Effects of Foliar Redox Status on Leaf Vascular Organization Suggest Avenues for Cooptimization of Photosynthesis and Heat Tolerance

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
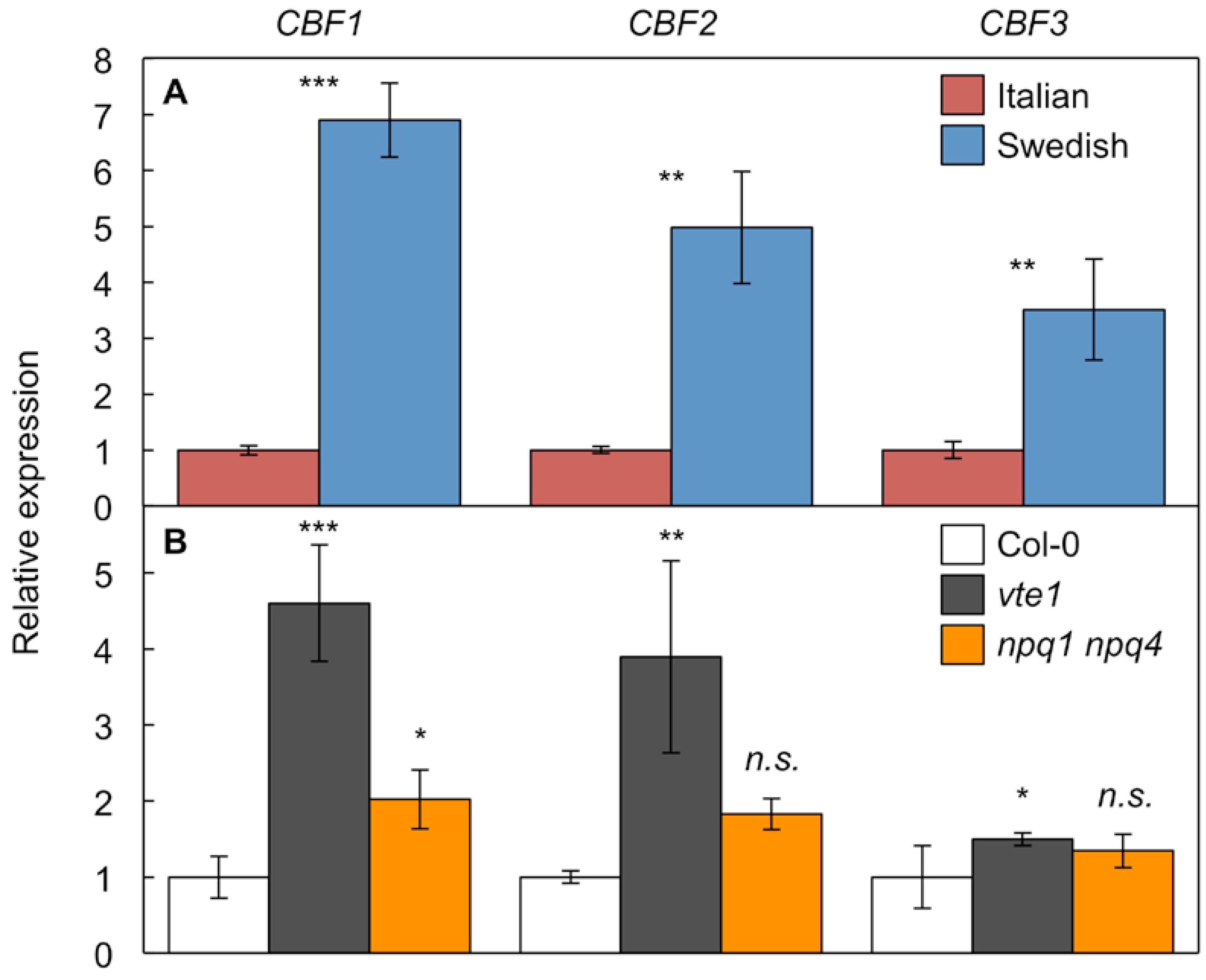
2. Results
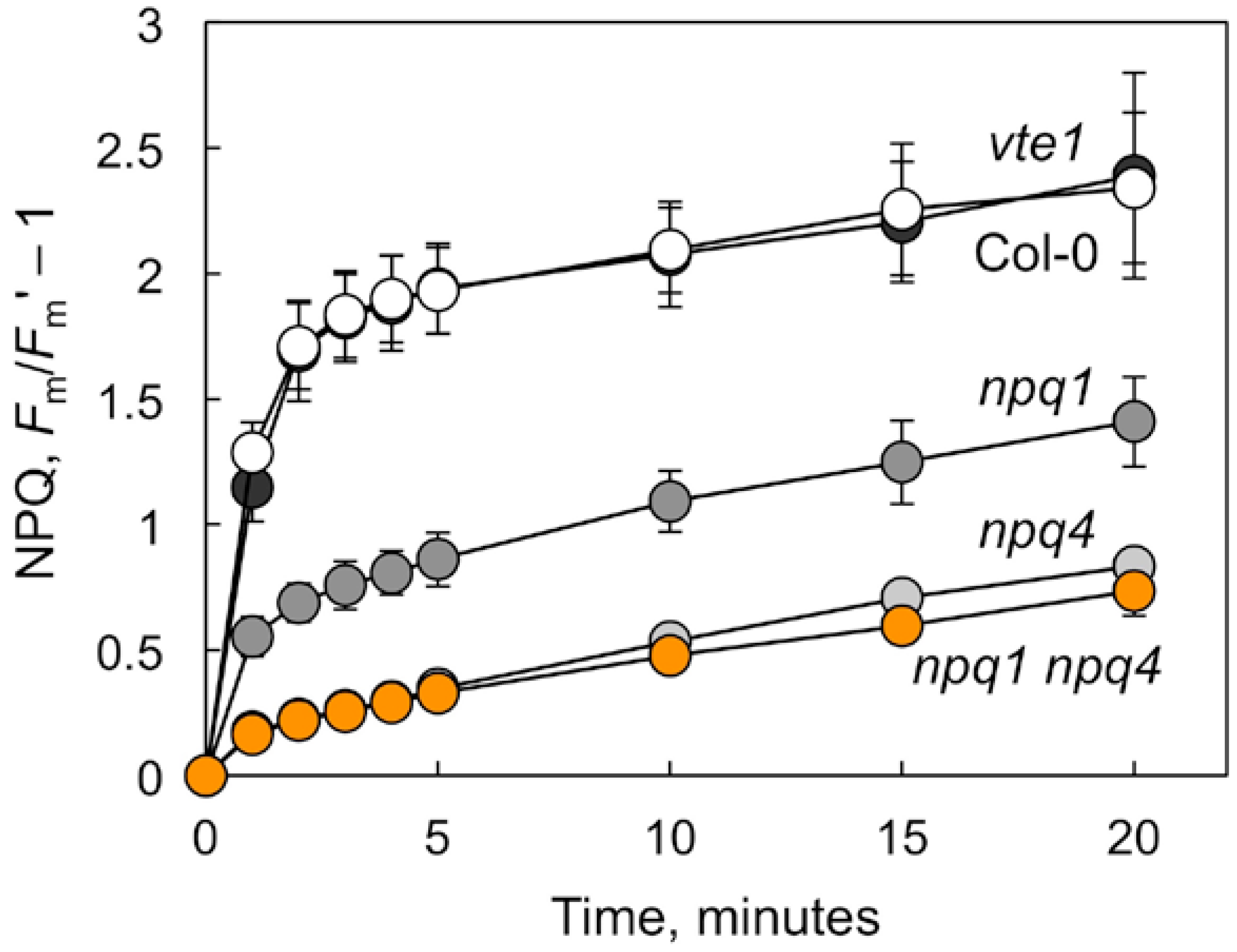
2.1. Non-photochemical Quenching and Foliar Pigment Levels in Mutants Deficient in Thermal Energy Dissipation or in Tocopherols
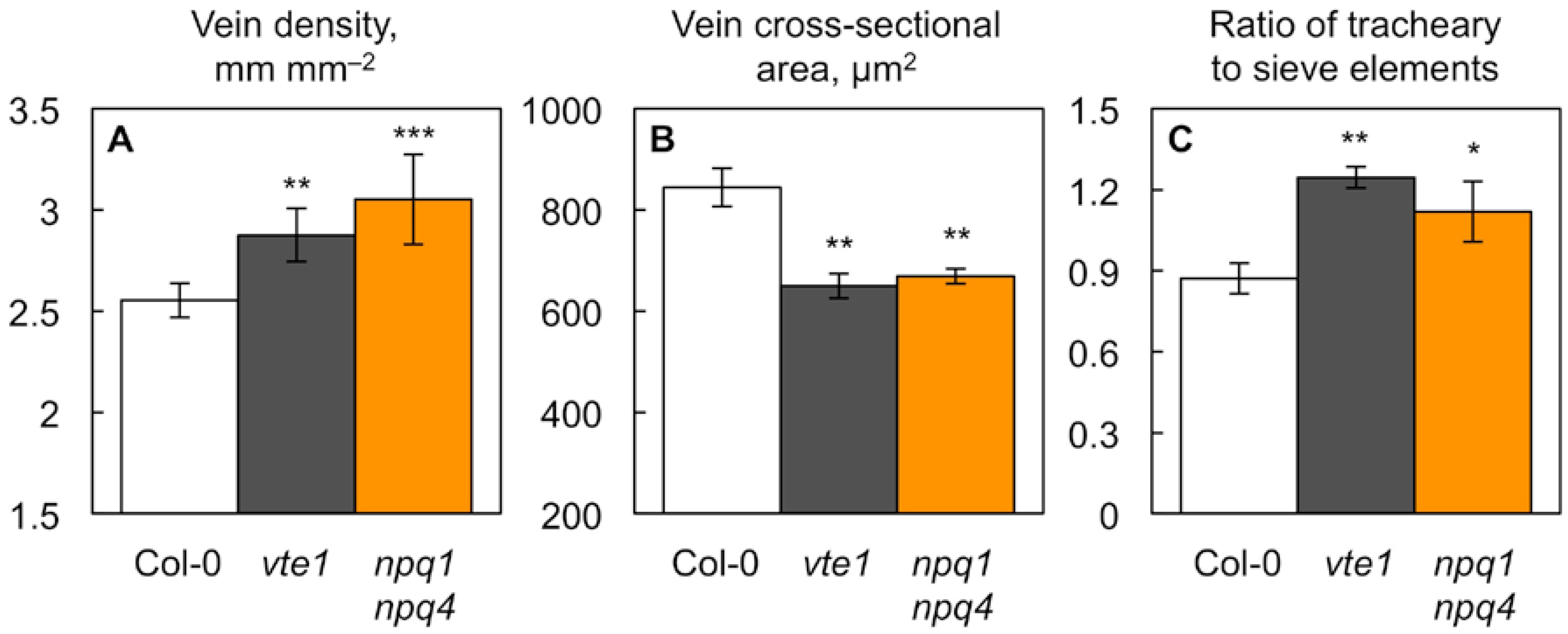
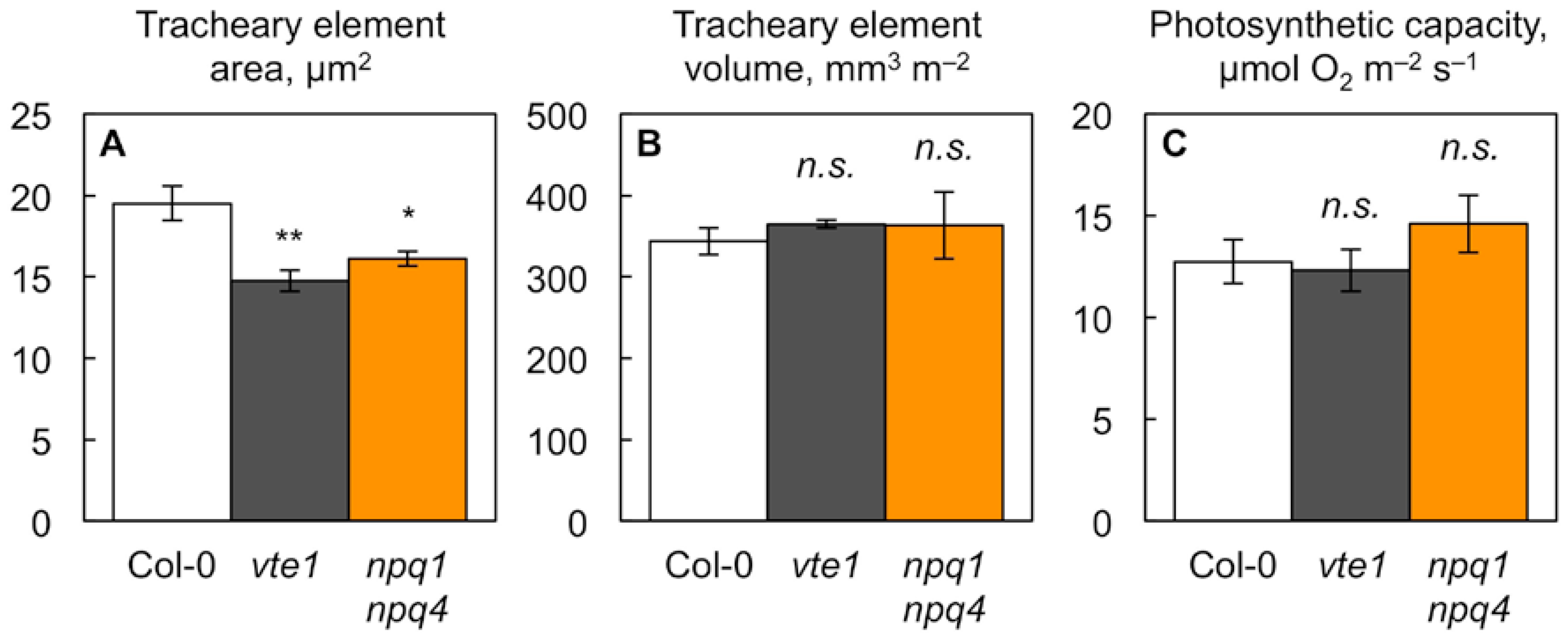
2.2. Impact of Foliar Antioxidant Status on Leaf Vascular Organization
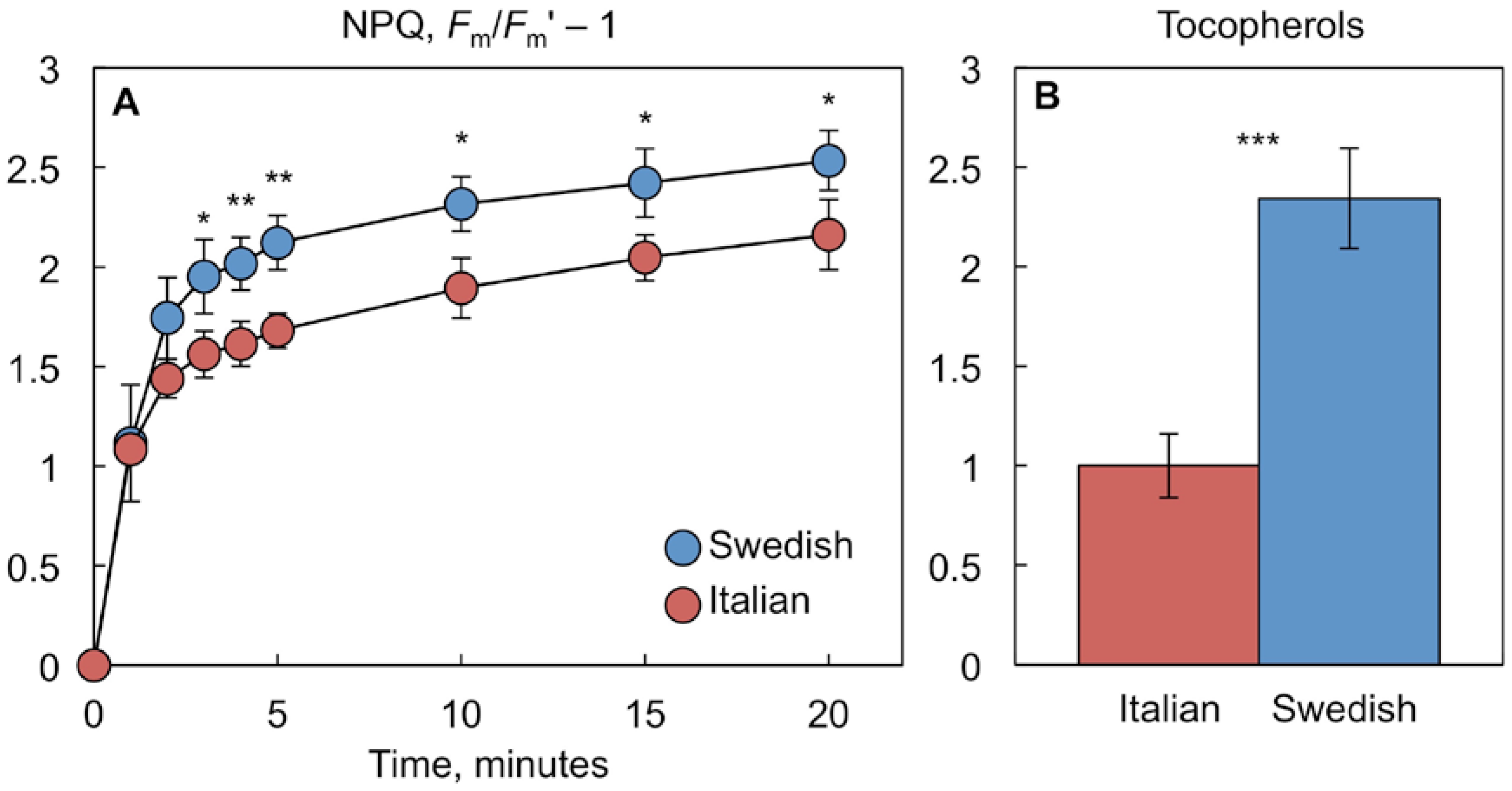
2.3. Comparison of Antioxidant-Deficient Mutants with a Pair of Natural A. thaliana Accessions Differing in NPQ and Tocopherol Levels
3. Discussion
3.1. Vascular Phenotype in Antioxidant Mutants
3.2. Interaction of Redox Signaling Networks with other Regulators
3.3. Comparison of Antioxidant-Deficient Phenotype with Phenotype of Species Adapted to Specific Environments
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Photosynthesis and Chlorophyll Fluorescence
4.3. Pigments
4.4. Minor Vein Anatomy
4.5. Gene Expression
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABRC | Arabidopsis Biological Resource Center |
| cDNA | Complementary deoxyribonucleic acid |
| CBF | C-repeat binding factor |
| Col-0 | Columbia-0 (a wild-type Arabidopsis thaliana line) |
| DREB | Dehydration-responsive element binding factor |
| Fm | Maximal chlorophyll fluorescence determined in leaves darkened for 5 min |
| Fm‘ | Maximal chlorophyll fluorescence determined in a leaf exposed to light |
| MDPI | Multidisciplinary Digital Publishing Institute |
| NCBI | National Center for Biotechnology Information |
| NPQ | Non-photochemical quenching of chlorophyll fluorescence calculated as Fm/Fm‘ − 1 |
| npq1 | An Arabidopsis thaliana mutant line deficient in violaxanthin deepoxidase |
| npq1 npq4 | An Arabidopsis thaliana mutant line deficient in both violaxanthin deepoxidase and PsbS |
| npq4 | An Arabidopsis thaliana mutant line deficient in the PsbS protein |
| n.s. | Not significantly different |
| PsbS | Photosystem II subunit S |
| qPCR | Quantitative polymerase chain reaction |
| RNA | Ribonucleic acid |
| ROS | Reactive oxygen species |
| UBC21 | Gene for ubiquitin-conjugating enzyme E2 21 |
| UTR | Untranslated region |
| VAZ | Violaxanthin + Antheraxanthin + Zeaxanthin |
| vte1 | An Arabidopsis thaliana mutant line deficient in tocopherols |
References
- Dhankher, O.M.; Foyer, C.H. Climate resilient crops for improving global food security and safety. Plant Cell Environ. 2018, 41, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Feild, T.S.; Jordan, G.J. Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiol. 2007, 144, 1890–1898. [Google Scholar] [CrossRef] [PubMed]
- Scoffoni, C.; Chatelet, D.S.; Pasquet-kok, J.; Rawls, M.; Donoghue, M.J.; Edwards, E.J.; Sack, L. Hydraulic basis for the evolution of photosynthetic productivity. Nat. Plants. 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed]
- Hacke, U.G.; Sperry, J.S. Functional and ecological xylem anatomy. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Medek, D.E.; Stuart, S.A.; Pasquet-Kok, J.; Egerton, J.J.G.; Salari, H.; Sack, L.; Ball, M.C. Xylem traits mediate a trade-off between resistance to freeze–thaw-induced embolism and photosynthetic capacity in overwintering evergreens. New Phytol. 2011, 191, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Cohu, C.M.; Muller, O.; Adams, W.W., III; Demmig-Adams, B. Leaf anatomical and photosynthetic acclimation to cool temperature and high light in two winter versus two summer annuals. Physiol. Plant. 2014, 152, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W., III; Stewart, J.J.; Cohu, C.M.; Muller, O.; Demmig-Adams, B. Habitat temperature and precipitation of Arabidopsis thaliana ecotypes determine the response of foliar vasculature, photosynthesis, and transpiration to growth temperature. Front. Plant Sci. 2016, 7, 1026. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J.; Demmig-Adams, B.; Cohu, C.M.; Wenzl, C.A.; Muller, O.; Adams, W.W., III. Growth temperature impact on leaf form and function in Arabidopsis thaliana ecotypes from northern and southern Europe. Plant Cell Environ. 2016, 39, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- Polutchko, S.K.; Stewart, J.J.; Demmig-Adams, B.; Adams, W.W., III. Evaluating the link between photosynthetic capacity and leaf vascular organization with principal component analysis. Photosynthetica 2018, 56, 392–403. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Stewart, J.J.; Baker, C.R.; Adams, W.W., III. Optimization of photosynthetic productivity in contrasting environments by regulons controlling plant form and function. Int. J. Mol. Sci. 2018, 19, 872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bartels, D. Molecular responses to dehydration and desiccation in desiccation-tolerant angiosperm plants. J. Exp. Bot. 2018, 69, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Schippers, J.H.M.; Foyer, C.H.; van Dongen, J.T. Redox regulation in shoot growth, SAM maintenance and flowering. Curr. Opin. Plant Biol. 2016, 29, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Kurepin, L.V.; Dahal, K.P.; Savitch, L.V.; Singh, J.; Bode, R.; Ivanov, A.G.; Hurry, V.; Hüner, N.P.A. Role of CBFs as integrators of chloroplast redox, phytochrome and plant hormone signaling during cold acclimation. Int. J. Mol. Sci. 2013, 14, 12729–12763. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.K.; Gupta, K.; Lopato, S.; Agarwal, P. Dehydration responsive element binding transcription factors and their applications for the engineering of stress tolerance. J. Exp. Bot. 2017, 68, 2135–2148. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, C.G.; Casalongué, C.A.; Simontacchi, M.; Marquez-Garcia, B.; Foyer, C.H. Interactions between hormone and redox signaling pathways in the control of growth and cross tolerance to stress. Environ. Exp. Bot. 2013, 94, 73–88. [Google Scholar] [CrossRef]
- Karpinska, B.; Zhang, K.; Rasool, B.; Pastok, D.; Morris, J.; Verrall, S.R.; Hedley, P.E.; Hancock, R.D.; Foyer, C.H. The redox state of the apoplast influences the acclimation of photosynthesis and leaf metabolism to changing irradiance. Plant Cell Environ. 2017, 41, 1083–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.J.; Adams, W.W., III; Cohu, C.M.; Demmig-Adams, B. Tocopherols modulate leaf vein arrangement and composition without impacting photosynthesis. Photosynthetica 2018, 56, 382–391. [Google Scholar] [CrossRef]
- Havaux, M.; Eymery, F.; Porfirova, S.; Rey, P.; Dörmann, P. Vitamin E protects against photoinhibition and photooxidative stress in Arabidopsis thaliana. Plant Cell 2005, 17, 3451–3469. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Song, W.; Sage, T.L.; DellaPenna, D. Tocopherols play a crucial role in low-temperature adaptation and phloem loading in Arabidopsis. Plant Cell 2006, 18, 2710–2732. [Google Scholar] [CrossRef] [PubMed]
- Ledford, H.K.; Niyogi, K.K. Singlet oxygen and photo-oxidative stress management in plants and algae. Plant Cell Environ. 2005, 28, 1037–1045. [Google Scholar] [CrossRef]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Cohu, C.M.; Amiard, V.; van Zadelhoff, G.; Veldink, G.A.; Muller, O.; Adams, W.W., III. Emerging trade-offs—Impact of photoprotectants (PsbS, xanthophylls, and vitamin E) on oxylipins as regulators of development and defense. New Phytol. 2013, 197, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Niyogi, K.K.; Björkman, O.; Grossman, A.R. Chlamydomonas xanthophyll cycle mutants identified by video imaging of chlorophyll fluorescence quenching. Plant Cell 1997, 9, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Björkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 2000, 403, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Dogra, V.; Rochaix, J.-D.; Kim, C. Singlet oxygen-triggered chloroplast-to-nucleus retrograde signalling pathways: An emerging perspective. Plant Cell Environ. 2018, 41, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Ballottari, M.; Mozzo, M.; Girardon, J.; Hienerwadel, R.; Bassi, R. Chlorophyll triplet quenching and photoprotection in the higher plant monomeric antenna protein Lhcb5. J. Phys. Chem. B 2013, 117, 11337–11348. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, S.; Li, Z.R.; Niyogi, K.K.; Bassi, R.; Dall’Osto, L. The Arabidopsis szl1 mutant reveals a critical role of β-carotene in photosystem I photoprotection. Plant Physiol. 2012, 159, 1745–1758. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J.; Adams, W.W., III; Cohu, C.M.; Polutchko, S.K.; Lombardi, E.M.; Demmig-Adams, B. Differences in light-harvesting, acclimation to growth light environment, and leaf structural development between Swedish and Italian ecotypes of Arabidopsis thaliana. Planta 2015, 242, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Oakley, C.G.; Savage, L.; Lotz, S.; Larson, G.R.; Thomashow, M.F.; Kramer, D.M.; Schemske, D.W. Genetic basis of photosynthetic responses to cold in two locally adapted populations of Arabidopsis thaliana. J. Exp. Bot. 2018, 69, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Gehan, M.A.; Park, S.; Gilmour, S.J.; An, C.; Lee, C.-M.; Thomashow, M.F. Natural variation in the C-repeat binding factor cold response pathway correlates with local adaptation of Arabidopsis ecotypes. Plant J. 2015, 84, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.D.; Sperry, J.S.; Hacke, U.G. The relationship between xylem conduit diameter and cavitation caused by freezing. Am. J. Bot. 1999, 86, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Pittermann, J.; Sperry, J.S. Tracheid diameter is the key trait determining the extent of freezing-induced embolism in conifers. Tree Physiol. 2003, 23, 907–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittermann, J.; Sperry, J.S. Analysis of freeze-thaw embolism in conifers. The interaction between cavitation pressure and tracheid size. Plant Physiol. 2006, 140, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Lens, F.; Tixier, A.; Cochard, H.; Sperry, J.S.; Jansen, S.; Herbette, S. Embolism resistance as a key mechanism to understand adaptive plant strategies. Curr. Opin. Plant Biol. 2013, 16, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Lens, F.; Espino, S.; Karimi, Z.; Klepsch, M.; Schenk, H.J.; Schmitt, M.; Schuldt, B.; Jansen, S. Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem. IAWA J. 2016, 37, 152–171. [Google Scholar] [CrossRef]
- Choat, B.; Cobb, A.R.; Jansen, S. Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function. New Phytol. 2007, 177, 608–626. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.A.; Brodribb, T.J.; Lucani, C.J.; DaMatta, F.M.; McAdam, S.A.M. Coordinated plasticity maintains hydraulic safety in sunflower leaves. Plant Cell Environ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pfautsch, S.; Harbusch, M.; Wesolowski, A.; Smith, R.; Macfarlane, C.; Tjoelker, M.G.; Reich, P.B.; Adams, M.A. Climate determines vascular traits in the ecologically diverse genus Eucalyptus. Ecol. Lett. 2016, 19, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Blackman, C.J.; Brodribb, T.J.; Jordan, G.J. Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol. 2010, 188, 1113–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scoffoni, C.; Albuquerque, C.; Brodersen, C.R.; Townes, S.V.; John, G.P.; Cochard, H.; Buckley, T.N.; McElrone, A.J.; Sack, L. Leaf vein xylem conduit diameter influences susceptibility to embolism and hydraulic decline. New Phytol. 2017, 213, 1076–1092. [Google Scholar] [CrossRef] [PubMed]
- Dunbar-Co, S.; Sporck, M.J.; Sack, L. Leaf trait diversification and design in seven rare taxa of the Hawaiian Plantago radiation. Int. J. Plant Sci. 2009, 170, 61–75. [Google Scholar] [CrossRef]
- Havaux, M.; Niyogi, K.K. The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism. Proc. Natl. Acad. Sci. USA 1999, 96, 8762–8767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havaux, M.; García-Plazaola, J.I. Beyond non-photochemical fluorescence quenching: The overlapping antioxidant functions of zeaxanthin and tocopherols. In Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cyanobacteria, Advances in Photosynthesis and Respiration; Demmig-Adams, B., Garab, G., Adams, W.W., III, Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 40, pp. 583–603. [Google Scholar]
- Hüner, N.P.A.; Bode, R.; Dahal, K.; Hollis, L.; Rosso, D.; Krol, M.; Ivanov, A.G. Chloroplast redox imbalance governs phenotypic plasticity: The “grand design of photosynthesis” revisited. Front. Plant Sci. 2012, 3, 255. [Google Scholar] [CrossRef] [PubMed]
- Hüner, N.P.A.; Dahal, K.; Kurepin, L.V.; Savitch, L.; Singh, J.; Ivanov, A.G.; Kane, K.; Sarhan, F. Potential for increased photosynthetic performance and crop productivity in response to climate change: Role of CBFs and gibberellic acid. Front. Chem. 2014, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Hüner, N.P.A.; Dahal, K.; Bode, R.; Kurepin, L.V.; Ivanov, A.G. Photosynthetic acclimation, vernalization, crop productivity and ‘the grand design of photosynthesis’. J. Plant Physiol. 2016, 203, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Sign. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Phillips, J. Drought stress tolerance. In Genetic Modification of Plants, Biotechnology in Agriculture and Forestry; Kempken, F., Jung, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 64, pp. 139–157. [Google Scholar]
- Cavender-Bares, J.; Cortes, P.; Rambal, S.; Joffre, R.; Miles, B.; Rocheteau, A. Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan. New Phytol. 2005, 168, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W., III; Demmig-Adams, B.; Rosenstiel, T.N.; Ebbert, V. Dependence of photosynthesis and energy dissipation activity upon growth form and light environment during the winter. Photosynth. Res. 2001, 7, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Muller, O.; Stewart, J.J.; Cohu, C.M.; Polutchko, S.K.; Demmig-Adams, B.; Adams, W.W., III. Leaf architectural, vascular and photosynthetic acclimation to temperature in two biennials. Physiol. Plant. 2014, 152, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Sattler, S.E.; Cahoon, E.B.; Coughlan, S.J.; DellaPenna, D. Characterization of tocopherol cyclases from higher plants and cyanobacteria. Evolutionary implications for tocopherol synthesis and function. Plant Physiol. 2003, 132, 2184–2195. [Google Scholar] [CrossRef] [PubMed]
- Niyogi, K.K.; Grossman, A.R.; Björkman, O. Arabidopsis mutants define a central role for the xanthophyll cycle in the regulation of photosynthetic energy conversion. Plant Cell 1998, 10, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J.; Polutchko, S.K.; Adams, W.W., III; Demmig-Adams, B. Acclimation of Swedish and Italian ecotypes of Arabidopsis thaliana to light intensity. Photosynth. Res. 2017, 134, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Ågren, J.; Schemske, D.W. Reciprocal transplants demonstrate strong adaptive differentiation of the model organism Arabidopsis thaliana in its native range. New Phytol. 2012, 194, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Delieu, T.; Walker, D.A. Polarographic measurement of photosynthetic oxygen evolution by leaf discs. New Phytol. 1981, 89, 165–178. [Google Scholar] [CrossRef]
- Demmig, B.; Björkman, O. Comparison of the effect of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Cohu, C.M.; Muller, O.; Demmig-Adams, B.; Adams, W.W., III. Minor loading vein acclimation for three Arabidopsis thaliana ecotypes in response to growth under different temperature and light regimes. Front. Plant Sci. 2013, 4, 240. [Google Scholar] [CrossRef] [PubMed]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Dumlao, M.R.; Darehshouri, A.; Cohu, C.M.; Muller, O.; Mathias, J.; Adams, W.W., III; Demmig-Adams, B. Low temperature acclimation of photosynthetic capacity and leaf morphology in the context of phloem loading type. Photosynth. Res. 2012, 113, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carotenoid Levels | Col-0 | vte1 | npq1 | npq4 | npq1 npq4 |
|---|---|---|---|---|---|
| Zeaxanthin, mmol mol−1 Chl | 35 ± 1 | 35 ± 1 | 2.1 ± 0.5 *** | 35 ± 3 | 2.1 ± 0.3 *** |
| Zeaxanthin, % of VAZ pool | 65 ± 1 | 64 ± 2 | 3.9 ± 0.9 *** | 63 ± 3 | 4.0 ± 0.7 *** |
| Lutein, mmol mol−1 Chl | 142 ± 1 | 145 ± 4 | 142 ± 0 | 142 ± 2 | 142 ± 0 |
| β-carotene, mmol mol−1 Chl | 83 ± 5 | 82 ± 3 | 87 ± 2 | 86 ± 2 | 83 ± 5 |
| Vascular Features | Col-0 | vte1 | npq1 npq4 |
|---|---|---|---|
| Tracheary elements per minor vein | 6.8 ± 0.4 | 8.6 ± 0.5 * | 7.2 ± 0.5 |
| Tracheary elements per minor vein × vein density | 17 ± 1 | 25 ± 1 ** | 22 ± 1 * |
| Sieve elements per minor vein | 7.8 ± 0.0 | 7.0 ± 0.1 * | 6.6 ± 0.4 ** |
| Sieve elements per minor vein × vein density | 20 ± 0 | 20 ± 0 | 20 ± 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stewart, J.J.; Baker, C.R.; Sharpes, C.S.; Wong-Michalak, S.T.; Polutchko, S.K.; Adams, W.W., III; Demmig-Adams, B. Effects of Foliar Redox Status on Leaf Vascular Organization Suggest Avenues for Cooptimization of Photosynthesis and Heat Tolerance. Int. J. Mol. Sci. 2018, 19, 2507. https://doi.org/10.3390/ijms19092507
Stewart JJ, Baker CR, Sharpes CS, Wong-Michalak ST, Polutchko SK, Adams WW III, Demmig-Adams B. Effects of Foliar Redox Status on Leaf Vascular Organization Suggest Avenues for Cooptimization of Photosynthesis and Heat Tolerance. International Journal of Molecular Sciences. 2018; 19(9):2507. https://doi.org/10.3390/ijms19092507
Chicago/Turabian StyleStewart, Jared J., Christopher R. Baker, Carlie S. Sharpes, Shannon Toy Wong-Michalak, Stephanie K. Polutchko, William W. Adams, III, and Barbara Demmig-Adams. 2018. "Effects of Foliar Redox Status on Leaf Vascular Organization Suggest Avenues for Cooptimization of Photosynthesis and Heat Tolerance" International Journal of Molecular Sciences 19, no. 9: 2507. https://doi.org/10.3390/ijms19092507