In Vitro Preservation of Transgenic Tomato (Solanum lycopersicum L.) Plants Overexpressing the Stress-Related SlAREB1 Transcription Factor

Abstract
:1. Introduction
2. Results
2.1. Overexpression of SlAREB1 in Tomato
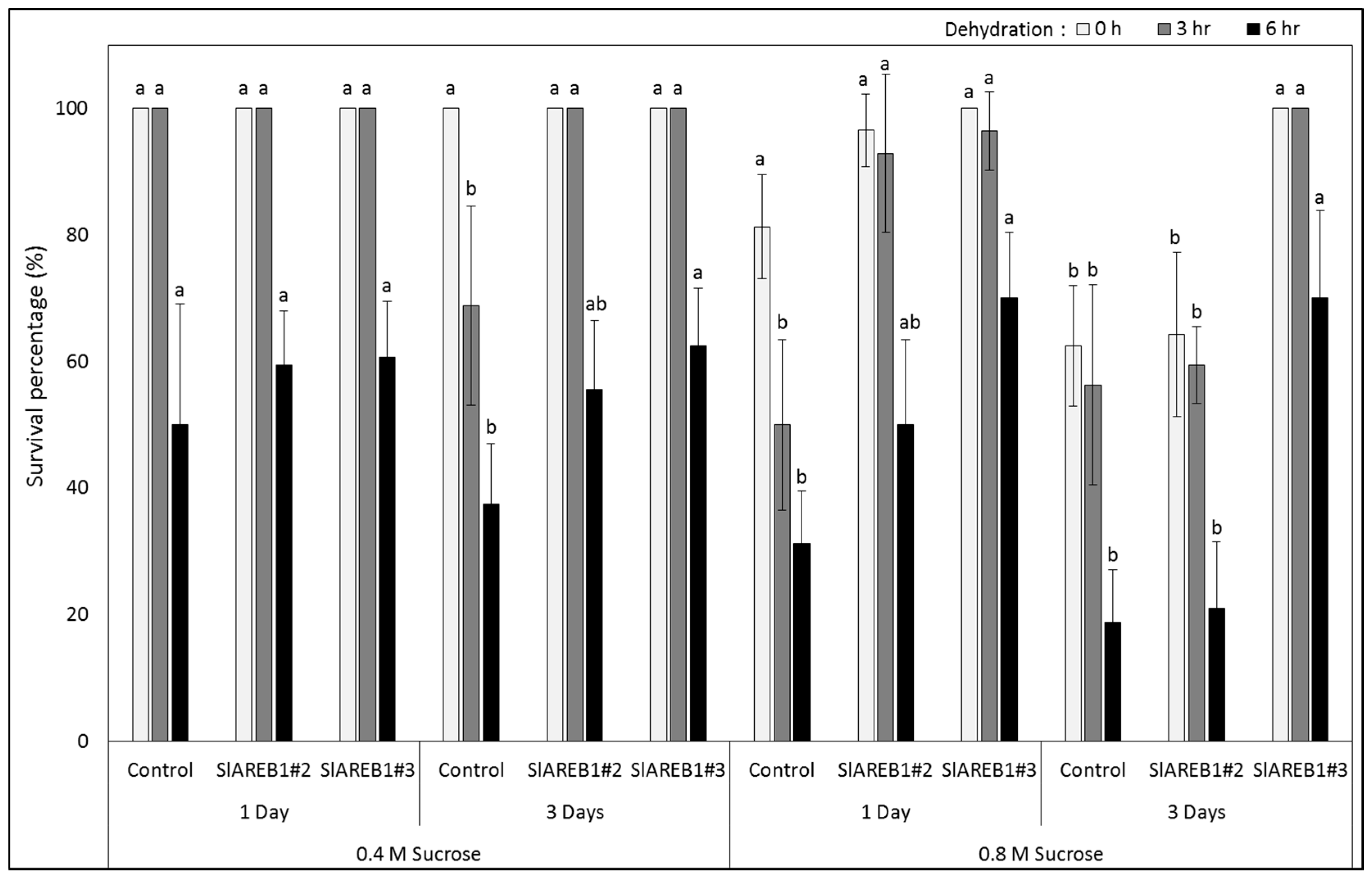
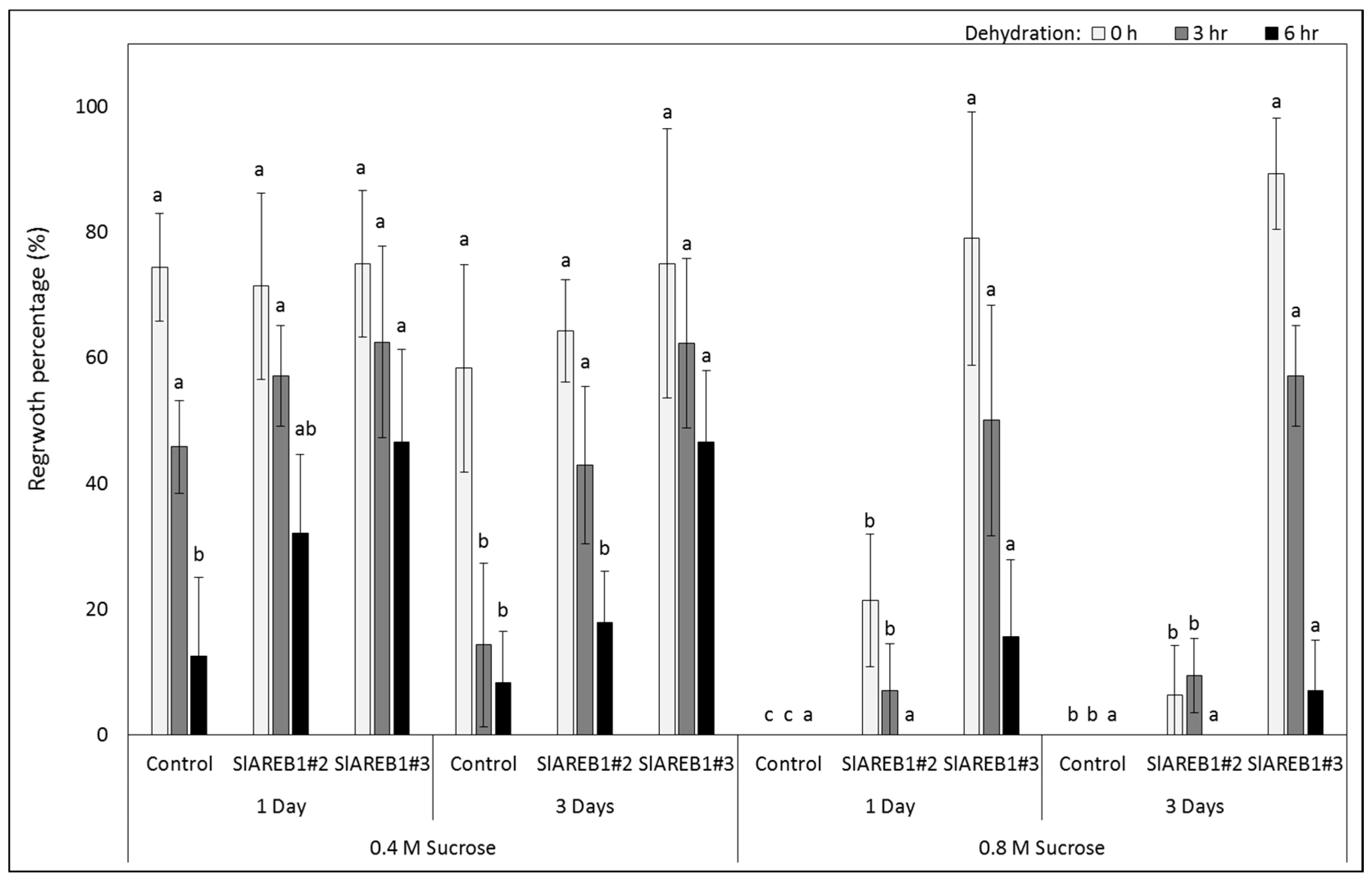
2.2. Slow Growth Preservation
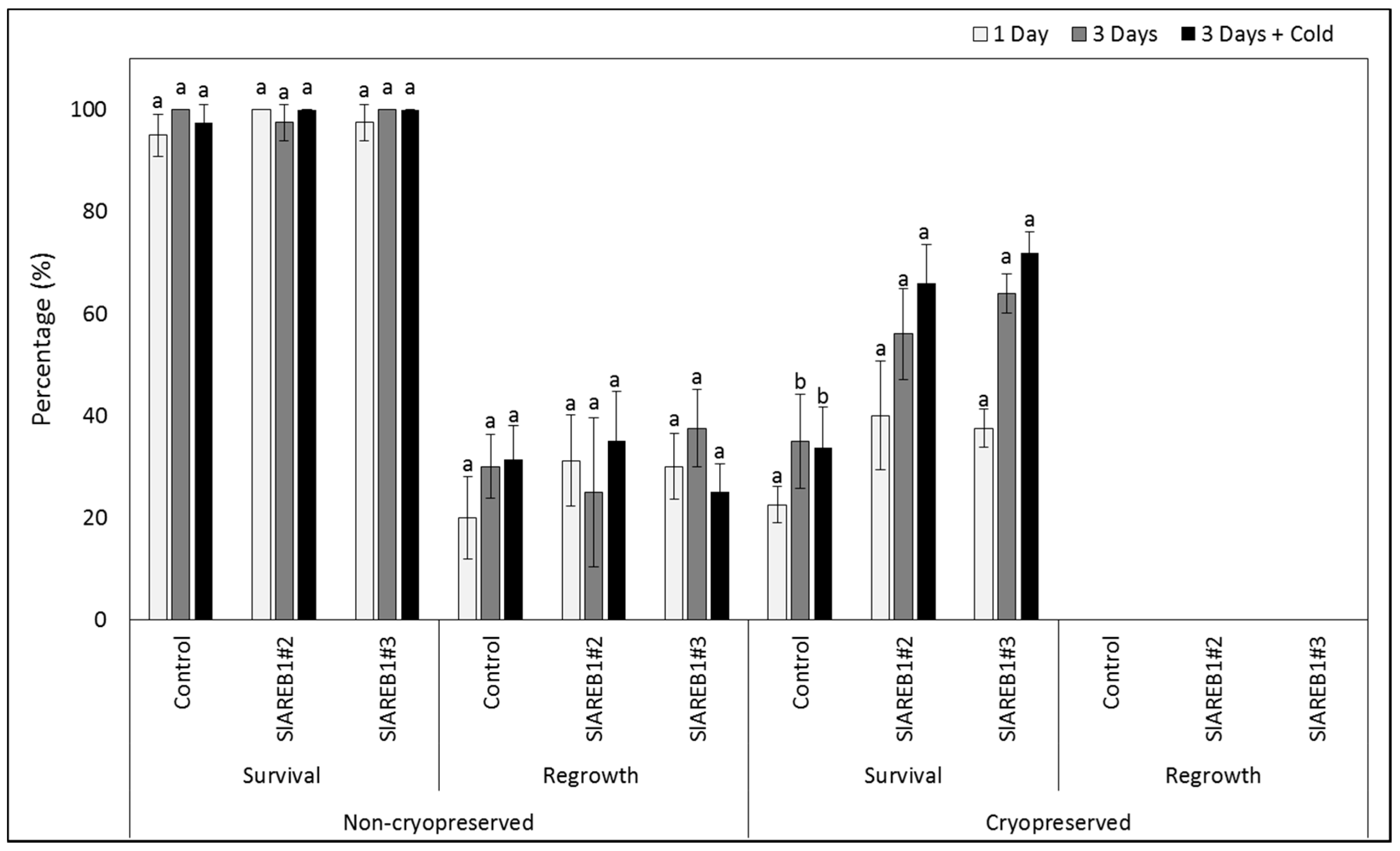
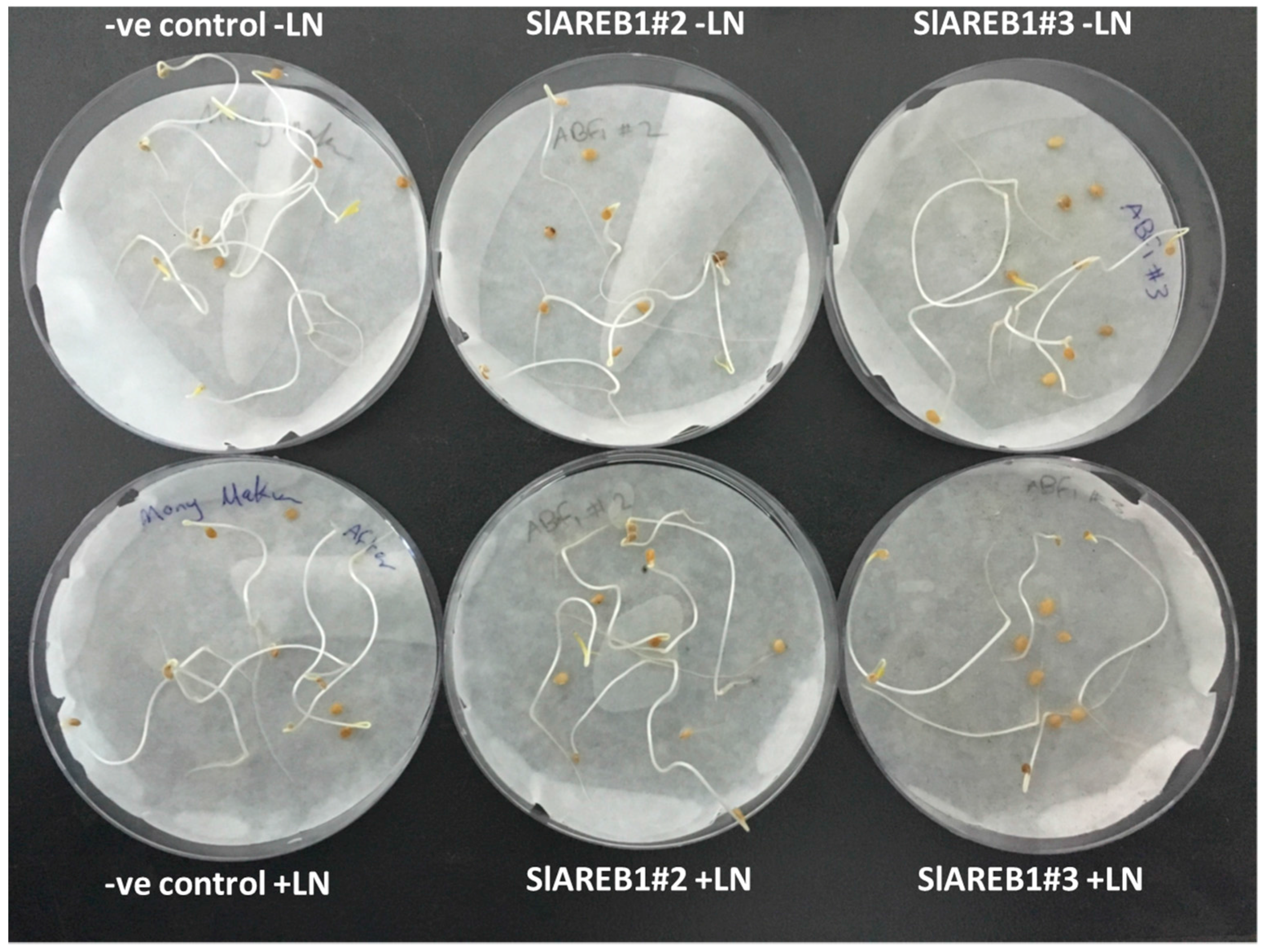
2.3. Cryopreservation
3. Discussion
4. Materials and Methods
4.1. Cloning of SlAREB1 Gene
4.2. Plant Material
4.3. In Vitro Establishment of Transgenic Lines
4.4. Slow Growth Preservation
4.5. Cryopreservation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| AREB | ABA responsive element binding |
| LN | Liquid nitrogen |
| MS media | Murashige and Skoog Medium |
| PVS2 | Plant Vitrification solution 2 |
| PCR | Polymerase chain reaction |
| RT-PCR | Real time-Polymerase chain reaction |
| SPSS | Statistical Package for Social Sciences |
| TTC | 2,3,5-Triphenyltetrazolium chloride salt solution |
| V-cryoplates | Vitrification-cryoplates |
References
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Nakashima, K.; Miyakawa, T.; Kuromori, T.; Tanokura, M.; Shinozaki, K. Molecular basis of the core regulatory network in ABA responses: Sensing, signalling and transport. Plant Cell Physiol. 2010, 51, 1821–1839. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Ann. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Fujita, M.; Fujita, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Engineering drought tolerance in plants: Discovering and tailoring genes to unlock the future. Curr. Opin. Biotechnol. 2006, 17, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y. The role ABF family bZIP class transcription factors in stress response. Physiol. Plant. 2006, 126, 519–527. [Google Scholar] [CrossRef]
- Orellana, S.; Yañez, M.; Espinoza, A.; Verdugo, I.; González, E.; Ruiz-Lara, S.; Casaretto, J.A. The transcription factor SlAREB1 confers drought, salt stress tolerance and regulates biotic and abiotic stress-related genes in tomato. Plant Cell Environ. 2010, 33, 2191–2208. [Google Scholar] [CrossRef] [PubMed]
- Bastias, A.; López-Climent, M.; Valcárcel, M.; Rosello, S.; Gómez-Cadenas, A.; Casaretto, J.A. Modulation of organic acids and sugar content in tomato fruits by an abscisic acid-regulated transcription factor. Physiol. Plant. 2011, 141, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.K. Plant genetic resources: Advancing conservation and use through biotechnology. Afr. J. Biotechnol. 2004, 3, 136–145. [Google Scholar] [CrossRef]
- Kulus, D.; Zalewska, M. Cryopreservation as a tool used in long-term storage of ornamental species: A review. Sci. Hort. 2014, 168, 88–107. [Google Scholar] [CrossRef]
- Engelmann, F. In vitro conservation methods. In Biotechnology and Plant Genetic Resources Conservation and Use; Ford-Lloyd, B.V., Newburry, J.H., Callow, J.A., Eds.; CABI: Wallingford, England, UK, 1997; pp. 119–161. ISBN 0851991424. [Google Scholar]
- Engelmann, F. Plant cryopreservation: Progress and prospects. In Vitro Cell Dev. Biol. Plant 2004, 40, 427–433. [Google Scholar] [CrossRef]
- Pence, V.C. Tissue cryopreservation for plant conservation: Potential and challenges. Int. J. Plant. Sci. 2014, 175, 40–45. [Google Scholar] [CrossRef]
- Kaviani, B. Conservation of plant genetic resources by cryopreservation. Aust. J. Crop. Sci. 2011, 5, 778–800. [Google Scholar]
- Engelmann, F. Importance of cryopreservation for the conservation of plant genetic resources. In Cryopreservation of Tropical Plant Germplasm: Current Research Progress and Application; Engelmann, F., Takagi, H., Eds.; JIRCAS: Tsukuba, Japan; IPGRI: Rome, Italy, 2000; pp. 8–20. [Google Scholar]
- Kami, D. Cryopreservation of Plant Genetic Resources, Current Frontiers in Cryobiology; Katkov, I., Ed.; In Tech: Munich, Germany, 2012. [Google Scholar]
- Fabre, J.; Dereuddre, J. Encapsulation-dehydration: A new approach to cryopreservation of Solanum shoot tips. Cryo Lett. 1990, 11, 413–426. [Google Scholar]
- Coste, A.; Deliu, C.; Vălimăreanu, S.; Halmagyi, A. Cryopreservation and acclimatization of Lycopersiconesculentum (Mill.) genotypes. Not. Bot. Horti. Agrobo. 2014, 42, 466–471. [Google Scholar] [CrossRef]
- Coste, A.; Şuteu, D.; Bacila, L.; Deliu, C.; Vălimăreanu, S.; Halmagyi, A. Genetic integrity assessment of cryopreserved tomato (Lycopersicon esculentum Mill.) genotypes. Turk. J. Biol. 2015, 39, 638–648. [Google Scholar] [CrossRef]
- Niino, T.; Yamamoto, S.; Fukui, K.; Castillo Martínez, C.R.; Valle Arizaga, M.; Matsumoto, T.; Engelmann, F. Dehydration improves cryopreservation of mat rush (Juncus decipiens Nakai) basal stem buds on cryo-plates. Cryo Lett. 2013, 34, 549–560. [Google Scholar]
- Engelmann-Sylvestre, I.; Engelmann, F. Cryopreservation of in vitro-grown shoot tips of Clinopodium odorum using aluminium cryo-plates. In Vitro Cell Plant 2015, 51, 185–191. [Google Scholar] [CrossRef]
- Salma, M.; Fkic, L.; Engelmann-Sylvestrea, I.; Niino, T.; Engelmann, F. Comparison of droplet-vitrification and d-cryoplate for cryopreservation of date palm (Phoenix dactylifera L.) polyembryonic masses. Sci. Hort. 2014, 179, 91–97. [Google Scholar] [CrossRef]
- Stanwood, P.C. Cryopreservation 0/Plant Cells and Organs. In Cryopreservation of Seed Germplasm for Genetic Conservation; Kartha, K.K., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 199–225. [Google Scholar]
- Zevallos, B.; Cejas, I.; Engelmann, F.; Carputo, D.; Aversano, R.; Scarano, M.T.; Yanes, E.; Martínez-Montero, M.; Lorenzo, J.C. Phenotypic and molecular characterization of plants regenerated from non-cryopreserved and cryopreserved wild Solanum lycopersicum Mill. seeds. Cryo Lett. 2014, 35, 216–225. [Google Scholar]
- Corredoira, E.; San-José, M.C.; Vieitez, A.M.; Ballester, A. Improving genetic transformation of European chestnut and cryopreservation of transgenic lines. Plant Cell Tissue Organ Cult. 2007, 91, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Li, J.W.; Li, H.H.; Wang, R.R.; Gao, X.; Wang, Q. Cryopreservation for retaining morphology, genetic integrity, and foreign genes in transgenic plants of Torenia fournier. Acta Physiol. Plant 2016, 38, 8. [Google Scholar] [CrossRef]
- Osorio-Saenz, A.; Oscar Mascorro-Gallardo, J.; del Rocío Valle-Sandoval, M.; Teresa González-Arnao, M.; Engelmann, F. Genetically engineered trehalose accumulation improves cryopreservation tolerance of chrysanthemum (Dendranthema grandiflorum Kitam.) shoot-tips. Cryo Lett. 2011, 32, 477–486. [Google Scholar]
- Yáñez, M.; Cáceres, S.; Orellana, S.; Bastías, A.; Verdugo, I.; Ruiz-Lara, S.; Casaretto, J.A. An abiotic stress-responsive bZIP transcription factor from wild and cultivated tomatoes regulates stress-related genes. Plant Cell Rep. 2009, 28, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Pereira, J. In vitro conservation of Piper aduncum and Piper hispidinervum under slow-growth conditions. Pesq. Agropec. Bras. 2011, 46, 384–389. [Google Scholar] [CrossRef]
- Jarret, R.; Gawel, N. Abscisic acid-induced growth inhibition of sweet potato (Ipomoea batatas L.) in vitro. Plant Cell Tissue Organ Cult. 1991, 24, 13–18. [Google Scholar] [CrossRef]
- Capuana, M.; Di Lonardo, S. In vitro conservation of chestnut (Castanea sativa) by slow growth. In Vitro Cell. Dev. Biol. Plant 2013, 49, 605–610. [Google Scholar] [CrossRef]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [PubMed]
- Karipidis, C.H.; Douma, D. Tomato pollen storage at freeze and cryogenic temperature effects on viability and fecundity. Acta Hort. 2011, 908, 257–263. [Google Scholar] [CrossRef]
- Grout, B.W.; Westcott, R.J.; Henshaw, G.G. Survival of shoot meristems of tomato seedlings frozen in liquid nitrogen. Cryobiology 1978, 15, 478–483. [Google Scholar] [CrossRef]
- Yamamoto, S.; Rafique, T.; Arizaga, M.; Fukui, K.; Gutierrez, E.; Martinez, C.; Watanabe, K. The Aluminium cryo-plate increases efficiency of cryopreservation protocols for potato shoot tips. Am. J. Potato Res. 2015, 92, 250–257. [Google Scholar] [CrossRef]
- Rafique, T.; Yamamoto, S.; Fukui, K.; Mahmood, Z.; Niino, T. Cryopreservation of sugarcane using the V cryo-plate technique. Cryo Lett. 2015, 36, 51–59. [Google Scholar]
- Yamamoto, S.; Rafique, T.; Fukui, K.; Sekizawa, K.; Niino, T. Development of a cryopreservation procedure using aluminium cryo-plates. Cryo Lett. 2011, 32, 256–265. [Google Scholar]
- Marković, Z.; Chatelet, P.; Sylvestre, I.; Kontić, J.; Engelmann, F. Cryopreservation of grapevine (Vitis vinifera L.) in vitro shoot tips Cent. Eur. J. Biol. 2013, 8, 993–1000. [Google Scholar] [CrossRef]
- Moges, A.D.; Karam, N.S.; Shibli, R.A. Cryopreservation of African violet (Saintpaulia ionantha Wendl.) shoots tips. In Vitro Cell Dev. Biol. Plant 2004, 40, 389–395. [Google Scholar] [CrossRef]
- Sharaf, S.A.; Shibli, R.A.; Kasrawi, M.A.; Baghdadi, S.H. Cryopreservation of wild shih (Artemisia herba-alba Asso.) shoot-tips by encapsulation-dehydration and encapsulation-vitrification. Plant Cell Tissue Organ Cult. 2012, 108, 437–444. [Google Scholar] [CrossRef]
- Al-Ababneh, S.; Shibli, R.A.; Karam, N.S.; Shatnawi, M.A. Cryopreservation of bitter almond (Amygdalus communis L.) shoot tips by encapsulation-dehydration and vitrification. Adv. Hort. Sci. 2003, 17, 15–20. [Google Scholar]
- Tarré, E.; Pires, B.B.M.; Guimarães, A.P.M.; Carneiro, L.A.; Forzza, R.C.; Mansur, E. Germinability after desiccation, storage and cryopreservation of seeds from endemic Encholirium Mart. ex Schult. & Schult. f. and Dyckia Schult. & Schult. f. species (Bromeliaceae). Acta Bot. Bras. 2007, 21, 777–783. [Google Scholar] [CrossRef]
- Finkelstein, R.; Gampala, S.L.; Lynch, T.J.; Thomas, T.L.; Rock, C.D. Redundant and distinct functions of the ABA Response loci ABA-INSENSITIVE(ABI)5 and ABRE-BINDING FACTOR (ABF)3. Plant Mol. Biol. 2005, 59, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–479. [Google Scholar] [CrossRef]
- Al-Abdallat, A.M.; Al-Debei, H.S.; Ayad, J.Y.; Hasan, S. Over-expression of SlSHN1 gene improves drought tolerance by increasing cuticular wax accumulation in tomato. Int. J. Mol. Sci. 2014, 15, 19499–19515. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, C.T.; Moran Lauter, A.N.; Scott, M.P. Determination of transgene copy number by real-time quantitative PCR. Methods Mol. Biol. 2009, 526, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Rabaa, M.; Shibli, R.A.; Shatnawi, M.A. Cryopreservation of Teucrium polium L. shoot-tips by vitrification and encapsulation-dehydration. Plant Cell Tissue Organ Cult. 2012, 110, 371–382. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sucrose Concentration (mm) | Plant Height (cm) | Roots Number | Leaves Number | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | SlAREB1#2 | SlAREB1#3 | Control | SlAREB1#2 | SlAREB1#3 | Control | SlAREB1#2 | SlAREB1#3 | |
| 6 weeks | |||||||||
| 0 | 1.00 a * ± 0.00 ** | 1.00 a ± 0.00 | 1.21 a ± 0.10 | 0.00 a ± 0.00 | 0.00 a ± 0.00 | 0.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.14 a ± 0.67 |
| 100 | 5.32 a ± 0.59 | 5.85 a ± 0.42 | 5.99 a ± 0.66 | 7.14 a ± 1.12 | 7.66 a ± 1.55 | 7.89 a ± 2.49 | 7.86 a ± 1.28 | 7.29 a ± 1.67 | 6.43 a ± 1.37 |
| 200 | 1.71 b ± 0.24 | 6.13 a ± 1.18 | 6.47 a ± 0.50 | 0.00 b ± 0.00 | 10.71 a ± 2.31 | 9.57 a ± 1.86 | 3.57 b ± 1.03 | 6.29 a ± 1.23 | 5.71 a ± 1.23 |
| 300 | 1.14 b ± 0.13 | 1.97 b ± 0.36 | 3.84 a ± 0.92 | 0.00 b ± 0.00 | 0.57 b ± 0.56 | 3.43 a ± 0.83 | 2.00 b ± 0.61 | 1.57 b ± 0.83 | 6.00 a ± 1.22 |
| 12 weeks | |||||||||
| 0 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.34 a ± 0.11 | 0.00 a ± 0.00 | 0.00 a ± 0.00 | 0.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.43 a ± 0.77 |
| 100 | 8.47 a ± 1.10 | 8.55 a ± 0.68 | 9.10 a ± 1.10 | 10.14 a ± 2.39 | 9.23 a ± 1.29 | 10.86 a ± 1.32 | 12.29 a ± 2.71 | 11.46 a ± 2.77 | 10.43 a ± 1.58 |
| 200 | 2.54 b ± 0.21 | 7.27 a ± 1.29 | 8.77 a ± 0.72 | 0.14 b ± 0.04 | 13.57 a ± 3.33 | 13.43 a ± 3.03 | 5.57 b ± 1.59 | 10.86 a ± 2.79 | 9.43 a ± 1.24 |
| 300 | 1.24 b ± 0.12 | 2.22 b ± 0.44 | 5.27 a ± 0.76 | 0.00 b ± 0.00 | 0.86 b ± 0.73 | 7.43 a ± 2.10 | 2.57 b ± 0.56 | 2.43 b ± 0.56 | 8.57 a ± 2.19 |
| Transgenic Lines | Sucrose Concentration (mm) | ABA Concentration (µm) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0.00 | 100 | 200 | 300 | 0 | 4 | 8 | 12 | |
| Control | 0.00 a * ± 0.00 ** | 78.6 a ± 10.97 | 38.5 b ± 13.00 | 21.4 b ± 10.97 | 71.4 a ± 12.07 | 71.4 a ± 12.07 | 81.8 a ± 10.31 | 83.0 a ± 9.96 |
| SlAREB1#2 | 0.00 a ± 0.00 | 81.8 a ± 10.31 | 71.4 a ± 12.07 | 38.5 b ± 13.00 | 76.9 a ± 11.26 | 78.6 a ± 10.97 | 84.6 a ± 9.64 | 83.0 a ± 9.96 |
| SlAREB1#3 | 0.00 a ± 0.00 | 85.7 a ± 9.35 | 78.6 a ± 10.97 | 92.3 a ± 7.12 | 85.7 a ± 9.35 | 69.2 a ± 12.97 | 78.6 a ± 10.97 | 78.56 a ± 10.97 |
| ABA Concentration (µm) | Plant Height (cm) | Roots Number | Leaves Number | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | SlAREB1#2 | SlAREB1#3 | Control | SlAREB1#2 | SlAREB1#3 | Control | SlAREB1#2 | SlAREB1#3 | |
| 6 weeks | |||||||||
| 0 | 5.04 a * ± 0.55 ** | 5.14 a ± 1.31 | 5.01 a ± 0.70 | 5.86 a ± 1.42 | 6.57 a ± 2.01 | 5.57 a ± 1.03 | 6.14 a ± 1.42 | 5.71 a ± 1.58 | 5.29 a ± 1.00 |
| 4 | 2.91 a ± 0.66 | 2.46 a ± 0.37 | 2.11 a ± 0.31 | 5.43 a ± 1.81 | 4.57 a ± 1.20 | 1.86 b ± 1.55 | 5.14 a ± 1.42 | 5.29 a ± 1.80 | 2.86 b ± 1.13 |
| 8 | 1.41 a ± 0.11 | 1.43 a ± 0.15 | 1.53 a ± 0.25 | 0.00 a ± 0.00 | 0.14 a ± 0.40 | 0.00 a ± 0.00 | 2.71 a ± 0.52 | 3.14 a ± 0.95 | 3.00 a ± 0.86 |
| 12 | 1.46 a ± 0.20 | 1.51 a ± 0.13 | 1.43 a ± 0.18 | 0.00 a ± 0.00 | 0.29 a ± 0.80 | 0.00 a ± 0.00 | 2.57 a ± 0.83 | 3.29 a ± 0.80 | 3.14 a ± 0.40 |
| 12 weeks | |||||||||
| 0 | 7.19 a ± 1.00 | 7.33 a ± 1.75 | 6.93 a ± 0.56 | 9.86 a ± 2.69 | 8.71 a,b ± 1.18 | 7.57 b ± 2.10 | 13.00 a ± 3.56 | 11.71 a ± 2.91 | 11.00 a ± 1.83 |
| 4 | 6.90 a ± 1.19 | 7.14 a ± 1.61 | 2.26 b ± 0.32 | 8.43 a ± 2.79 | 6.86 a ± 0.95 | 1.86 b ± 1.55 | 9.71 a ± 2.42 | 8.29 a ± 1.58 | 3.86 b ± 1.42 |
| 8 | 1.53 a ± 0.15 | 1.94 a ± 0.29 | 1.60 a ± 0.15 | 0.14 a ± 0.40 | 0.57 a ± 0.83 | 0.14 a ± 0.40 | 5.43 a ± 0.83 | 5.14 a ± 0.95 | 4.43 a ± 0.83 |
| 12 | 1.73 a ± 0.27 | 1.73 a ± 0.26 | 1.61 a ± 0.20 | 0.00 a ± 0.00 | 0.29 a ± 0.52 | 0.00 a ± 0.00 | 5.29 a ± 0.80 | 4.14 a ± 0.73 | 3.71 a ± 1.00 |
| Transgenic Lines | Non-Cryopreserved | Cryopreserved | ||||||
|---|---|---|---|---|---|---|---|---|
| Number of Days | Number of Days | |||||||
| 5 | 7 | 10 | 14 | 5 | 7 | 10 | 14 | |
| Control | 60 a * ± 7.07 ** | 80 a ± 7.07 | 84 a ± 5.47 | 100 a ± 0.00 | 60 a ± 7.07 | 78 a ± 7.07 | 86 a ± 5.47 | 100 a ± 0.00 |
| SlAREB1#2 | 50 a ± 12.24 | 72 a ± 4.47 | 78 a ± 5.47 | 94 a ± 5.47 | 60 a ± 12.24 | 72 a ± 4.47 | 76 a ± 5.47 | 96 a ± 8.49 |
| SlAREB1#3 | 25 b ± 7.07 | 44 b ± 5.47 | 50 b ± 10.00 | 54 b ± 8.49 | 30 b ± 7.07 | 42 b ± 5.47 | 50 b ± 10.00 | 74 b ± 5.47 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Abdallat, A.M.; Shibli, R.A.; Akash, M.W.; Rabbaa, M.; Al-Qudah, T. In Vitro Preservation of Transgenic Tomato (Solanum lycopersicum L.) Plants Overexpressing the Stress-Related SlAREB1 Transcription Factor. Int. J. Mol. Sci. 2017, 18, 1477. https://doi.org/10.3390/ijms18071477
Al-Abdallat AM, Shibli RA, Akash MW, Rabbaa M, Al-Qudah T. In Vitro Preservation of Transgenic Tomato (Solanum lycopersicum L.) Plants Overexpressing the Stress-Related SlAREB1 Transcription Factor. International Journal of Molecular Sciences. 2017; 18(7):1477. https://doi.org/10.3390/ijms18071477
Chicago/Turabian StyleAl-Abdallat, Ayed M., Rida A. Shibli, Muhanad W. Akash, Manar Rabbaa, and Tamara Al-Qudah. 2017. "In Vitro Preservation of Transgenic Tomato (Solanum lycopersicum L.) Plants Overexpressing the Stress-Related SlAREB1 Transcription Factor" International Journal of Molecular Sciences 18, no. 7: 1477. https://doi.org/10.3390/ijms18071477