Downregulation of C-Terminal Tensin-Like Protein (CTEN) Suppresses Prostate Cell Proliferation and Contributes to Acinar Morphogenesis


Abstract
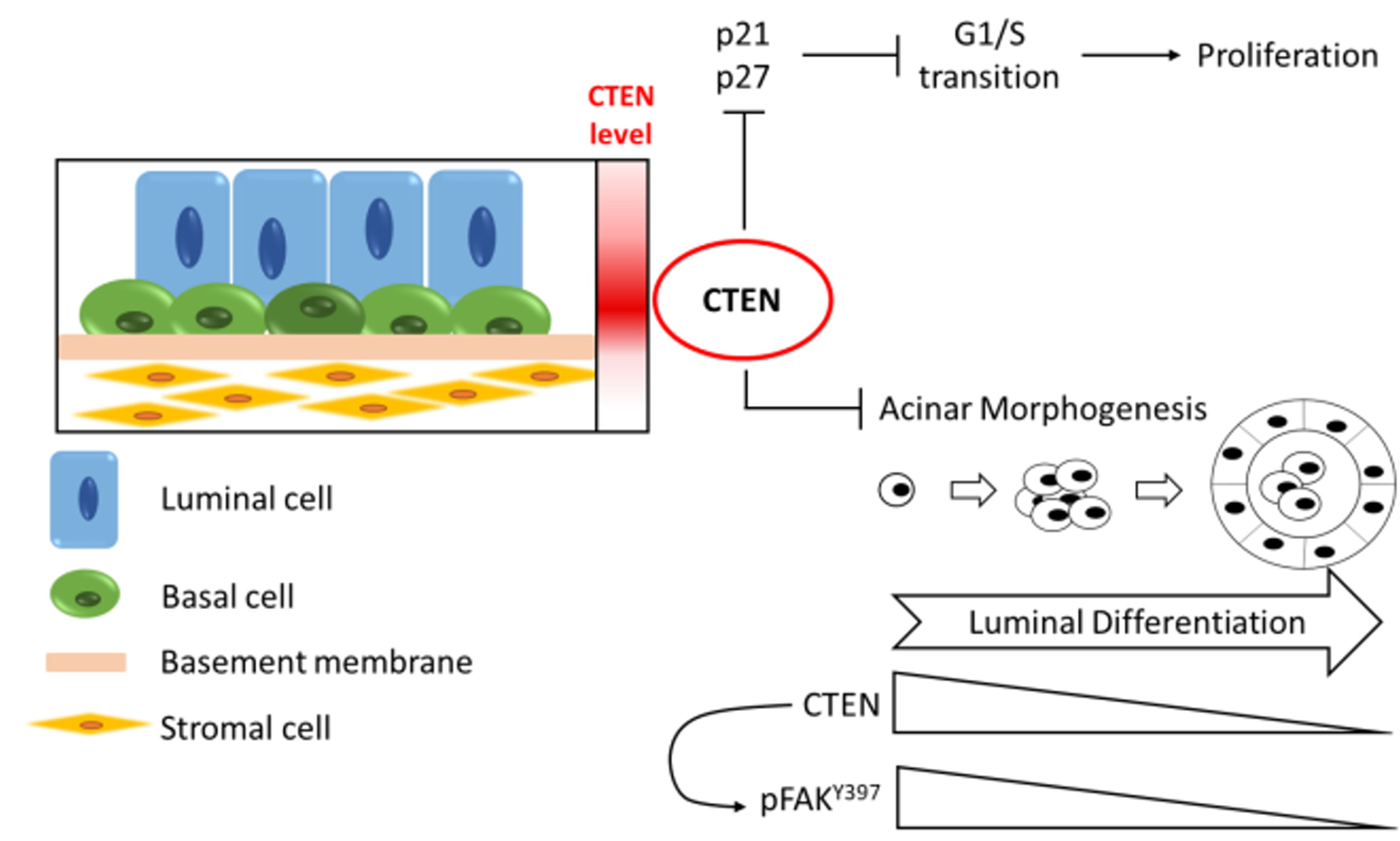
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. CTEN Is Highly Expressed in Prostate Basal Epithelial Cells
2.2. Depletion of CTEN Attenuates Prostate Cell Proliferation
2.3. Downregulation of CTEN Is Required for Normal Acinar Formation
2.4. CTEN Interacts with Integrin β1 and Increases the Activity of FAK and RhoA
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Bioinformatics Analyses
4.3. RNA Interference
4.4. Cell Proliferation Assay
4.5. Cell Cycle Analyses
4.6. Constructions of Lentiviral Expression System
4.7. Acinar Morphogenesis Assay
4.8. Western Analyses
4.9. Immunofluorescence Microscopy
4.10. Coimmunoprecipitation Assay
4.11. GTP-RhoA Pull-Down Assay
4.12. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three dimensional |
| AR | Androgen receptor |
| BPH | Benign prostatic hyperplasia |
| CDK | Cyclin-dependent kinase |
| CK18 | Cytokeratin 18 |
| CK5 | Cytokeratin 5 |
| CK8 | Cytokeratin 8 |
| CTEN | C-terminal tensin-like protein |
| EGFP | Enhanced green fluorescent protein |
| ERK | extracellular-signal regulated kinase |
| FAK | Focal adhesion kinase |
| GEO | Gene Expression Omnibus |
| MAPK | mitogen-activated protein kinase |
| PrEC | Prostate epithelial cell |
| PrSC | Prostate stromal cell |
| PrSMC | Prostate smooth muscle cell |
| PSA | Prostate-specific antigen |
| PSCA | Prostate stem cell antigen |
| PTB | Phosphotyrosine-binding |
| SH2 | Src homology 2 |
| rEGF | Recombinant epidermal growth factor |
References
- Toivanen, R.; Shen, M.M. Prostate organogenesis: Tissue induction, hormonal regulation and cell type specification. Development 2017, 144, 1382–1398. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.B.; Miranti, C.K. Disruption of prostate epithelial differentiation pathways and prostate cancer development. Front. Oncol. 2013, 3, 273. [Google Scholar] [CrossRef] [PubMed]
- Packer, J.R.; Maitland, N.J. The molecular and cellular origin of human prostate cancer. Biochim. Biophys. Acta 2016, 1863, 1238–1260. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.H.; Lo, T.B. Cten, a COOH-terminal tensin-like protein with prostate restricted expression, is down-regulated in prostate cancer. Cancer Res. 2002, 62, 4217–4221. [Google Scholar] [PubMed]
- Chen, N.-T.; Kuwabara, Y.; Conley, C.; Liao, Y.-C.; Hong, S.-Y.; Chen, M.; Shih, Y.-P.; Chen, H.-W.; Hsieh, F.; Lo, S.H. Phylogenetic analysis, expression patterns, and transcriptional regulation of human CTEN gene. Gene 2013, 520, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Wu, W.-M.; Chen, Y.-C.; Lo, S.H.; Liao, Y.-C. ΔNp63α transcriptionally regulates the expression of CTEN that is associated with prostate cell adhesion. PLoS ONE 2016, 11, e0147542. [Google Scholar] [CrossRef] [PubMed]
- Signoretti, S.; Waltregny, D.; Dilks, J.; Isaac, B.; Lin, D.; Garraway, L.; Yang, A.; Montironi, R.; McKeon, F.; Loda, M. p63 is a prostate basal cell marker and is required for prostate development. Am. J. Pathol. 2000, 157, 1769–1775. [Google Scholar] [CrossRef]
- Kurita, T.; Medina, R.T.; Mills, A.A.; Cunha, G.R. Role of p63 and basal cells in the prostate. Development 2004, 131, 4955–4964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signoretti, S.; Pires, M.M.; Lindauer, M.; Horner, J.W.; Grisanzio, C.; Dhar, S.; Majumder, P.; McKeon, F.; Kantoff, P.W.; Sellers, W.R.; et al. p63 regulates commitment to the prostate cell lineage. Proc. Natl. Acad. Sci. USA 2005, 102, 11355–11360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.; Le Magnen, C.; Mitrofanova, A.; Ouyang, X.S.; Califano, A.; Abate-Shen, C. Identification of an NKX3.1-G9a-UTY transcriptional regulatory network that controls prostate differentiation. Science 2016, 352, 1576–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia-Gaur, R.; Donjacour, A.A.; Sciavolino, P.J.; Kim, M.; Desai, N.; Young, P.; Norton, C.R.; Gridley, T.; Cardiff, R.D.; Cunha, G.R.; et al. Roles for Nkx3.1 in prostate development and cancer. Genes Dev. 1999, 13, 966–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Mutton, L.N.; Prins, G.S.; Bieberich, C.J. Distinct regulatory elements mediate the dynamic expression pattern of Nkx3.1. Dev. Dyn. 2005, 234, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kruithof-de Julio, M.; Economides, K.D.; Walker, D.; Yu, H.L.; Halili, M.V.; Hu, Y.P.; Price, S.M.; Abate-Shen, C.; Shen, M.M. A luminal epithelial stem cell that is a cell of origin for prostate cancer. Nature 2009, 461, 495–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruithof-de Julio, M.; Shibata, M.; Desai, N.; Reynon, M.; Halili, M.V.; Hu, Y.P.; Price, S.M.; Abate-Shen, C.; Shen, M.M. Canonical Wnt signaling regulates Nkx3.1 expression and luminal epithelial differentiation during prostate organogenesis. Dev. Dyn. 2013, 242, 1160–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, D.A.; Xin, L.; Lukacs, R.U.; Cheng, D.H.; Witte, O.N. Isolation and functional characterization of murine prostate stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Webber, M.M.; Bello, D.; Kleinman, H.K.; Hoffman, M.P. Acinar differentiation by non-malignant immortalized human prostatic epithelial cells and its loss by malignant cells. Carcinogenesis 1997, 18, 1225–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webber, M.M.; Quader, S.T.A.; Kleinman, H.K.; Bello-DeOcampo, D.; Storto, P.D.; Bice, G.; DeMendonca-Calaca, W.; Williams, D.E. Human cell lines as an in vitro/in vivo model for prostate carcinogenesis and progression. Prostate 2001, 47, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bello-DeOcampo, D.; Kleinman, H.K.; Deocampo, N.D.; Webber, M.M. Laminin-1 and α6β1 integrin regulate acinar morphogenesis of normal and malignant human prostate epithelial cells. Prostate 2001, 46, 142–153. [Google Scholar] [CrossRef]
- Tyson, D.R.; Inokuchi, J.; Tsunoda, T.; Lau, A.; Ornstein, D.K. Culture requirements of prostatic epithelial cell lines for acinar morphogenesis and lumen formation in vitro: Role of extracellular calcium. Prostate 2007, 67, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; Kawano, Y.; Bengoa, N.; Walker, M.M.; Maltry, N.; Niehrs, C.; Waxman, J.; Kypta, R. Downregulation of Dickkopf-3 disrupts prostate acinar morphogenesis through TGF-β/Smad signalling. J. Cell Sci. 2013, 126, 1858–1867. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.; Amit, I.; Citri, A.; Shay, T.; Carvalho, S.; Lavi, S.; Milanezi, F.; Lyass, L.; Amariglio, N.; Jacob-Hirsch, J.; et al. A reciprocal tensin-3-cten switch mediates EGF-driven mammary cell migration. Nat. Cell Biol. 2007, 9, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Streuli, C.H. An integrin-ILK-microtubule network orients cell polarity and lumen formation in glandular epithelium. Nat. Cell Biol. 2013, 15, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.Y.; Jin, S.-P.; Kim, Y.K.; Lee, H.; Han, S.; Lee, D.H.; Chung, J.H. Integrin-β4–TNS4–Focal adhesion kinase signaling mediates keratinocyte proliferation in human skin. J. Investig. Dermatol. 2017, 137, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Lakshman, N.; Kim, A.; Bayless, K.J.; Davis, G.E.; Petroll, W.M. Rho plays a central role in regulating local cell-matrix mechanical interactions in 3D culture. Cell Motil. Cytoskeleton 2007, 64, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Deevi, R.K.; Cox, O.T.; O’Connor, R. Essential function for PDLIM2 in cell polarization in three-dimensional cultures by feedback regulation of the β1-integrin-RhoA signaling axis. Neoplasia 2014, 16, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Ihrie, R.A.; Marques, M.R.; Nguyen, B.T.; Horner, J.S.; Papazoglu, C.; Bronson, R.T.; Mills, A.A.; Attardi, L.D. Perp is a p63-regulated gene essential for epithelial integrity. Cell 2005, 120, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D.K.; Carroll, J.S.; Leong, C.O.; Cheng, F.; Brown, M.; Mills, A.A.; Brugge, J.S.; Ellisen, L.W. p63 regulates an adhesion programme and cell survival in epithelial cells. Nat. Cell Biol. 2006, 8, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Clements, S.E.; Techanukul, T.; Lai-Cheong, J.E.; Mee, J.B.; South, A.P.; Pourreyron, C.; Burrows, N.P.; Mellerio, J.E.; McGrath, J.A. Mutations in AEC syndrome skin reveal a role for p63 in basement membrane adhesion, skin barrier integrity and hair follicle biology. Br. J. Dermatol. 2012, 167, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Muharram, G.; Sahgal, P.; Korpela, T.; De Franceschi, N.; Kaukonen, R.; Clark, K.; Tulasne, D.; Carpén, O.; Ivaska, J. Tensin-4-dependent MET stabilization is essential for survival and proliferation in carcinoma cells. Dev. Cell 2014, 29, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Aratani, K.; Komatsu, S.; Ichikawa, D.; Ohashi, T.; Miyamae, M.; Okajima, W.; Imamura, T.; Kiuchi, J.; Nishibeppu, K.; Kosuga, T.; et al. Overexpression of CTEN relates to tumor malignant potential and poor outcomes of adenocarcinoma of the esophagogastric junction. Oncotarget 2017, 8, 84112–84122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wade Harper, J.; Adami, G.R.; Wei, N.; Keyomarsi, K.; Elledge, S.J. The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef]
- Toyoshima, H.; Hunter, T. p27, a novel inhibitor of G1 cyclin-Cdk protein kinase activity, is related to p21. Cell 1994, 78, 67–74. [Google Scholar] [CrossRef]
- Besson, A.; Dowdy, S.F.; Roberts, J.M. CDK Inhibitors: Cell Cycle Regulators and Beyond. Dev. Cell 2008, 14, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Bruni-Cardoso, A.; Carvalho, H.F. Dynamics of the epithelium during canalization of the rat ventral prostate. Anat. Rec. 2007, 290, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Bonkhoff, H.; Wernert, N.; Dhom, G.; Remberger, K. Relation of endocrine-paracrine cells to cell proliferation in normal, hyperplastic, and neoplastic human prostate. Prostate 1991, 19, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Bonkhoff, H.; Stein, U.; Remberger, K. The proliferative function of basal cells in the normal and hyperplastic human prostate. Prostate 1994, 24, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Hudson, D.L.; Guy, A.T.; Fry, P.; O’Hare, M.J.; Watt, F.M.; Masters, J.R.W. Epithelial cell differentiation pathways in the human prostate: Identification of intermediate phenotypes by keratin Expression. J. Histochem. Cytochem. 2001, 49, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.; Foster, F.; Wood, A.; Owens, T.; Brennan, K.; Streuli, C.H.; Gilmore, A.P. Oncogenic activation of FAK drives apoptosis suppression in a 3D-culture model of breast cancer initiation. Oncotarget 2016, 7, 70336–70352. [Google Scholar] [CrossRef] [PubMed]
- McNeal, J.E. Normal histology of the prostate. Am. J. Surg. Pathol. 1988, 12, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Sugimura, Y.; Kawamura, J.; Donjacour, A.A.; Cunha, G.R. Morphological and functional heterogeneity in the rat prostatic gland. Biol. Reprod. 1991, 45, 308–321. [Google Scholar] [CrossRef] [PubMed]
- El-Alfy, M.; Pelletier, G.; Hermo, L.S.; Labrie, F. Unique features of the basal cells of human prostate epithelium. Microsc. Res. Tech. 2000, 51, 436–446. [Google Scholar] [CrossRef]
- Lang, S.H.; Stark, M.; Collins, A.; Paul, A.B.; Stower, M.J.; Maitland, N.J. Experimental prostate epithelial morphogenesis in response to stroma and three-dimensional Matrigel culture. Cell Growth Differ. 2001, 12, 631–640. [Google Scholar] [PubMed]
- Heer, R.; Collins, A.T.; Robson, C.N.; Shenton, B.K.; Leung, H.Y. KGF suppresses alpha2beta1 integrin function and promotes differentiation of the transient amplifying population in human prostatic epithelium. J. Cell Sci. 2006, 119, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Litvinov, I.V.; Vander Griend, D.J.; Xu, Y.; Antony, L.; Dalrymple, S.L.; Isaacs, J.T. Low-calcium serum-free defined medium selects for growth of normal prostatic epithelial stem cells. Cancer Res. 2006, 66, 8598–8607. [Google Scholar] [CrossRef] [PubMed]
- Lamb, L.E.; Knudsen, B.S.; Miranti, C.K. E-cadherin-mediated survival of androgen-receptor-expressing secretory prostate epithelial cells derived from a stratified in vitro differentiation model. J. Cell Sci. 2010, 123, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthaus, W.R.; Iaquinta, P.J.; Drost, J.; Gracanin, A.; Van Boxtel, R.; Wongvipat, J.; Dowling, C.M.; Gao, D.; Begthel, H.; Sachs, N.; et al. Identification of multipotent luminal progenitor cells in human prostate organoid cultures. Cell 2014, 159, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Inokuchi, J.; Lau, A.; Tyson, D.R.; Ornstein, D.K. Loss of annexin A1 disrupts normal prostate glandular structure by inducing autocrine IL-6 signaling. Carcinogenesis 2009, 30, 1082–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandellini, P.; Profumo, V.; Casamichele, A.; Fenderico, N.; Borrelli, S.; Petrovich, G.; Santilli, G.; Callari, M.; Colecchia, M.; Pozzi, S.; et al. MiR-205 regulates basement membrane deposition in human prostate: Implications for cancer development. Cell Death Differ. 2012, 19, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.H. C-terminal tensin-like (CTEN): A promising biomarker and target for cancer. Int. J. Biochem. Cell Biol. 2014, 51, 150–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Mizokami, A.; Izumi, K.; Narimoto, K.; Shima, T.; Zhang, J.; Dai, J.; Keller, E.T.; Namiki, M. CTEN/tensin 4 expression induces sensitivity to paclitaxel in prostate cancer. Prostate 2010, 70, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.G.; Chang, S.L.; Erho, N.; Yu, M.; Lehrer, J.; Alshalalfa, M.; Speers, C.; Cooperberg, M.R.; Kim, W.; Ryan, C.J.; et al. Associations of luminal and basal subtyping of prostate cancer with prognosis and response to androgen deprivation therapy. JAMA Oncol. 2017, 3, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.A.; Mitrofanova, A.; Bergren, S.K.; Abate-Shen, C.; Cardiff, R.D.; Califano, A.; Shen, M.M. Lineage analysis of basal epithelial cells reveals their unexpected plasticity and supports a cell-of-origin model for prostate cancer heterogeneity. Nat. Cell Biol. 2013, 15, 274–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoyanova, T.; Cooper, A.R.; Drake, J.M.; Liu, X.; Armstrong, A.J.; Pienta, K.J.; Zhang, H.; Kohn, D.B.; Huang, J.; Witte, O.N.; et al. Prostate cancer originating in basal cells progresses to adenocarcinoma propagated by luminal-like cells. Proc. Natl. Acad. Sci. USA 2013, 110, 20111–20116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, D.A.; Zong, Y.; Memarzadeh, S.; Xin, L.; Huang, J.T.; Witte, O.N. Basal epithelial stem cells are efficient targets for prostate cancer initiation. Proc. Natl. Acad. Sci. USA 2010, 107, 2610–2615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.A.; Toivanen, R.; Bergren, S.K.; Chambon, P.; Shen, M.M. Luminal cells are favored as the cell of origin for prostate cancer. Cell Rep. 2014, 8, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.S.; Huang, J.T.; Guo, C.Y.; Garraway, I.P.; Witte, O.N. Identification of a cell of origin for human prostate cancer. Science 2010, 329, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Strand, D.W.; Goldstein, A.S. The many ways to make a luminal cell and a prostate cancer cell. Endocr. Relat. Cancer 2015, 22, T187–T197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, E.M.; Marchionni, L.; Huang, Z.; Simons, B.; Blackman, A.; Yu, W.; Parmigiani, G.; Berman, D.M. Androgen-induced programs for prostate epithelial growth and invasion arise in embryogenesis and are reactivated in cancer. Oncogene 2008, 27, 7180–7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, C.; Mecham, B.; Dumpit, R.; Coleman, I.; Bhattacharjee, M.; Chen, Q.; Sikes, R.A.; Nelson, P.S. Conserved Gene Expression programs integrate mammalian prostate development and tumorigenesis. Cancer Res. 2009, 69, 1739–1747. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Kitaoka, M.; Hamada, Y.; Walker, M.M.; Waxman, J.; Kypta, R.M. Regulation of prostate cell growth and morphogenesis by Dickkopf-3. Oncogene 2006, 25, 6528–6537. [Google Scholar] [CrossRef] [Green Version]
- Romero, D.; Kypta, R. Dickkopf-3 function in the prostate: Implications for epithelial homeostasis and tumor progression. BioArchitecture 2013, 3, 42–44. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Shih, Y.P.; Li, T.; Carraway, K.L.; Lo, S.H. CTEN prolongs signaling by EGFR through reducing its ligand-induced degradation. Cancer Res. 2013, 73, 5266–5276. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-C.; Si, L.; White, R.W.D.; Lo, S.H. The phosphotyrosine-independent interaction of DLC-1 and the SH2 domain of cten regulates focal adhesion localization and growth suppression activity of DLC-1. J. Cell Biol. 2007, 176, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Debnath, J.; Muthuswamy, S.K.; Brugge, J.S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods 2003, 30, 256–268. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.-M.; Liao, Y.-C. Downregulation of C-Terminal Tensin-Like Protein (CTEN) Suppresses Prostate Cell Proliferation and Contributes to Acinar Morphogenesis. Int. J. Mol. Sci. 2018, 19, 3190. https://doi.org/10.3390/ijms19103190
Wu W-M, Liao Y-C. Downregulation of C-Terminal Tensin-Like Protein (CTEN) Suppresses Prostate Cell Proliferation and Contributes to Acinar Morphogenesis. International Journal of Molecular Sciences. 2018; 19(10):3190. https://doi.org/10.3390/ijms19103190
Chicago/Turabian StyleWu, Wei-Ming, and Yi-Chun Liao. 2018. "Downregulation of C-Terminal Tensin-Like Protein (CTEN) Suppresses Prostate Cell Proliferation and Contributes to Acinar Morphogenesis" International Journal of Molecular Sciences 19, no. 10: 3190. https://doi.org/10.3390/ijms19103190