. After a 1-week acclimatization phase, the rats were divided into 2 main groups (
Figure 9). The first group was a control group (
n = 5), whereas the second group was a diabetic group (
n = 25). The diabetes model was induced by intraperitoneal administration of streptozotocin (STZ, 65 mg/kg of body weight) followed by nicotinamide (NA, 230 mg/kg) 15 min later according to a previous method [
23]. After a 1-week induction of STZ-NA, the oral glucose tolerance test (OGTT) was performed to evaluate diabetes condition. The diabetes rats were divided into equally into 5 groups. The first group was diabetes rats without any treatment (DM) and another group were treated with either one of three different doses of fucoxanthin (DMF1, 13 mg/kg; DMF2, 26 mg/kg; DMF5, 65 mg/kg) or rosiglitazone (DMR, 0.57 mg/kg) for 4 weeks. The OGTT protocol was performed according to a previously used method [
54]. The OGTT was carried out by orally administering glucose (2 g/kg) after 12 h of fasting and blood was drawn to measure glucose levels at 0, 30, 60, 90, and 120 min after glucose injection. The rats were euthanized by CO
2 exposure in an empty chamber. The whole blood and organs (hypothalamus, epididymis, and testis) were collected then kept for future analysis.
4.4. Blood Collection, Supernatant Homogenized Tissue, and Sperm Cells Preparation
The whole blood was collected by using a heparinized-syringe to collection tube. Blood was then centrifuge at 1000×
g for 15 min to separate the plasma (supernatant). The blood plasma was then kept at −80 °C for future analysis [
55]. Whereas, supernatant homogenized tissue (hypothalamus and testis) was adapted from the ELISA kit manufacture’s protocol and previous methods [
56,
57]. Briefly, 100 mg of tissue was suspended in 900 μL of cold PBS and then homogenized by using a micro-tube homogenizer. The tissue preparation then kept in −20 °C overnight and then thawed in room temperature before use. Afterward, it was centrifuged at 5000 rpm for 15 min. The supernatant was collected for future analysis.
Sperm cells were prepared by using the swim-up technique from epididymis tissue according to the previous method [
58]. Briefly, the epididymides tissues were cut in the Roswell Park Memorial Institute medium (RPMI), they were shaken at 100 rpm for 10 min to remove seminal plasma, and the pellet was suspended in fresh RPMI medium. The preparation was shaken at 100×
g for 10 min, incubated for 37 °C in 5% CO
2 incubator for 30 min, and the sperm cells were collected from the supernatant for future analysis.
4.5. Reactive Oxygen Species Analysis
The catalase, glutathione peroxidase (GPx), and superoxide dismutase (SOD) enzymatic antioxidants of plasma and supernatant of homogenized testis tissue were analyzed by using commercial kits. Whereas, malonaldehyde (MDA) was estimated by following the previous method [
59]. Plasma or supernatant of homogenized tissue (100 μL) was mixed with 200 μL of a reactive solution (15% (
w/
v) trichloroacetic acid in 0.25 N HCl and 0.375% (
w/
v) thiobarbituric acid in 0.25 N HCl) and was placed at 100 °C in a water bath for 15 min. After the cooling process, the mixture was added by 300 μL of
n-butanol, and it was centrifuged at 1500×
g for 10 min. The clear supernatant was measured at the 532 nm absorbance. For the sperm MDA analysis, 1 mL of sperm (10
6 cells/mL) was added to 2 mL of reactive solution and then followed the same procedure with plasma or supernatant of tissue analysis.
4.6. Glucose, Insulin, Proinflammatory Cytokines, and Hormones Analysis
The blood glucose was determined by using commercial kits. Insulin, tumor necrosis factor (TNF)-α, and interleukin (IL)-6 were analyzed by using enzyme-linked immunosorbent assays (ELISA) kits. The luteinizing hormone (LH) was analyzed by using immunoradiometric assay (RIA) kits. Whereas, the testosterone was analyzed by using ELISA kits. All the procedures were performed according to the manufacturer’s protocols.
4.7. Kiss1, GPR54, and SOCS-3 mRNA Expression Analysis
The quantitative of Kiss1, GPR54, and SOCS-3 mRNA expression was determined by using the conventional reverse transcription polymerase chain reaction (PCR) technique. The RNA was prepared by using RNeasy Lipid Tissue Mini Kit (Qiagen, Hilden, Germany) and cDNA was prepared using HiScript I Reverse Transcriptase (Bionovas Biotechnology Co., Ltd., Ontario, CA, USA) according to the manufacturer’s protocols. PCRs were performed in a 30 μL reaction volume containing 3 μL of cDNA, 10 × Taq buffer (containing 15 mM MgCl2), Taq DNA polymerase (5 U/μL), and dNTP mixture (10 mM) in order. The PCR primers specific to each gene was described in
Table 6. Then, 2% agarose gel was prepared and each well was loaded to the DNA of the sample and the DNA marker, which was mixed with tracking dye. After electrophoresis, the gel was stained with ethidium bromide (EtBr). After dyeing, the gel was observed by UV light and photographed. The image was quantified and compared with the marker to determine the molecular weight. The data of gene quantification obtained by computer analysis was divided by internal control, then used as a result.
4.8. Sperm Count and Morphology Assay
The procedure of sperm count and morphology assay followed the previous method [
60,
61]. This method studied the sperm shape abnormality in cauda epididymis of the rats. A thousand sperms per animal were screened to the different types of abnormality in one of the cauda epididymis. Five types of abnormalities, such as amorphous, hookless, banana shape, fused and double-headed were evaluated and finally represented as percentage total abnormality.
4.9. Testis Histopathological Analysis
The testis of rats was dissected, fixed in 10% neutral buffered formalin, and processed for paraffin embedding, and stained with hematoxylin and eosin (H&E) according to previous methods [
62]. The seminiferous tubules structure was observed under an inverted vertical phase-contrast microscope (Olympus CK2, Japan).
4.10. Statistical Analysis
All the values were shown as the mean ± standard deviation (S.D.). To determine the significant (p < 0.05) differences among all groups using all the parameters, one-way analyses of variance (ANOVA) with Duncan post hoc test were used using SPSS 22.0 (IBM Corp., Armonk, NY, USA).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}