Pulsed Electromagnetic Fields Increase Pigmentation through the p-ERK/p-p38 Pathway in Zebrafish (Danio rerio)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
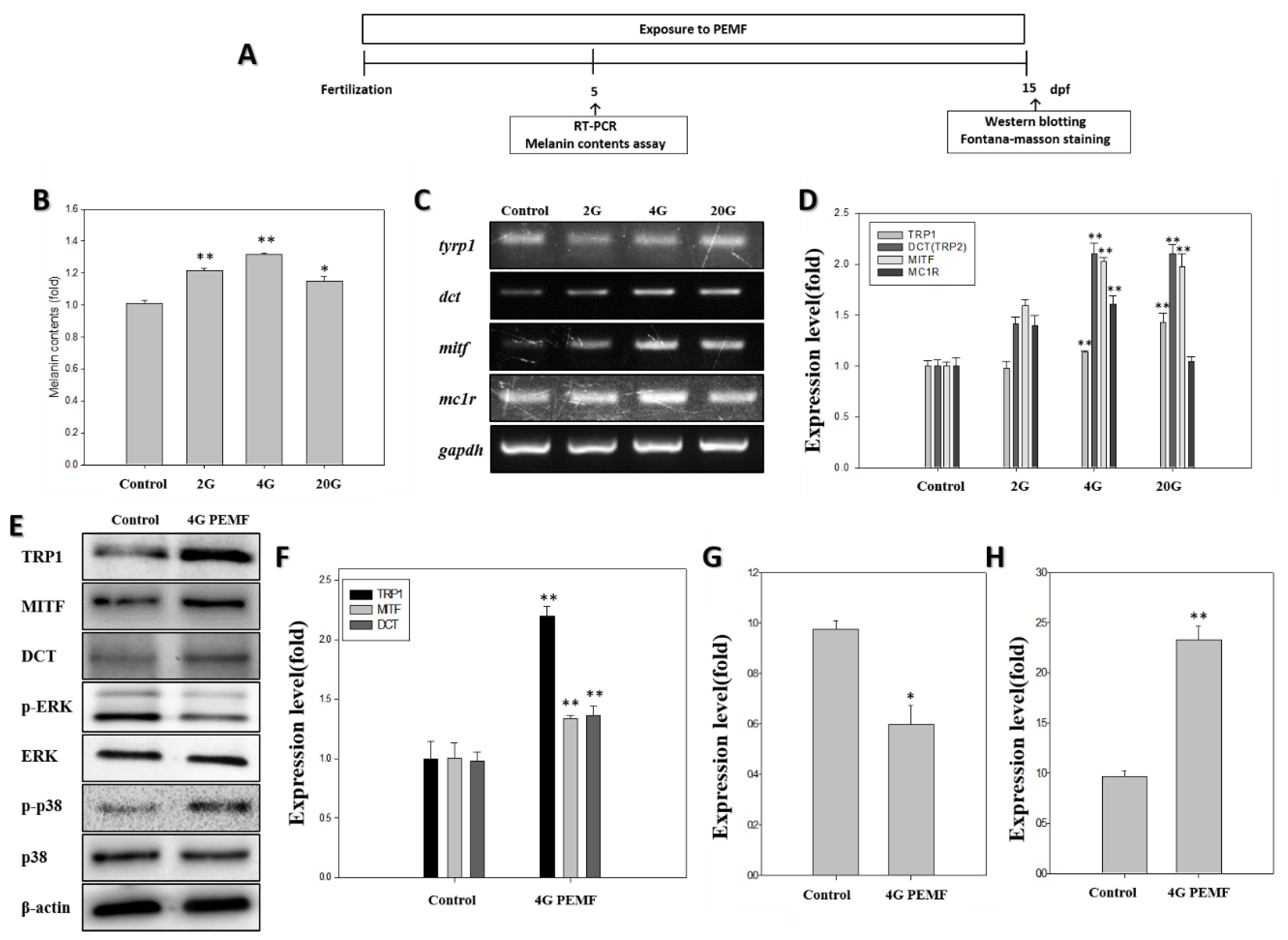
2. Results
2.1. Melanin Assay
2.2. RT-PCR
2.3. Western Blotting
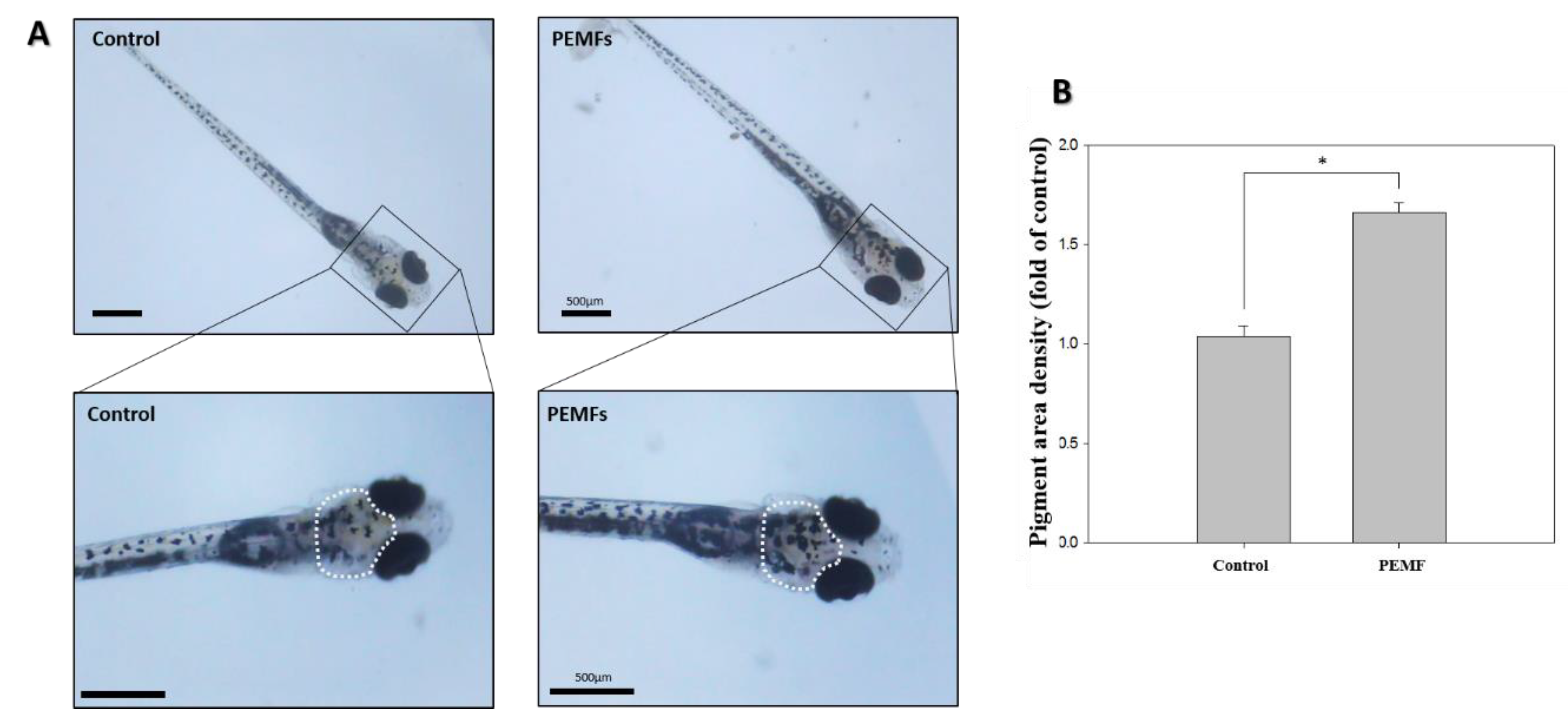
2.4. Pigmentation
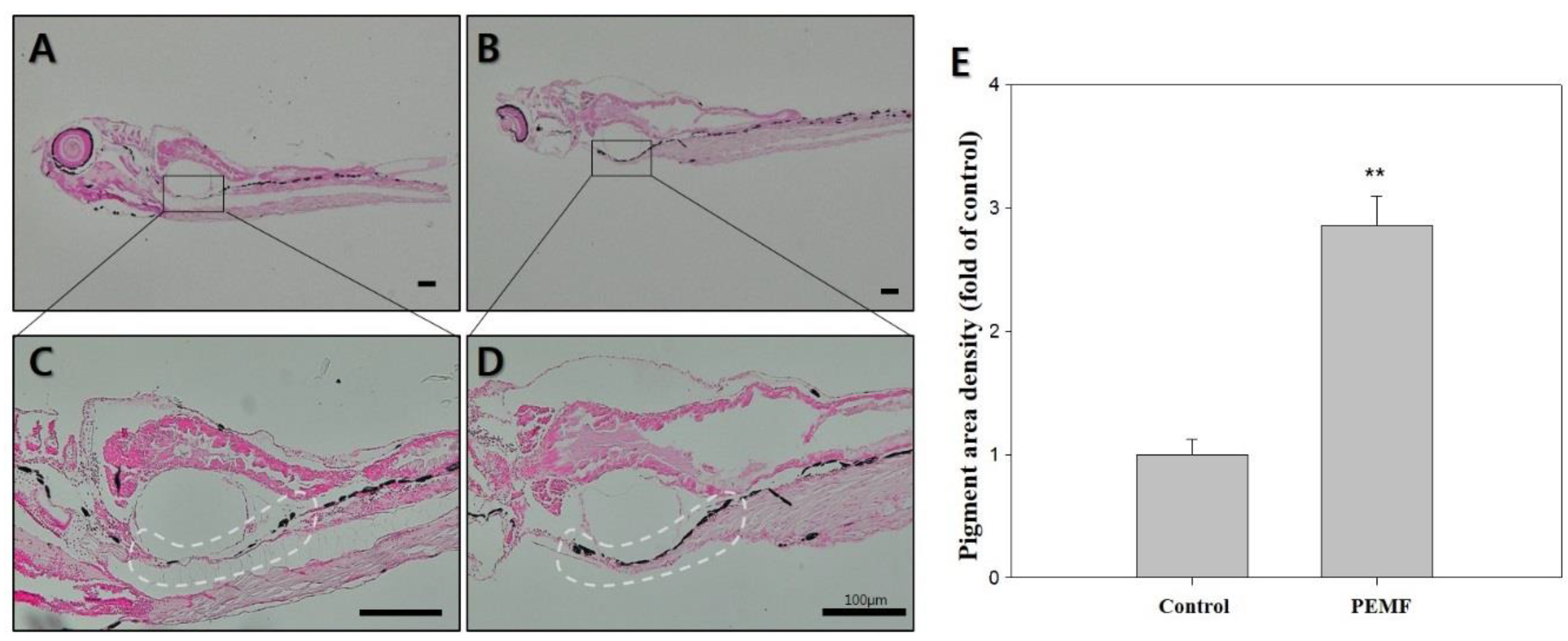
2.5. Fontana-Masson Staining
3. Discussion
4. Materials and Methods
4.1. Zebrafish Maintenance and Embryo Collection
4.2. PEMFs Exposure
4.3. Microscopy
4.4. Melanin Assay
4.5. Reverse Transcriptase-PCR Analysis
4.6. Western Blotting
4.7. Fontana-Masson Silver Staining
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jung, G.D.; Yang, J.Y.; Song, E.S.; Par, J.W. Stimulation of melanogenesis by glycyrrhizin in B16 melanoma cells. Exp. Mol. Med. 2001, 33, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.Y.; Koo, J.H.; Song, Y.G.; Kwon, K.B.; Lee, J.H.; Sohn, H.S.; Park, B.H.; Jhee, E.C.; Park, J.W. Stimulation of melanogenesis by scoparone in B16 melanoma cells. Acta Pharmacol. Sin. 2006, 27, 1467–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lommerts, J.E.; Uitentuis, S.E.; Bekkenk, M.W.; de Rie, M.A.; Wolkerstorfer, A. The role of phototherapy in the surgical treatment of vitiligo: A systematic review. J. Eur. Acad. Dermatol. Venereol. JEADV 2018, 32, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Lurz, C.; Hamm, H.; Brocker, E.B.; Hofmann, U.B. Narrow-band UVB311 nm vs. broad-band UVB therapy in combination with topical calcipotriol vs. placebo in vitiligo. Int. J. Dermatol. 2005, 44, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Fritz, K.; Salavastru, C. [The 308 nm Excimer laser:Treatment of vitiligo and hypopigmentation]. Der Hautarzt Zeitschrift fur Dermatologie Venerologie und verwandte Gebiete 2018, 69, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Hann, S.K.; Oh, S.H. Combination treatment with excimer laser and narrowband UVB light in vitiligo patients. Photodermatol. Photoimmunol. Photomed. 2016, 32, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Lotti, T.; Franca, K. UVA 1 laser in the treatment of patients with vitiligo. Dermatol. Ther. 2017, 30. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.W.; Babitha, S.; Kim, E.K.; Shin, J. A successful helium-neon laser and topical tacrolimus combination therapy in one child with vitiligo. Dermatol. Ther. 2015, 28, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Majid, I.; Imran, S. Depigmentation Therapy with Q-Switched Nd: YAG Laser in Universal Vitiligo. J. Cutan. Aesthet. Surg. 2013, 6, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Njoo, M.D.; Vodegel, R.M.; Westerhof, W. Depigmentation therapy in vitiligo universalis with topical 4-methoxyphenol and the Q-switched ruby laser. J. Am. Acad. Dermatol. 2000, 42, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Chung, B.S.; Choi, K.C. Depigmentation therapy with Q-switched ruby laser after tanning in vitiligo universalis. Dermatol. Surg. 2001, 27, 969–970. [Google Scholar] [PubMed]
- Yuan, J.; Xin, F.; Jiang, W. Underlying Signaling Pathways and Therapeutic Applications of Pulsed Electromagnetic Fields in Bone Repair. Cell. Physiol. Biochem. 2018, 46, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Benazzo, F.; Cadossi, M.; Cavani, F.; Fini, M.; Giavaresi, G.; Setti, S.; Cadossi, R.; Giardino, R. Cartilage repair with osteochondral autografts in sheep: Effect of biophysical stimulation with pulsed electromagnetic fields. J. Orthop. Res. 2008, 26, 631–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weintraub, M.I.; Herrmann, D.N.; Smith, A.G.; Backonja, M.M.; Cole, S.P. Pulsed electromagnetic fields to reduce diabetic neuropathic pain and stimulate neuronal repair: A randomized controlled trial. Arch. Phys. Med. Rehab. 2009, 90, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Leone, L.; Fusco, S.; Mastrodonato, A.; Piacentini, R.; Barbati, S.A.; Zaffina, S.; Pani, G.; Podda, M.V.; Grassi, C. Epigenetic modulation of adult hippocampal neurogenesis by extremely low-frequency electromagnetic fields. Mol. Neurobiol. 2014, 49, 1472–1486. [Google Scholar] [CrossRef] [PubMed]
- Greenough, C.G. The effects of pulsed electromagnetic fields on blood vessel growth in the rabbit ear chamber. J. Orthop. Res. 1992, 10, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Romero-Sierra, C.; Halter, S.; Tanner, J.A.; Roomi, M.W.; Crabtree, D. Electromagnetic fields and skin wound repair. J. Microw. Power 1975, 10, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-E.; Kim, Y.-M.; Kang, K.-H.; Kim, S.-C.; Park, J.-K.; Seo, Y.-K. Pigmentation effect of electromagnetic fields at various intensities to melanocytes. Tissue Eng. Regen. Med. 2016, 13, 560–567. [Google Scholar] [CrossRef]
- Kim, Y.M.; Cho, S.E.; Kim, S.C.; Jang, H.J.; Seo, Y.K. Effects of Extremely Low Frequency Electromagnetic Fields on Melanogenesis through p-ERK and p-SAPK/JNK Pathways in Human Melanocytes. Int. J. Mol. Sci. 2017, 18, 2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Cho, S.E.; Seo, Y.K. The activation of melanogenesis by p-CREB and MITF signaling with extremely low-frequency electromagnetic fields on B16F10 melanoma. Life Sci. 2016, 162, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Azuma, N.; Duzgun, S.A.; Ikeda, M.; Kito, H.; Akasaka, N.; Sasajima, T.; Sumpio, B.E. Endothelial cell response to different mechanical forces. J. Vasc. Surg. 2000, 32, 789–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.S.; Kim, Y.J.; Kim, M.O.; Kang, M.; Oh, S.W.; Nho, Y.H.; Park, S.H.; Lee, J. Cannabidiol upregulates melanogenesis through CB1 dependent pathway by activating p38 MAPK and p42/44 MAPK. Chem.-Biol. Interact. 2017, 273, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Nishina, A.; Miura, A.; Goto, M.; Terakado, K.; Sato, D.; Kimura, H.; Hirai, Y.; Sato, H.; Phay, N. Mansonone E from Mansonia gagei Inhibited alpha-MSH-Induced Melanogenesis in B16 Cells by Inhibiting CREB Expression and Phosphorylation in the PI3K/Akt Pathway. Biol. Pharm. Bull. 2018, 41, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lee, J.; Jeong, Y.T.; Byun, G.H.; Kim, J.H. Melanogenesis inhibition activity of floralginsenoside A from Panax ginseng berry. J. Ginseng Res. 2017, 41, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Yang, H.M.; Kang, S.M.; Ahn, G.N.; Roh, S.W.; Lee, W.; Kim, D.K.; Jeon, Y.J. Whitening Effect of Octaphlorethol A Isolated from Ishige foliacea in an In Vivo Zebrafish Model. J. Microbiol. Biotechnol. 2015, 25, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.Y.; Kim, J.H.; Ko, D.H.; Kim, C.H.; Hwang, J.S.; Ahn, S.; Kim, S.Y.; Kim, C.D.; Lee, J.H.; Yoon, T.J. Zebrafish as a new model for phenotype-based screening of melanogenic regulatory compounds. Pigment Cell Res. 2007, 20, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Preece, A.W.; Kaune, W.; Grainger, P.; Preece, S.; Golding, J. Magnetic fields from domestic appliances in the UK. Phys. Med. Biol. 1997, 42, 67–76. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, C.; Costantini, E.; Kamal, M.A.; Reale, M. Experimental model for ELF-EMF exposure: Concern for human health. Saudi J. Biol. Sci. 2015, 22, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Chen, C.; Deng, P.; Zhu, G.; Lin, M.; Zhang, L.; Xu, S.; He, M.; Lu, Y.; Duan, W.; et al. Extremely Low-Frequency Electromagnetic Fields Promote In Vitro Neuronal Differentiation and Neurite Outgrowth of Embryonic Neural Stem Cells via Up-Regulating TRPC1. PLoS ONE 2016, 11, e0150923. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dai, Y.; Zhu, X.; Xu, H.; Cai, P.; Xia, R.; Mao, L.; Zhao, B.Q.; Fan, W. Extremely low-frequency electromagnetic fields enhance the proliferation and differentiation of neural progenitor cells cultured from ischemic brains. Neuroreport 2015, 26, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Destefanis, M.; Viano, M.; Leo, C.; Gervino, G.; Ponzetto, A.; Silvagno, F. Extremely low frequency electromagnetic fields affect proliferation and mitochondrial activity of human cancer cell lines. Int. J. Radiat. Biol. 2015, 91, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Podda, M.V.; Leone, L.; Barbati, S.A.; Mastrodonato, A.; Li Puma, D.D.; Piacentini, R.; Grassi, C. Extremely low-frequency electromagnetic fields enhance the survival of newborn neurons in the mouse hippocampus. Eur. J. Neurosci. 2014, 39, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, W.B.; Liu, K.J.; Ao, L.; Cao, J.; Zhong, J.L.; Liu, J.Y. Overexpression of miR-26b-5p regulates the cell cycle by targeting CCND2 in GC-2 cells under exposure to extremely low frequency electromagnetic fields. Cell Cycle 2016, 15, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-S.; Kuan, Y.-D.; Chiu, K.-H.; Wang, W.-K.; Chang, F.-H.; Liu, C.-H.; Lee, C.-H. The extract of Rhodobacter sphaeroides inhibits melanogenesis through the MEK/ERK signaling pathway. Mar. Drugs 2013, 11, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Regazzetti, C.; De Donatis, G.M.; Ghorbel, H.H.; Cardot-Leccia, N.; Ambrosetti, D.; Bahadoran, P.; Chignon-Sicard, B.; Lacour, J.P.; Ballotti, R.; Mahns, A.; et al. Endothelial Cells Promote Pigmentation through Endothelin Receptor B Activation. J. Investig. Dermatol. 2015, 135, 3096–3104. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.; Lundegaard, P.R.; Reynolds, N.L.; Dorin, J.R.; Porteous, D.J.; Jackson, I.J.; Patton, E.E. mc1r Pathway regulation of zebrafish melanosome dispersion. Zebrafish 2008, 5, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Jin, E.J.; Thibaudeau, G. Effects of lithium on pigmentation in the embryonic zebrafish. Biochim. Biophys. Acta 1999, 1449, 93–99. [Google Scholar] [CrossRef]
- Jang, J.Y.; Kim, H.N.; Kim, Y.R.; Choi, W.Y.; Choi, Y.H.; Shin, H.K.; Choi, B.T. Partially purified components of Nardostachys chinensis suppress melanin synthesis through ERK and Akt signaling pathway with cAMP down-regulation in B16F10 cells. J. Ethnopharmacol. 2011, 137, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Jin, C.L.; Oh, J.H.; Oh, I.G.; Park, C.H.; Chung, J.H. Ardisia crenata extract stimulates melanogenesis in B16F10 melanoma cells through inhibiting ERK1/2 and Akt activation. Mol. Med. Rep. 2015, 11, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Wang, H.; Chu, J.H.; Chou, G.X.; Yu, Z.L. Activation of p38 MAPK pathway contributes to the melanogenic property of apigenin in B16 cells. Exp. Dermatol. 2011, 20, 755–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Cui, J.; Zhu, H. Suppression of type I collagen in human scleral fibroblasts treated with extremely low-frequency electromagnetic fields. Mol. Vis. 2013, 19, 885–893. [Google Scholar] [PubMed]
- Xu, L.; Xu, Q.H.; Zhou, X.Y.; Yin, L.Y.; Guan, P.P.; Zhang, T.; Liu, J.X. Mechanisms of silver_nanoparticles induced hypopigmentation in embryonic zebrafish. Aquat. Toxicol. 2017, 184, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Lee, S.H. Sesamol decreases melanin biosynthesis in melanocyte cells and zebrafish: Possible involvement of MITF via the intracellular cAMP and p38/JNK signalling pathways. Exp. Dermatol. 2015, 24, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.-Y.; Seo, Y.-K.; Park, J.-M.; Seo, M.-J.; Park, J.-K.; Kim, J.-W.; Park, C.-S. Fermented rice bran downregulates MITF expression and leads to inhibition of α-MSH-induced melanogenesis in B16F1 melanoma. Biosci. Biotechnol. Biochem. 2009, 73, 1704–1710. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-M.; Lim, H.-M.; Ro, H.-S.; Ki, G.-E.; Seo, Y.-K. Pulsed Electromagnetic Fields Increase Pigmentation through the p-ERK/p-p38 Pathway in Zebrafish (Danio rerio). Int. J. Mol. Sci. 2018, 19, 3211. https://doi.org/10.3390/ijms19103211
Kim Y-M, Lim H-M, Ro H-S, Ki G-E, Seo Y-K. Pulsed Electromagnetic Fields Increase Pigmentation through the p-ERK/p-p38 Pathway in Zebrafish (Danio rerio). International Journal of Molecular Sciences. 2018; 19(10):3211. https://doi.org/10.3390/ijms19103211
Chicago/Turabian StyleKim, Yu-Mi, Han-Moi Lim, Hyang-Seon Ro, Ga-Eun Ki, and Young-Kwon Seo. 2018. "Pulsed Electromagnetic Fields Increase Pigmentation through the p-ERK/p-p38 Pathway in Zebrafish (Danio rerio)" International Journal of Molecular Sciences 19, no. 10: 3211. https://doi.org/10.3390/ijms19103211